
Культура столонов и регуляция роста растений и клубнеобразования у картофеля in vitro (03. 00. 12 физиология и биохимия растений)
| Вид материала | Автореферат |
СодержаниеЭкспериментальная часть Результаты исследований Регуляция клубнеобразования у различных Динамика накопления биомассы органов картофеля у Список работ, опубликованных по теме диссертации |
- Регенерационная активность разных генотипов пшеницы и эгилопса в культуре in vitro, 499.46kb.
- Эпигенетическая регуляция фотосинтетической ассимиляции со 2 у хлопчатника (03. 00., 544.35kb.
- Рабочая программа дисциплины «физиология» (физиология растений) Код дисциплины по учебному, 269.31kb.
- «Физиология растений», 308.24kb.
- Регуляция активности свободных мультиферментных комплексов цикла кальвина у высших, 632.07kb.
- «Физиология растений», 24.13kb.
- Учебно-методический комплекс дисциплины "Физиология растений" вузовского компонента, 1046.86kb.
- Научно-методической комиссии по технологии возделывания, защите растений, хранению, 1342.92kb.
- Лекция 11 Тема: Высшие растения. Происхождение высших наземных растений. Отдел Моховидные, 276.22kb.
- Программа-минимум кандидатского экзамена по специальности 06. 01. 09 «Растениеводство», 94.9kb.
На правах рукописи
Назарова Нигора Нурахмадовна
Культура столонов и регуляция роста растений и клубнеобразования у картофеля in vitro
(03.00.12 – физиология и биохимия растений)
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Душанбе-2006
Работа выполнена в Институте физиологии растений и генетики АН Республики Таджикистан
Научный руководитель: член-корреспондент
АН Республики Таджикистан,
доктор биологических наук, профессор К.А. Алиев
Официальные оппоненты: доктор биологических наук, профессор Бободжанов В.А.,
член-корр. АН Республики Таджикистан, доктор сельскохозяйственных наук, Фелалиев А.С.
Ведущая организация: Таджикский аграрный университет
Защита состоится «__»________2006г. в___часов на заседании диссертационного совета Д047.001.01 при Институте физиологии растений и генетики АН Республики Таджикистан (734063, г. Душанбе, ул. Айни, 299/2).
lab.gen@mail.ru
С диссертацией можно ознакомиться в Центральной научной библиотеке им. Индиры Ганди АН Республики Таджикистан.
Автореферат разослан «__»__________2006 г.
Ученый секретарь
диссертационного совета
доктор биологических наук Джумаев Б.Б.
ВВЕДЕНИЕ
Актуальность работы. Прогресс в области современной биотехнологии растений непосредственно связан с разработкой методических приемов культивирования клеток in vitro. Этим методом получен ряд сомаклональных линий, форм и сортов растений с ценными хозяйственными признаками, которые используются во многих странах мира.
С другой стороны, человеческое сообщество остро нуждается в новых знаниях и технологиях, с тем, чтобы увеличить производство продовольствия, улучшить состояние здравохранения и охрану окружающей среды. В решение этих проблем биотехнология вносит большой вклад. Поэтому применение методов биотехнологии в Республике Таджикистан является одним из важнейших факторов повышения урожайности сельскохозяйственных культур и, следовательно, экономического потенциала.
С использованием методов генной и клеточной инженерии получен ряд линий, форм культурных растений (картофель, соя, кукуруза и др.), обладающих признаками устойчивости к вирусам, патогенам и пластичности к стрессовым экологическим факторам. Благодаря этому сокращаются сроки получения новых сортов с хозяйственно полезными признаками и получения свободных от вирусов и патогенов семенного материала, что имеет большую перспективу для Республики Таджикистан.
В теоретическом плане представляет интерес изучение степени сортовой вариабельности ДНК, выявленной методом полимеразной цепной реакции (PCR), и физиологических процессов в культуре in vitro с целью разработки методов микроклонального размножения и микроклубнеобразования картофеля.
Зависимость клубнеобразования от действия таких факторов, как световой режим (фитохром), гормональный статус,метаболизм углеводов и т.д. показана в ряде работ (Чайлахян, 1984; 1988; Аксенова и др., 2002; Кузнецов, Кулаева, 2004).
Вместе с тем, вопрос о конкретных путях действия фитогормонов и углеводов и их взаимодействии в процессе инициации клубнеобразования остается открытым, т. к. имеющиеся результаты не всегда можно итерпретировать однозначно (Hussey, Stacey, 1995; Davies,1984).
Так, для перехода регенерантов к клубнеобразованию in vitro необходимо наличие высокой концентрации сахарозы (5-10%), но есть сведения о том, что сахароза мало влияет на инициацию клубнеобразования. Также много неясного относительно роли фитогормонов в инициации клубней. Так показано, что гиббереллины способствуют росту столонов, но ингибируют формирование клубней (Simko, 1994; Eving, 1995; Xin Xu et. al., 1998).
Для активизации процесса клубнеобразования использовались различные сочетания гормонов и углеводов (Palmer, Smith, 1987; Бутенко,1990). Но эффективного образования микроклубней не было достигнуто, так как повышенное содержание сахарозы или кинетина не индуцировали дополнительного образования столонов с последующим образованием клубней. Не было получено положительных результатов и при культивировании микрочеренков картофеля в темноте по методу G. Hussey и N.I. Stacey (1984), а также при изменении температурного режима (+12… +160С). Поэтому исследование условий, стимулирующих столоно- и клубнеобразование у картофеля in vitro, остаётся актуальной задачей. Нам известны всего лишь две публикации (Mingo-Castel et.al., 1976; Sassey, Stacey, 1984), в которых сообщается о попытке культивировать столоны картофеля in vitro.
В последние годы многие используют новый подход для регуляции клубнеобразования картофеля in vitro. Так, в частности используются генетически трансформированные регенеранты картофеля с изменённым гормональным статусом, такие, как трансформанты, несущие гены roLB и roLC Agrobacterium Rhizogenes (Аксёнова и др., 2002).
Было бы важно изучить возможность использования столонов картофеля в гормональной и углеводной регуляции экспрессии генов, инициации и роста клубней в системе in vitro. Такое изучение до сих пор не проводилось при создании системы свободных от вирусов и патогенов клубней картофеля с использованием методов биотехнологии.
Считаем, что изучение процессов инициации и роста клубней in vitro с использованием культуры столонов позволит выяснить роль углеводов и гормонов в инициации роста клубней и применить возможности методов биотехнологии для клонального размножения картофеля.
