
Бактерии-деструкторы ароматических углеводородов и их хлорпроизводных: разнообразие, особенности метаболизма, функциональная геномика 03. 02. 03 Микробиология
| Вид материала | Автореферат диссертации |
- Рабочей программы учебной дисциплины (модуля) микробиология, 48.16kb.
- Твердофазная люминесценция полициклических ароматических углеводородов в условиях адсорбционного, 262.95kb.
- Задание егэ по теме «Углеводороды» а задания, 45.66kb.
- «Взаимосвязь предельных, непредельных и ароматических углеводородов», 11.54kb.
- Свистунова наталья Юрьевна биологические особенности лекарственных и ароматических, 416.08kb.
- Вопросы для подготовки к зачету по дисциплине: «Эксплуатационные материалы», 51.34kb.
- Оценка фотодинамического воздействия in vitro на бактерии из микробоценозов ротовой, 331.98kb.
- Функциональная и рецепторная характеристика белков суперсемейства иммуноглобулинов, 1127.13kb.
- Бактерии рода enterococcus в озере байкал: Распределение, видовой состав, механизм, 275.08kb.
- Ароматические углеводороды. Бензол представитель аренов. Строение молекулы и физические, 54.51kb.
На правах рукописи
ПЛОТНИКОВА Елена Генриховна
Бактерии-деструкторы ароматических углеводородов
и их хлорпроизводных: разнообразие, особенности метаболизма, функциональная геномика
03.02.03 Микробиология
Автореферат диссертации на соискание ученой степени
доктора биологических наук
Пермь - 2010
Работа выполнена в лаборатории химического мутагенеза Института экологии и генетики микроорганизмов УрО РАН, Пермь
Научные консультанты:
доктор медицинских наук,
чл. – корр. РАН, профессор Демаков Виталий Алексеевич
доктор биологических наук Евтушенко Людмила Ивановна
Официальные оппоненты:
доктор биологических наук Октябрьский Олег Николаевич
доктор биологических наук Донова Марина Викторовна
доктор медицинских наук Несчисляев Валерий Александрович
Ведущая организация: Институт микробиологии им. С.Н. Виноградского РАН, Москва
Защита состоится «__»__________ 2010 г. в ___ часов на заседании диссертационного совета ДМ 004.019.01 в Институте экологии и генетики микроорганизмов УрО РАН по адресу: 614081, г. Пермь, ул. Голева, д. 13. Факс: (342) 2446711.
С диссертацией можно ознакомиться в библиотеке Института экологии и генетики микроорганизмов УрО РАН.
Автореферат разослан «__ » ___________ 2010 г.
Ученый секретарь диссертационного совета,
кандидат биологических наук Максимова Юлия Геннадьевна
Общая характеристика работы
Актуальность проблемы. Проблема очистки наземных и водных экосистем, загрязненных токсичными, устойчивыми к разложению и представляющими опасность для здоровья человека химическими соединениями, занимает центральное место в ряду актуальных задач современной экологии. Среди поллютантов, обладающих канцерогенными и мутагенными свойствами, а также тенденцией к биоаккумуляции, наиболее распространенными являются полициклические ароматические углеводороды (ПАУ), бифенил и его хлорированные производные (Hall, Grover, 1999; ATSDR, 2000). ПАУ поступают в окружающую среду с разливами нефти, отходами коксохимических, газоперерабатывающих производств и химических предприятий, образуются при неполном сгорании топлива и бытовых отходов (Seo et al., 2009). Полихлорированные бифенилы (ПХБ) широко применялись в течение нескольких десятилетий при производстве лакокрасочных материалов, пластификаторов, пестицидов, в электротехнической промышленности. В настоящее время производство и использование ПХБ, как особо стойких органических загрязнителей, запрещено Стокгольмской Конвенцией (ссылка скрыта). Наряду с задачами по восстановлению экосистем, загрязненных этой группой токсикантов, остро стоит проблема детоксикации больших объемов невостребованных коммерческих смесей, созданных на основе хлорбифенилов (Васильева, Стрижакова, 2007; Pieper, Seeger, 2008). Биоремедиация почв и других объектов окружающей среды с использованием микроорганизмов-деструкторов (хлор)ароматических соединений имеет ряд известных преимуществ по сравнению с другими методами экобиотехнологии (Sutherland et al., 1995; Pieper, 2005; Cao et al., 2009).