Цель и задачи работы. Целью настоящей работы является изучение особенностей культивирования изолированных столонов, возможности их использования в микроклональном размножении и регуляции клубнеобразования у генотипов картофеля. В соответствии с этим поставлены следующие задачи:
- разработка условий культивирования изолированных столонов картофеля;
- изучение ростовых процессов у регенерантов, полученных из изолированных столонов;
- изучение процессов микроклубнеобразования в зависимости от условий фотопериода;
- сравнительный анализ этапов клубнеобразования у картофеля в зависимости от воздействия углеводов в культуре in vitro.
Научная новизна работы.
- При изучении роста и размножения регенерантов картофеля in vitro обнаружена сезонность их морфогенной потенции, проявляющася в процессе микроклубнеобразования у пробирочных растений, а также при образовании столонов.
- Обнаружены некоторые особенности индукции роста изолированных столонов при изменении условий их культивирования in vitro. Полученные из столонов регенеранты обладают более высокой способностью к клубнеобразованию по сравнению с исходными меристемными растениями.
- Раработана эффективная методика культивирования столонов in vitro.
- Обнаружена сезонная зависимость каллусообразования у пробирочных растений, индуцируемых из столонов.
- Установленно, что процесс клубнеобразования зависит от содержания углеводов и является фенотипической реакцией генотипов картофеля. При сравнении клубнеобразования различных генотипов картофеля (исходного сорта Жуковский-ранний и его температуроустойчивой линии-регененранта) показана неоднозначная реакция по ряду характерных параметров роста, развития и индекса урожая на уровне пробирочных растений. Наиболее существенным отличием является усиление иннициации и роста клубней у клеточно-модифицированных растений под влиянием высокой концентрации сахарозы в культуральной среде по сравнению с исходным сортом Жуковский-ранний.
- Обнаружено, что процесс иннициации и темпа роста микроклубней в зависимости от концентрации сахарозы однонаправленны, но не одинаковы, так как масса сформировавшихся клубней в заключительной фазе их роста во многом определяется генотипом и, следовательно, требуют дифференцированных условий инициации и роста клубней in vitro.
- Установлена зависимость действия углеводов и гормонов в процессе дифференцировки столоновых клеток картофеля в культуре тканей in vitro, вызывающая активацию морфогенетической потенции растений.
Практическая значимость. Показана возможность использовния культуры столонов in vitro в практической биотехнологии картофеля. Регенеранты, полученные из культуры столонов, обладают значительно более высокими темпами микроклубнеобразования и свободны от вирусов и патогенов, что позволяет наладить новую технологию микроклонального размножения с целью их ускоренного внедрения в семеноводство картофеля.
Апробация работы: Материалы диссертации доложены/представлены на следующих конференциях, симпозиумах, совещаниях:
- Актуальные проблемы и перспективы развития физиологии растений, Душанбе, 2004;
- Transgenic plants and biosafety , Moscow, 2004;
- Экономика и наука Горно-Бадахшанской автономной области: прошлое, настоящее, будущее, Хорог, 2005.
- Использование оздоровленного материала в семеноводстве картофеля, Муминобад, 2005.
- Конференции молодых ученых Таджикского аграрного университета, Душанбе, 2003-2005 гг.
- Физиологические и молекулярно-генетические аспекты сохранения биоразнообразия, Вологда, Россия, 2005
- International Congress BioVision Alexandria, 2006
Структура и объем работы: Работа состоит из введения, обзора литературы, экспериментальной части, включающей описание материалов и методов, результатов исследований и их обсуждение, выводов, списка литературы. Работа изложена на 117 стр., содержит 26 рисунков, 9 таблиц. Количество цитированных источников 125.
Публикации. По материалам диссертации опубликовано 9 работ.
Экспериментальная часть
Объект исследований. В качестве объекта исследований служили сорт картофеля (Solanum tuberosum L.) Жуковский ранний и температуроустойчивая линия (ТУ-регенеранты), полученая от сорта Жуковский ранний (Давлятназарова и др., 2003). В Республике Таджикистан сорт Жуковский ранний зарекомендовал себя как перспективный, высокоурожайный и находится в процессе районирования. Эти растения переведены на культурную среду с использованием культуры апикальных меристем по рекомендации Института картофельного хозяйства РАСХН (2000) и по О.С. Мелик-Саркисову и др. (1988).
Столонообразование in vitro. В опытах использовали пробирочные растения картофеля сорта Жуковский ранний. Для культивирования растений in vitro использовали питательную среду Мурасиге и Скуга (МС), содержащую 0,6 % сахарозы и 1; 1,5; 2,5 мг/мл кинетина в зависимости от задач эксперимента.
Растения выращивали в световой комнате при освещении 6 тыс. люкс, при 16 –часовом фотопериоде и температуре +200С. Для индукции столонов использовали следующий способ: пробирочные растения в течение 12 ч поддерживали в световом режиме, затем помещали в темноту при температуре +22оС в течение 20-25 дней. В течение всего периода образования столонов питательную среду в пробирках не меняли. Измерение высоты растений и длины столонов отмечали каждые 10-15 дней.
Культивирование столонов in vitro. В работе использовали меристемные растения картофеля, культивируемые in vitro, сортов Жуковский ранний и Пикассо (среднепоздний). Культивирование изолированных столонов проводили на среде Мурасиге Скуга (МС) с добавлением 3-8 % сахарозы, 0,1-1,0 мг/л кинетина, 0,1-0,5 мг/л α-нафтилуксусной кислоты (НУК) в темноте в течение 8-30 дней, с переводом на свет при +220С. Растения выращивали при 16-часовом фотопериоде и температуре +22…+230С. В течение всего периода культивирования столонов питательную среду не меняли.
Для опытов по столоно- и клубнеобразованию использовали столоны размером 5-6 мм. С целью предотвращения истощения питательной среды использовали пробирки больших размеров (32X270 мм). После образования регенерантов из сегментов столонов растения черенковали обычным методом. Измерение высоты растений, длины столонов, инициацию клубней отмечали каждые 30 дней.
Условия образования микроклубней картофеля. В работе использовали регенеранты картофеля, полученные на основе сорта Жуковский ранний, несущие постоянно экспрессирующие гены устойчивости к высокой температуре (ТУ-регенеранты), и контрольные регенеранты сорта Жуковский ранний. Эти растения размножали клонированием in vitro на среде МС, содержащей 0,7% агара и витамины. Растения выращивали при 16-часовом освещении люминесцентными лампами дневного света.
В опытах использовали черенки с одним листочком, их высаживали на среду МС с 6% агаром и витаминами, содержащую от 2 до 9% сахарозы (в зависимости от варианта), с добавкой или без 0,5-1,5 мг/л кинетина или нафтилуксусной кислоты (НУК). Растения культивировали в течение 70 дней при 16-часовом освещении или в темноте. Через каждые 5 дней регистрировали число образовавшихся клубней, а в конце опыта – сырую массу всех органов растений-регенерантов: корней, побегов с листьями и клубней. Содержание вирусов в регенерантах определяли методом иммуноферментного анализа (ИФА). Необходимые реактивы для анализа вирусов получали из Института картофельного хозяйства РАСХН (Коренево, Россия). Опыты повторяли 2-3 раза. Анализировали по 25 растений регенерантов. На графиках и в таблицах приведены усредненные арифметические значения всех опытов.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Образование столонов картофеля в зависимости от
сроков посадки черенков in vitro
Общее накопление сырой массы и распределение ее по органам растений – регенерантов в зависимости от сезонности показало, что распределение сырой биомассы по органам (стеблям, корням) существенно менялось в зависимости от сезонности культивирования растений в среде МС in vitro. Основная доля сырой массы стеблей накапливалась при выращивании в марте-мае и в январе-феврале. Накопление основной массы корней также приходилось на эти периоды. Увеличение биомассы корней имело место также при культивировании растений–регенерантов в июле-августе при низком приросте стеблей.
Таблица 1
Зависимость столонообразования от сроков черенкования
пробирочных растений (% растений, имеющих столоны)
| Сроки черенкования | Сроки выращивания, дни | Масса клубней, г | ||||
| 15 | 30 | 40 | 50 | 60 | ||
| Столонообразование | ||||||
| Сентябрь-октябрь | | 0 | 0 | 12 | 10 | 0,155±0,017 |
| Ноябрь–декабрь | 0 | 0 | 0 | 9 | 11 | 0,147±0,012 |
| Январь– февраль | 0 | 0 | 0 | 7 | 12 | 0,214±0,018 |
| Март – апрель | 0 | 0 | 42 | 97 | 93 | 0,388±0,010 |
| Май- июнь | 0 | 0 | 37 | 97 | 98 | 0,385±0,010 |
| Июль – август | 0 | 0 | 14 | 41 | 49 | 0,242±0,015 |
Высокий уровень образования столонов также был отмечен у растений, черенкованных в начале мая и конце июня (табл.1). Столонообразование к 40 дню выращивания наблюдалось у 37% растений и выросло до 98% после 50-60 дней выращивания. Следует отметить, что растения, черенкованные в июле-августе, развивались медленнее и образовывали столоны в 2 раза меньше. Растения, черенкованные в период с сентября по начало февраля, гораздо медленнее развивались и столонообразование практически не наблюдалось. Образование столонов у этих растений было отмечено в конце вегетации и в незначительном количестве (всего у 8-12% растений).
Среднее количество столонов на одно пробирочное растение, как правило, составляло 1,2 шт., а их длина - от 3,6 до 4,2 см. Количество ветвящихся столонов было очень небольшим и составляло обычно 1 на 40 пробирочных растений.
Особенности образования столонов in vitro. Столоны брали от пробирочных растений на самом раннем этапе инициации клубнеобразования. Стерильные столоны разрезали на небольшие сегменты размером 5-6 мм и погружали апикальной или нижней частями на питательную среду. Изолированные столоны картофеля культивировали в модифицированной среде МС, с добавкой 0,1 мг/л α-нафтилуксусной кислоты (НУК) и 0,5 мг/л фолиевой кислоты, 0,1 мг/л кинетина, 3 % сахарозы при 16-часовом фотопериоде. Столоны сначала выращивали в течение 10-12 дней в темноте, затем переносили на обычный световой режим согласно методике культивирования пробирочных растений (16-часовой фотопериод). В этих условиях у сегментов столоны не развивались. Побеги начали образовываться от верхней части, находящейся над поверхностью питательной среды. Корни образовывались также из части столонов, находящихся на поверхности питательной среды. Общее количество побегов составляло около 65 %. Жизнеспособными оставались только те побеги, которые не были погружены в питательную среду.
Через 45-50 дней культивирования получались полноценные регенеранты, не отличающиеся от исходных пробирочных меристемных растений. Полученные таким образом регенеранты из столонов разрезали на сегменты с одним или двумя узлами и погружали на новую среду выращивания. При выращивании изолированных столонов было заметно образование регенерантов как из нижней, так и верхних частей сегментов. Различия в индукции столоно- и клубнеобразования имели сортовую зависимость. Столоно- и клубнеобразование было высоким у сорта Жуковский ранний (90%), а у сорта Пикассо оно не превышало 50-65%. Таким образом, столоно- и клубнеобразование имело явно выраженный сортовой признак.
По высоте растений различий не было обнаружено. Высота растений обоих сортов не превышала 10-13 см, причем, во все периоды выращивания столоновых растений. Высота растений, по-видимому, в процессе столоно- и клубнеобразования особой роли не играла. Очевидно, значимым фактором был генетический и, возможно, гормональный статус растений. Количество столонов, образовавшихся у пробирочных растений, варьировало от 2 до 4, а их длина - от 1,5 до 5 см. Число образовавшихся микроклубней соответствовало количеству столонов на пробирочных растениях. Этот признак также имел сортовой характер.
Для объяснения этого явления в последующих опытах использовали различные варианты культуральной среды МС с добавлением гормонов и углеводов.
При повышении концентрации сахарозы до 8 % и кинетина до 1,0 мг/л не было обнаружено заметного изменения образования столонов и микроклубней у обоих сортов, но наблюдалось замедление роста и уменьшение количества столонов и клубней.
У некоторых регенерантов столоны начали ветвиться. На концах части столонов образовывались по одному клубневому утолщению. У большинства пробирочных растений столоны были короткими и на них сразу образовывались клубни.
Таким образом, изменение гормонального (1 мг/л кинетина) и углеводного (8% сахарозы) баланса в среде выращивания пробирочных растений не вызывало образования дополнительных столонов и микроклубней. У обоих использованных в опытах сортов картофеля отмечено изменение морфологии регенерантов (побегов). Верхняя часть побегов была похожа на клубни. На этих средах имело место укорачивание междоузлий. Кроме того, наблюдалось утолщение междоузлий и редукция листьев. При высоких концентрациях сахарозы, кинетина и НУК наблюдалось усиление столонообразования, их длина не превышала 1,2-1,5 см. При 3 % сахарозе и 0,1 мг/л НУК или кинетина она достигала 5-6 см. Таким образом, обнаружены некоторые закономерности индукции роста и развития столонов при изменении условий культивирования растений in vitro. Нам удалось разработать эффективный способ размножения изолированных столонов картофеля.
Рост и развитие регенерантов из столонов картофеля in vitro. Для выяснения морфогенетических процессов и увеличения фотосинтетической продуктивности в последние годы используются трансгенные растения картофеля (Аксёнова и др., 2002; Гришунина и др., 2004) как перспективное направление, имеющее как фундаментальное, так и прикладное значение.