К настоящему времени обнаружены и описаны бактерии различных эволюционных групп, способные разлагать нафталин, фенантрен, антрацен, другие высокомолекулярные ПАУ (Боронин и др., 1989; Бабошин и др., 2005; Kanaly, Harayama, 2000; Jones et al., 2003; Reng et al., 2008). Способность к трансформации ПХБ описана для широкого круга бактерий – протеобактерий (Acinetobacter, Alcaligenes, Cupriavidus, Pseudomonas, Sphingomonas), актинобактерий (Rhodococcus, Arthrobacter), споровых (Bacillaceae) (Bedard, Haberl, 1990; Furukawa, Fujihara, 2008; Pieper, Senger, 2008). Однако известны лишь единичные штаммы, исключительно представители протеобактерий (Burkholderia, Enterobacter, Pseudomonas и Ralstonia), которые могут полностью разлагать моно- и дихлорбифенилы (Kim, Picardal, 2001; Adebusoye et al., 2008).
Большинство известных штаммов-деструкторов трансформируют ПХБ до соответствующих хлорбензойных кислот (ХБК). Последующие этапы деградации хлорбензоатов в ПХБ-деградирующих микробных системах осуществляют обычно другие группы организмов (Unterman, 1996). Среди них значительная роль принадлежит бактериям, осуществляющим отщепление галогена на первых этапах разложения хлорбензоатов, в результате чего образуются менее токсичные соединения, доступные для использования в качестве субстрата для многих почвенных микроорганизмов (Карасевич, 1982; Scholten et al., 1991; Fetzner, 1998; Field, Sierra-Alvarez, 2008).
Развитие молекулярно-генетических методов и исследования в области структурной и функциональной геномики бактерий-деструкторов способствовало пониманию молекулярных механизмов биодеградации вышеназванных соединений. Показано, что генетические детерминанты (гены/опероны) бактериальной деструкции ПАУ, бифенила/ПХБ и ХБК локализованы в хромосоме и высокомолекулярных плазмидах, и могут присутствовать в клетке в нескольких копиях (Boronin, 1992; Sanseverino et al., 1993; Reineke, 1998; Shimizu et al., 2001; Dennis, Zylstra, 2004; Basta et al., 2005; Chain et al., 2006; Iwasaki et al., 2006). Имеются сведения о горизонтальном переносе генов биодеградации в составе мобильных генетических элементов (плазмид, транспозонов, интегронов), в том числе между бактериями различных филогенетических групп (Fulthorpe, Campbell, 1992; Herrick et al., 1997; Nishi et al., 2000; Gartemann, Eichenlaub, 2001; Habe, Omori, 2003; Reng et al., 2008). Установлена важная роль диоксигеназ на начальных этапах аэробной деструкции ПАУ и бифенила/ПХБ (McKay et al., 1997; Gibson, Parales, 2000; Seeger et al., 2001; Jakoncic et al., 2007; Schuler et al., 2009). Ферменты этой группы способны осуществлять трансформацию многих полиароматических соединений, что определяет их важную роль в развитии различных биотехнологий (Harayama, 1997; Boyd et al., 2001; Bamforth, Singleton, 2005; Shindo et al., 2007; Cao et al., 2009).
Функциональные особенности бактерий-деструкторов в комплексе с данными по структурной и функциональной геномике находятся в фокусе внимания современной микробной экологии, а также активно развивающейся системной биологии. Концепции системной биологии и синергия экологических подходов и технологий геномики, метагеномики, протеомики и метаболомики являются, с точки зрения современной науки, наиболее перспективным подходом к пониманию механизмов адаптации и (микро)эволюции индивидуальных микроорганизмов и микробных сообществ к условиям изменяющейся природной среды, особенно в условиях многофакторного негативного воздействия (Trigo et al., 2009). Изучение микроорганизмов, обитающих в определенных эколого-географических условиях и подверженных одновременному воздействию различных антропогенных факторов (в т.ч. высокий уровень токсичных поллютантов и повышенная минерализация среды), вызывает особый интерес. Актуальность таких исследований, наряду с теоретическими аспектами, обусловлена также и практическими задачами экобиотехнологии. Основной среди них является разработка новых стратегий биоремедиации, в том числе в связи с неэффективностью технологий, созданных на основе не адаптированных к соответствующим условиям микроорганизмов.