В целях изучения роли физиологических факторов в клубнеобразовании в настоящей работе приведены результаты исследования эффекта различных факторов на фотопериодическую реакцию клубнеобразования температуроустойчивых растений-регенерантов картофеля, выращенных в различных условиях фотопериода.
С этой целью исспользовали температуроустойчивые растения- регенеранты и исходную форму - сорт Жуковский ранний в качестве контрольного варианта (рис.1).
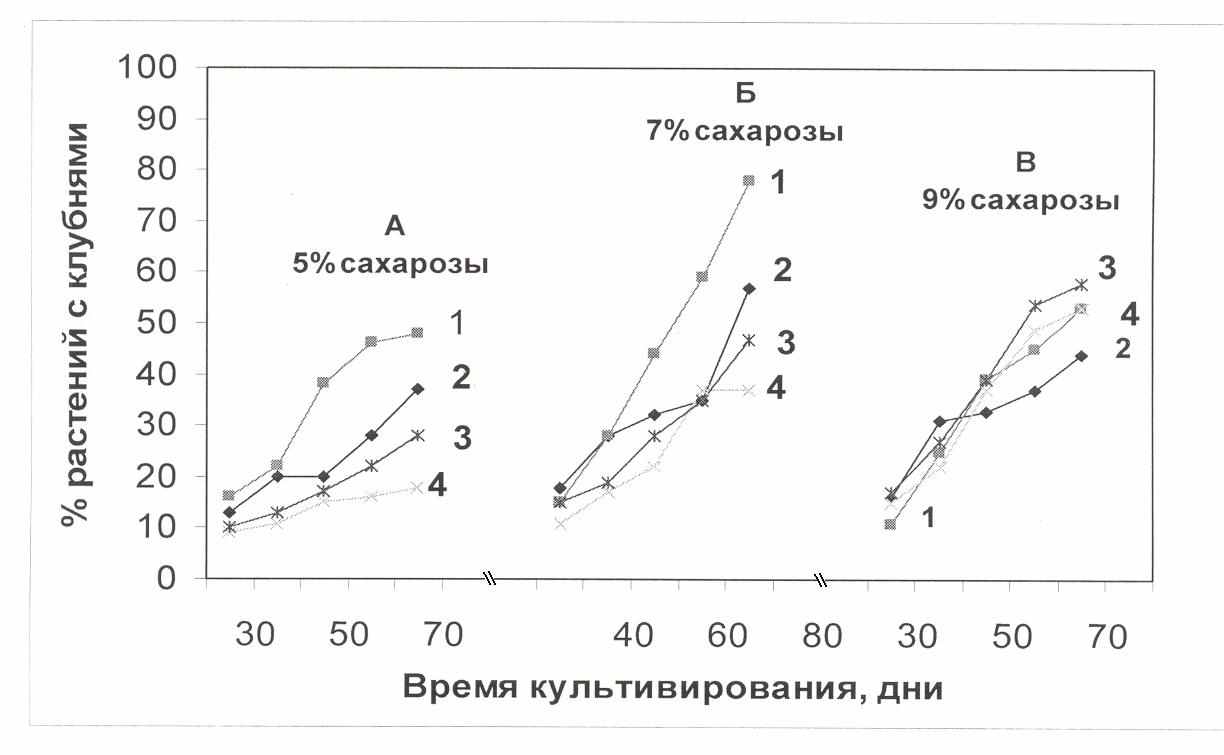
Рис.1. Динамика инициации образования клубней у регенерантов картофеля при различном содержании сахарозы в условиях длинного (ДД) и короткого (КД) дня.
1-КД, 2-ДД-исходный сорт Жуковский (контроль),
3-КД, 4-ДД-ТУ растения-регенеранты (опыт).
Контрольные растения при всех концентрациях сахарозы (5;7;9%) сформировали клубни как на ДД, так и на КД (кривые 1,2). Количество сформировавшихся клубней у этих растений на КД было выше по сравнению с растениями на ДД. При 7% -ой концентрации сахарозы контрольные растения формировали большее количество клубней, чем при 5 и 9%-ой концентрации сахарозы. Эти различия проявились чётко после 40-дневного культивирования растений. У ТУ-регенерантов наибольшая инициация клубней отмечена при 9% -ой концентрации сахарозы (кривая 4). ТУ-регенеранты проявили большую чувствительность к ингибирующему действию ДД, чем контрольные растения при такой же концентрации сахарозы (кривые 3,4). Следовательно, ТУ-регенерантам свойственна короткодневная фотопериодическая реакция клубнеобразования. Сравнение динамики инициации клубней у опытного варианта (ТУ-регенеранты) при 5;7;9 % содержания сахарозы выявило также зависимость ингибирующего действия ДД от уровня углеводного питания растений в условиях in vitro.
При низких концентрациях сахарозы (5%) в культуральной среде у ТУ-растений-регенерантов на ДД уровень клубнеобразования был очень низким (6-17 %) во все периоды культивирования растений (кривая 4). В то же время повышение содержание сахарозы до 9 % увеличивало долю растений с клубнями до 30% (кривая 4), т.е. высокое содержание сахарозы у ТУ- растений-регенерантов в условиях ДД не привело к полному снятию ингибирующего эффекта непрерывного освещения. Сравнение динамики инициации клубней у контрольного и опытного вариантов выявило различие эффекта сахарозы на ингибирующее действие ДД. Наиболее отчетливо эти различия проявились в поздние сроки культивирования. Вместе с тем, доля растений с клубнями в контрольном варианте была выше, чем в опытном во всех вариантах эксперимента (рис. 2). Сравнение образования клубней у растений обоих вариантов показало зависимость инициации клубней от фотопериода, а также от содержания сахарозы в культивируемой среде. Активирующее действие сахарозы на снятие ингибирующего эффекта ДД было выше у растений контрольного варианта, чем у ТУ - растений-регенерантов.

Рис. 2. Количество регенерантов, сформировавших клубни на 70 – й день клубнеобразования в зависимости от фотопериодического и от содержания сахарозы в культуральной среде: 1-5% сахарозы; 2-7% сахарозы; 3-9% сахарозы (сорт Жуковский и ТУ-регенеранты)
Так, количество растений с клубнями было на 20% выше в варианте КД, чем на ДД на среде 7% сахарозы в контрольном варианте. В то же время у ТУ-регенерантов влияние длины дня на клубнеобразование оказалось более зависимым от содержания сахарозы в культуральной среде. Эти растения на КД сформировали на 30% больше клубней, чем на ДД в зависимости от концентрации сахарозы. При 9% -ой концентрации сахарозы количество растений с клубнями было гораздо больше, чем при 5% и 7 % сахарозе. Следовательно, ТУ-регенеранты принципиально отличались по реакции клубнеобразования от контрольного сорта Жуковский ранний как на ДД, так и в зависимости от содержания сахарозы в культуральной среде (рис. 3).
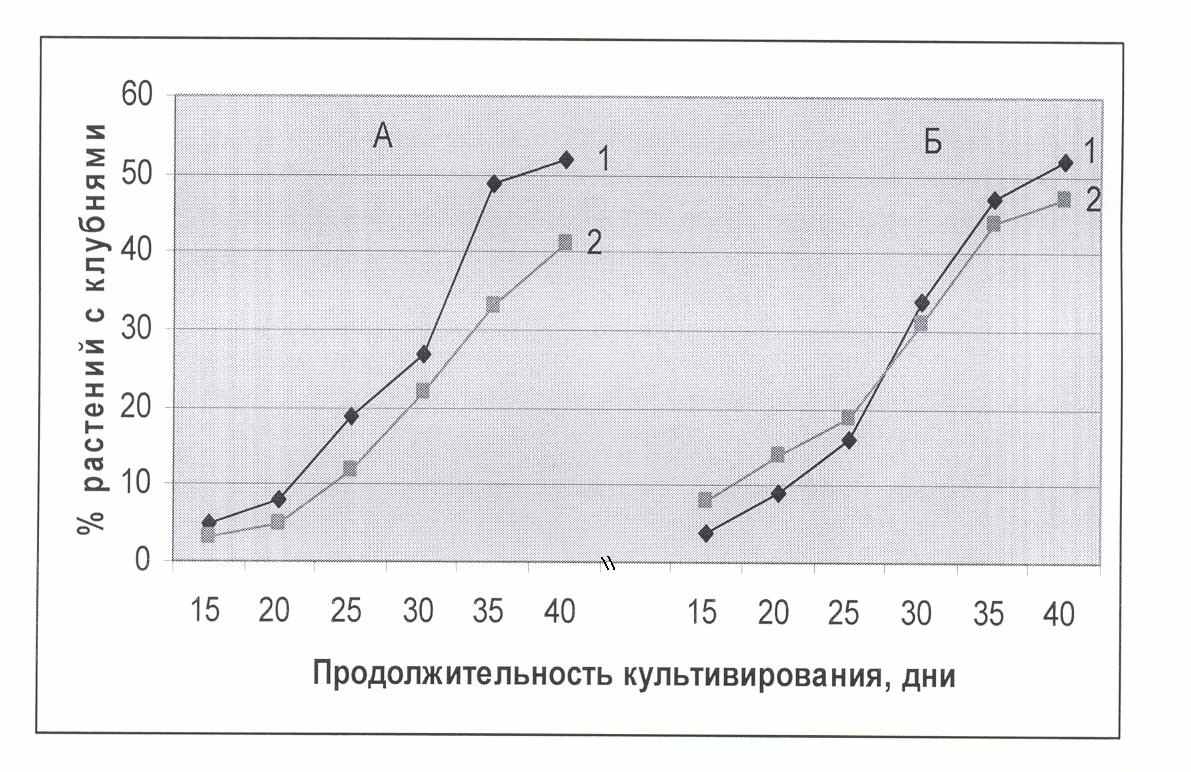
Рис.3. Динамика инициации образования клубней в условиях темноты на среде с различным содержанием сахарозы в культуральной среде на 40-й день выращивания
А - сорт Жуковский (контрольный вариант), 1- 7% сахарозы; 2- 9% сахарозы,
Б-температуроустойчивые растения-регенеранты (ТУ-регенеранты – опытный вариант), 1- 7% сахарозы; 2- 9% сахарозы.
В следующей серии опытов исследовали ростовые процессы растений в условиях разного фотопериода и при 7% и 9%-ой концентрации сахарозы в культуральной среде (табл.2). Как видно из данных таблицы, у растений контрольных вариантов больше образовалось мелких клубней, чем у ТУ- растений-регенерантов. Наблюдалось заметное увеличение общей сырой массы у растений опытных вариантов. Заметного увеличения массы клубней в зависимости от фотопериода в обоих вариантах опыта не наблюдалось. Также не была обнаружена зависимость величины массы клубней от концентрации сахарозы.
У всех опытных растений повышенное содержание сахарозы и КД вызвали увеличение сырой массы клубней. Следовательно, фотопериод оказал влияние на распределение сырой массы в пользу клубней. Доля массы клубней была несколько выше у клеточно - модифицированных растений на КД и при высокой концентрации сахарозы (9 %). Длина стебля растений в обоих вариантах опыта существенно не отличалась как на ДД, так и на КД. Концентрация сахарозы также не привела к изменению длины стебля растений.
Таблица 2
Действие ДД и КД на некоторые морфологические параметры
регенерантов картофеля в зависимости от содержания сахарозы
| Варианты | Содержа-ние сахарозы, % | Фотопериод | Средняя сырая масса, мг | Длина стебля, см | |
| клубней | растения | ||||
| Сорт Жуковский (контроль) | 7 | ДД | 104,4± 9,3 | 401,6±36,2 | 8,7±0,25 |
| | 9 | ДД | 92,3±6,4 | 332,4±50,2 | 10,3±0,5 |
| | 7 | КД | 114,2 ± 8,9 | 390,1±39,3 | 9,7±0,5 |
| | 9 | КД | 113,5±10,3 | 389,2±43,1 | 9,4±0,4 |
| ТУ-регенеран-ты (опыт) | 7 | ДД | 123,3±11,7 | 470,2±27,2 | 7,8±0,6 |
| | 9 | ДД | 136,1±10,2 | 563,4±22,4 | 8,2±0,6 |
| | 7 | КД | 122,6±9,7 | 395,4±33,3 | 8,5±0,4 |
| | 9 | КД | 161,7±14,2 | 672,4±35,2 | 9,5±0,6 |
Результаты опытов показали, что высокое содержание сахарозы в культуральной среде является основным условием инициации клубней. Культивируемые in vitro ТУ- растения - регенеранты существенно отличались от исходного сорта Жуковский ранний по ряду характерных изменений в регуляции инициации клубнеобразования. Наиболее характерным изменением в образовании клубней у ТУ - растений- регенерантов явилось усиление инициации клубней под влиянием высокой концентрации сахарозы, тогда как у контрольных растений картофеля (сорт Жуковский ранний) усиление образования клубней наблюдалось при низких концентрациях сахарозы. Необходимо отметить, что при культивировании растений обоих вариантов даже в непрерывной темноте инициация клубней ингибировалась высокой концентрацией сахарозы. Если у растений исходного варианта клубни образовывались более или менее равномерно в течение всего периода эксперимента, то у ТУ – растений - регенерантов клубни сформировались только в конце опыта. Таким образом, проведенные опыты свидетельствуют о сложности механизма клубнеобразования в зависимости от углеводного питания in vitro, фенотипическую реакцию генотипов картофеля на инициацию клубней, а также на рост вегетативных органов, которые и являются генотипическим признаком, проявляющимся даже на уровне культивирования растений картофеля in vitro.
Для выяснения этих вопросов в дальнейшем нами было изучена зависимость клубнеобразования у разных генотипов картофеля от концентрации сахарозы в культуральной среде регенерантов и полученных из них изолированных столонов картофеля.
Регуляция клубнеобразования у различных
генотипов картофеля in vitro
Результаты опытов показали, что высокое содержание сахарозы в культуральной среде является основным условием инициации клубней. Культивируемые in vitro ТУ - растения - регенеранты существенно отличались от исходного сорта Жуковский ранний по ряду характерных изменений в регуляции инициации клубнеобразования. Наиболее характерным изменением в образовании клубней у ТУ – растений - регенерантов явилось усиление инициации клубней под влиянием высокой концентрации сахарозы. Тогда как у обычных растений картофеля (сорт Жуковский ранний) усиление образования клубней наблюдалось при низких концентрациях сахарозы.
Культивирование регенерантов на среде с кинетином (0,5-1,0 мг/л) не оказывало существенного влияния на новообразование различных органов, в том числе клубней. У растений всех вариантов образовались одинаковые сходные с контрольным вариантом по форме клубни (без кинетина). Как у ТУ-регенеранта, так и у исходного сорта Жуковский ранний при культивировании регенерантов на среде, содержащей различные концентрации сахарозы (2-9%) и низкие концентрации кинетина, образовались одинаковые по форме клубни. Количество клубней на одно пробирочное растение также было одинаковым в обоих вариантах и составляло от одного до двух клубней на растение. Особой формой отличались клубни, полученные на среде с высоким содержанием сахарозы (7-9%) и 1-2 мг/л кинетина-они имели неправильную форму,причем во всех вариантах опыта.
Новообразование органов – корней, стеблей, листьев между вариантами существенно отличалось. Так, рост корней резко ингибировался на среде, содержащей высокую концентрацию сахарозы (7-9 %) и 5 мг/л кинетина, к тому же у основания растений отмечено появление каллуса и густой щетки корней. В контрольном варианте рост корней задерживался и наблюдалась короткая щетка корней, на концах которых обычно появлялись небольшие бугорки темного цвета. Рост стеблей также угнетался при высоком содержании сахарозы и кинетина. Вместе с тем, ТУ - растения - регенеранты практически не регенерировались при высоком содержании сахарозы и кинетина. У них отмечалась густая щетка корней и удлинение побегов (стеблей). Культивирование регенерантов на средах, содержащих высокие концентрации сахарозы и кинетина, стимулировало формирование «сидячих» клубней непосредственно в пазушной почке ТУ-регенеранта, тогда как у исходного сорт Жуковский ранний клубни чаще всего формировались на кончиках более развитых побегов.
Динамика накопления биомассы органов картофеля у
разных генотипов картофеля in vitro
На рис. 4 приведена динамика накопления сырой массы и распределение ее по различным органам регенерантов в зависимости от концентрации сахарозы и кинетина. Общее накопление сырой массы напрямую зависело от концентрации фитогормона и сахарозы. Кинетин существенно менял распределение биомассы по органам регенеранта. На среде с кинетином основное содержание сырой массы отмечено в стеблях, тогда как на среде с высокой концентрацией сахарозы - в клубнях.

Рис.4. Сырая масса органов регенерантов различных генотипов картофеля в течение 70 дней культивирования.
1 - сырая масса стебля,
2 - сырая масса клубня,
3 - сырая масса корней.
А - контрольный вариант (сорт Жуковский),
Б - опытный вариант (ТУ-регенерант).
Такая тенденция прослеживалась и имела место как в варианте с ТУ - растениями-регенерантами, так и у исходного сорта Жуковский ранний. Причем, у контрольного варианта количество формирующихся клубней зависело больше всего от содержания сахарозы в среде культивирования, а у ТУ-регенеранта (опытный вариант) формирование клубней зависело от наличия кинетина и более высокой концентрации сахарозы. Во всех вариантах и на испытанных культивированных средах клубни имели противоположно направленный характер изменения. Это, возможно, является следствием антагонизма в акцептировании питательных веществ между растущими стеблями и формирующимися клубнями картофеля в условиях in vitro. По всей вероятности, в гетеротрофных условиях питания (за счет культуральной среды и света) растущие органы регенеранта могут быть конкурирующими акцепторами. В последующих опытах были использованы различные концентрации сахарозы в процессе индукции столонов и инициации образования клубней. На рис. 5 показана зависимость инициации клубней (процент пробирочных растений (клонов), сформировавших клубни) от содержания сахарозы в среде выращивания, содержащей 0,5 мг/л кинетина. Как видно из приведенных данных, динамика инициации клубней на начальных этапах развития регенерантов в обоих вариантах существенно не отличалась. Средняя масса одного клубня, которая отражает скорость роста клубней у разных генотипов, существенно различалась.


Рис.5. Доля регенерантов, сформировавших клубни (А), и масса одного клубня (Б) при культивировании растений на среде с различными концентрациями сахарозы на 70-й день выращивания или в конце вегетации.
А. 1- доля растений с клубнями (контрольный вариант-сорт Жуковский),
2 - доля растений с клубнями (опытный вариант – ТУ - растение -регенерант).
Б. 1- масса одного клубня (контрольный вариант),
2 - масса одного клубня (опытный вариант).
У регенерантов опытных вариантов (ТУ - растение-регенерант) при низких концентрациях сахарозы (2%) в среде культивирования инициации клубней не происходило. При концентрации сахарозы 5% наблюдалось формирование одиночных клубней, а при 9% инициация клубней резко усиливалась. При повышении содержания сахарозы до 10% инициация образования клубней была небольшой.
В контрольных вариантах (исходный сорт Жуковский ранний) максимальное число клубней сформировалось при 5-7% концентрации сахарозы в культуральной среде. Инициация клубней у растений контрольных вариантов была существенно выше при низких концентрациях сахарозы. С дальнейшим повышением содержания сахарозы до 9%, хотя ускорялись темпы роста клубней, количество регенерантов, образовавших клубни, было ниже, чем при 5-7% концентрации сахарозы в культуральной среде. Таким образом, полученные результаты позволяют заключить, что в обоих вариантах эксперимента порог концентрации сахарозы, необходимый для инициации клубней, у разных генотипов неодинаковый. Содержание сахарозы 5-7% оказалось более благоприятным для исходного сорта Жуковский ранний (контрольный вариант), а для ТУ - растений –регенерантов (опытный вариант) порог концентрации сахарозы, инициирующий клубнеобразование, лежит между 7% и 9%. Как в контрольном, так и в опытном вариантах наблюдалось ингибирование инициации клубней при высокой концентрации сахарозы. Сравнение кривых зависимости инициации клубней и их массы от содержания сахарозы показало, что эти параметры увеличивались с повышением концентрации сахарозы. Вместе с тем, необходимо отметить различие между темпами инициации клубней и массы образовавшихся клубней в зависимости от содержания сахарозы в культуральной среде. Так, при низких концентрациях сахарозы масса клубней была ниже, чем при оптимальной. Если для контрольных растений - регенерантов оптимальной была 7%-я концентрация сахарозы, то для опытных она была выше - 9%. Анализ полученных данных позволяет высказать мысль о том, что как в контрольных, так и в опытных вариантах пороговая и оптимальная концентрации сахарозы для темпов роста клубней были сдвинуты в сторону более повышенной концентрации сахарозы по сравнению с процессами их инициации. Так, регенеранты контрольных вариантов (сорт Жуковский ранний) характеризовались более высокой способностью к инициации клубней при низких концентрациях сахарозы в культуральной среде, чем в опытных вариантах. Вместе с тем, масса одного клубня контрольных вариантов колебалась в пределах от 0,111 мг до 364 мг, против 0,076-149 мг в опытных вариантах. Сходные результаты были получены при сравнении массы образовавшихся клубней с массой стеблей в обоих вариантах.
Таким образом, полученные результаты указывают на то, что для каждого генотипа картофеля in vitro свойственна своя специфическая реакция на концентрацию сахарозы. Процессы инициации и темпы роста клубней в зависимости от концентрации сахарозы однонаправленны, но не тождественны, поскольку масса сформировавшихся клубней в заключительной фазе эксперимента и их количество во многом определялись генотипом картофеля и, следовательно, требуют разработки условий клубнеобразования для каждого сорта, линий и форм регенерантов, что имеет практическую направленность.
Заключение
Культивирование столонов картофеля in vitro открывает новые возможности для изучения физиологии клубнеобразования и поиска новых биотехнологических приемов, увеличивающих индексы урожая, а также позволяющих раскрыть механизмы гормональной и углеводной регуляции роста и развития растений.
Показана возможность использования культуры столонов in vitro для регуляции клубнеобразования картофеля. Обнаружено отсутствие сезонной зависимости клубнеобразования у пробирочных растений, индуцируемых из столонов. Отмечено изменение морфологии побегов при увеличении концентрации сахарозы и гормонов. Разработана методика культивирования столонов in vitro.
Подобраны оптимальные условия клубнеобразования у сорта Жуковский. Результаты, сведенные в табл. 3, отчетливо показывают, что количество образующихся клубней повышается при добавлении в культуральную среду 2-5% -го раствора сахарозы в сочетании с НУК и кинетином. В этих условиях масса клубней также выше, чем в вариантах с использованием других сред. Таким образом, среды 6 и 7 можно рекомендовать для получения микроклубней в системе in vitro. Добавление ИУК в сочетании с кинетином также не стимулирует процесс инициации роста клубней.
Столоновые растения по ряду морфологических признаков-количество и длина междоузлий, размер листовой пластинки-сходны с меристемными растениями, но по накоплению общей биомассы и времени иннициации клубней проявились существенные различия (табл.4).
Таблица 4
Морфологическая характеристика меристемных (контрольных)
и столоновых растений картофеля сорта Жуковский-ранний
| Параметры | Контрольные растения | Столоновые растения |
| Количество междоузлий, шт. | 5,0 ± 1,3 | 5,1 ± 1,2 |
| Длина междоузлий, см | 1,4 ± 0,4 | 1,2 ± 0,3 |
| Общая площадь листьев, см² | 24,3 ± 3,9 | 23,7 ± 2,8 |
| Сырая масса, (листья, стебель, корень), г | 2,7 ± 0,8 | 3,4 ± 1,1 |
| Время корнеобразования (дни после черенкования) | 14,8 ± 2,1 | 8,4 ± 1,7 |
| Время инициации клубней (дни после черенкования) | 34,3 ± 5,9 | 22,7 ± 4,4 |
Так, для столоновых регенерантов характерно более быстрое формирование корневой системы и инициация клубней по сравнению с меристемными растениями. Столоновые растения, выращенные in vitro, в отличие от меристемных растений (контрольных) характеризуются более быстрым корнеобразованием и более быстрым накоплением биомассы (за счет корней), что может быть обусловлено изменением в уровне усвоения углеводов. По другим морфогенетическим параметрам, таким, как количество и длина междоузлий, общая площадь листьев, между столоновыми и меристемными регенерантами существенных различий не обнаружено.
Таблица 3
Продолжительность вегетации и продуктивность столоновых регенерантов картофеля
сорта Жуковский ранний в зависимости от условий культивирования in vitro
| Варианты опыта | Продолжи-тельность вегетации, дни | Формиро-вание клубней, % | Сырая масса побегов мг/растение | Количество клубней на растении, шт. | Общая сырая масса клубней, мг | Масса одного клубня, мг | Индекс урожая |
| Контроль (среда МС, 2 % сахароза, 1 мг/л ИМК) | 85±7 | 63 | 443±71 | 1,3 | 162±22 | 24±212 | 0,4 |
| Среда МС, 2 % сахароза, 1 мг/л ИМК 1 мг/л кинетина | 87±5 | 65 | 511±65 | 1,8 | 450±29 | 250±27 | 0,8 |
| Среда МС, 5 % сахароза, 0,1 мг/л кинетина, 1 мг/л ИМК | 80±7 | 74 | 506±7 | 2,2 | 607±32 | 176±43 | 1,2 |
| Среда МС, 7 % сахароза, 0,5 мг/л кинетина, 1 мг/л ИМК | 78±9 | 82 | 507±71 | 2,2 | 539±34 | 245±43 | 1,1 |
| Среда МС, 2 % сахароза. 1 мг/л ИМК 1 мг/л НУК | 78±4 | 88 | 717±5 | 2,0 | 294±23 | 47±14 | 0,4 |
| Среда МС, 5 % сахароза, 1 мг/л НУК, 0,5 мг/л кинетина | 73±8 | 87 | 793±77 | 3,1 | 1165±217 | 376±19 | 1,6 |
| Среда МС, 7 % сахароза, 1 мг/л НУК, 0,5 мг/л кинетин | 78±4 | 89 | 743±85 | 3,4 | 1123±48 | 362±32 | 1,6 |
Можно предположить, что различия в морфофизиологических параметрах у столоновых и меристемных регенерантов связаны с экспрессией генов гексокиназного цикла метаболизма углеводов, особенно фермента инвертазы, которая осуществляет конверсию сахарозы в глюкозу и фруктозу. Задержка этого процесса и накопление в среде культивирования альдегида-глюкозы может оказать сильное влияние на процесс дифференцировки меристемных клеток, поскольку может проявиться мутагенный эффект, что повлечет за собой появление геномных и цитоплазматических изменений и что не наблюдается у столоновых регенерантов. Это может привести к улучшению инициации и роста клубней, что наблюдается у столоновых растений – регенерантов.
Другая важнейшая особенность столоновых регенерантов, на наш взгляд, это сокрашение срока получения базисного семенного материала картофеля. Цикл получения базисного семенного материала у меристемных регенерантов доходит до 16 месяцев, а у столоновых примерно на 4 месяца меньше, т.е. составляет 12 месяцев.
Дальнейший прогресс в этом направлении во многом зависит не только от использования различных методических приёмов, но и более полного изучения особенностей культуры столонов и регуляции роста клубнеобразования в конкретных агрофизиологических условиях, что дает возможность разработать новые схемы селекции и семеноводства картофеля в Республике Таджикистан.
Выводы
- Разработан достаточно простой и эффективный способ культивирования изолированых столонов картофеля in vitro. Выявлены некоторые особенности индукции и роста столонов картофеля при изменении условий культивирования растений-регенерантов. Наиболее эффективной оказалась среда с добавлением 5-7% концентрации сахарозы и НУК (1,0 мг/л) в сочетании с кинетином (0,5 мг/л). В этих условиях культивирования столоны давали полноценные регенеранты с высокой степенью морфогенетической потенции.
- Показано, что высокое содержание сахарозы в культуральной среде является основным условием инициации микроклубнеобразования. Культивируемые in vitro температуроустойчивые регенеранты (ТУ-регенеранты) существенно отличались от исходного сорта Жуковский ранний по ряду характерных изменений в регуляции клубнеобразования. Активизируется инициация клубнеобразования под влиянием высокой концентрации сахарозы и продолжительности периода роста клубня, тогда как у исходного сорта Жуковский ранний инициация клубнеобразования наблюдалась при низких концентрациях сахарозы и более коротких промежутках времени. Это свидетельствует о том, что инициация и рост клубней это генотипически детерменированный процесс, проявляющийся даже в условиях in vitro.
- Установлено влияние фотопериода (ДД и КД) на инициацию, рост клубнеобразования и распределение сырой массы независимо от генотипа картофеля. Клубнеобразование было больше у ТУ-регенерантов на КД и при высокой концентрации сахарозы. Фотопериод в условиях in vitro не привел к изменению морфогенетических признаков растений-регенерантов как у ТУ-регенерантов, так и у исходного сорта Жуковский ранний.
- Установлено, что как у исходного сорта Жуковский ранний, так и у ТУ-регенерантов пороговая и оптимальная концентрации сахарозы для скорости роста были сдвинуты в сторону более повышенной концентрации сахарозы по сравнению с процессами инициации регенерантов столонов. Регенеранты исходного сорта Жуковский ранний характеризовались более высокой способностью к инициации клубней при низких концентрациях сахарозы в среде культивирования, чем у ТУ-регенерантов.
- Полученные результаты указывают на то, что каждому генотипу картофеля in vitro свойственна своя, специфическая реакция на условия культивирования. Процессы инициации роста клубней в зависимости от содержания углеводов однонаправленны, но не одинаковы. Очевидно, этот процесс контролируется группой независимых генов, экспрессия которых является важнейшим фактором регуляции клубнеобразования, роста и развития растений.
Список работ, опубликованных по теме диссертации
- Алиев К.А., Давлятназарова З.Б., Назарова Н.Н., Мирзохонова Г.О. Физиологические особенности регенерации растений картофеля. Международный симпозиум «Физиология трансгенного растения и проблемы биобезопасности» (Transgenic plants and biosafety), Москва, 2004. Тезисы докладов. С. 23-24.
- Давлятназарова З.Б., Каримов Б.К., Авгонова Х.Х., Мирзохонова Г.О., Назарова Н.Н., Алиев К.А. Регуляция клубнеобразования in vitro. Материалы научной конференции «Актуальные проблемы и перспективы развития физиологии растений». Душанбе; Дониш, 2004. С. 62-63.
- К.А. Алиев, Н.Н. Назарова, Г.О. Мирзохонова, Х.Х. Авганова, М.А. Алиев. Клеточно-модифицированные растения картофеля как модель изучения эффекта цитокининов на продуктивность in vitro. Материалы межд. конференции «Физиологические и молекулярно-генетические аспекты сохранения биоразнообразия», 19-23 сентября, Вологда, 2005. С.7.
- Давлятназарова З.Б., Мирзохонова Г.О., Назарова Н.Н., Алиев К.А. Регуляция клубнеобразования у различных генотипов картофеля in vitro. Материалы Респ. симпозиума «Экономика и наука Горно-Бадахшанской автономной области: прошлое, настоящее, будущее». Хорог, 2005. С.187-189.
- Назарова Н.Н., Давлятназарова З.Б., Мирзохонова Г.О., Каримов Б.Б., Алиев К.А.. Образование столонов и микроклубней картофеля в зависимости от сроков посадки in vitro. Докл. АН РТ, 2004, №11-12. С.92-101
- Давлятназарова З.Б., Назарова Н.Н., Мирзохонова Г.О., Каримов Б.К., Эсаналиева Ш.А., Алиев К.А. Рост и клубнеобразование регенерантов картофеля в зависимости от условий выращивания in vitro. Докл. АН РТ, 2004, №11-12. С.79-92.
- Назарова Н.Н., Мирзохонова Г.О., Алиева С.К., Каримов Б.К.,Алиев К.А. Некоторые особенности образования столонов in vitro. Известия АН РТ, 2005, №3-4(153). С. 36-39.
- Мирзохонова Г.О., Назарова Н.Н.,Давлятназарова З.Б., Алиев К.А. Регуляция клубнеобразования у различных генотипов картофеля in vitro. Известия АН РТ, 2005, № 3-4 (153). С.40-44.
- Мирзохонова Г.О., Назарова Н.Н.,Давлятназарова З.Б., Каримов Б.К., Алиев К.А. Гормональная регуляция инициации и роста клубней регенерантов картофеля in vitro. Известия АН РТ, 2005, № 3-4 (153). С.45-51.