Имеющиеся сведения о микроорганизмах, способных к деструкции полиароматических углеводородов в условиях высокой солености, немногочисленны и касаются, в основном, грамотрицательных или «морских» бактерий родов Pseudomonas, Сycloclasticus, Vibrio, Lutibacterium, Neptunomonas (Garsia-Valdes et al, 1988; Dyksterhouse et al., 1995; Geiselbrecht et al., 1998; Hedlud et al., 1999, 2001; Chung, King, 2001; Kasai et al., 2002; Garcýa et al., 2005; Abed et al., 2006), а также спорообразующих бактерий (Paenibacillus) (Daane et al., 2002). Крайне слабо изучены механизмы адаптации бактерий к таким условиям существования (Roberts, 2005; Oren, 2008), а также взаимодействия внутри микробных сообществ: регуляция их состава, метаболической активности и устойчивости к высокому содержанию солей. До начала наших работ практически отсутствовали сведения о разнообразии и функциональных особенностях грамположительных бактерий - резидентов наземных экосистем, в которых ведущими факторами формирования микробиоценозов являются соленость среды и токсичный поллютант. Все вышесказанное определяет актуальность и перспективность фундаментальных и прикладных исследований в этом направлении.
Цель и задачи исследования. Целью работы было исследование таксономического, генетического, функционального разнообразия аэробных бактерий-деструкторов ПАУ и ПХБ, а также создание и изучение бактериальных штаммов и ассоциаций с заданным биодеградативным потенциалом.
Основные задачи исследования:
- Выделение бактерий-деструкторов нафталина, фенантрена, бифенила/ПХБ, ХБК из почв/грунтов, загрязненных отходами химических и соледобывающего производств.
- Изучение генотипических и фенотипических характеристик изолятов и определение их таксономического положения с использованием принципов полифазной таксономии.
- Характеристика экологических особенностей и биодеградативного потенциала выделенных бактерий.
- Изучение разнообразия ключевых генов деструкции нафталина, бифенила/ПХБ и ХБК у бактерий.
- Конструирование метаболических путей деструкции хлорированных соединений на уровне бактериальной клетки и ассоциации бактерий.
- Выделение и изучение галотолерантных нафталинметаболизирующих консорциумов бактерий.
Научная новизна. Получены новые сведения о разнообразии микробных сообществ, обитающих в условиях повышенной солености, высокого уровня загрязнения среды токсичными поллютантами. Детально охарактеризовано 90 штаммов-деструкторов ПАУ, бифенила/ПХБ и ХБК - представителей классов Proteobacteria (роды Pseudomonas, Alcaligenes, Flavobacterium), Actinobacteria (Arthrobacter, Brevibacterium, Cellulomonas, Microbacterium, Rhodococcus, Dietzia, Janibacter, Kocuria, Streptomyces) и семейства Bacillaceae (Bacillus, Paenibacillus). Выявлены бактерии, разлагающие токсиканты в широком диапазоне температур (от +4 до 40ºС), рН среды (от 5 до 9) и при повышенной солености среды (до 90 г/л NaCl). Впервые описаны грамположительные бактерии, осуществляющие полное разложение ряда моно- и дихлорбифенилов: Microbacterium sp. В51 – орто моно(ди)ХБ; Rhodococcus ruber Р25 (=ИЭГМ 896) – (орто)пара-моноХБ и 2,4’диХБ. Впервые выделены в культуру и охарактеризованы галотолерантные бактерии-деструкторы ПАУ (нафталина и фенантрена), относящиеся к родам Rhodococcus, Arthrobacter, Bacillus, Paenibacillus и Pseudomonas. На основании филогенетической обособленности и фенотипических отличий от ближайших таксонов валидно описаны новый род Salinicola, включающий вид S. socius, а также новые виды рода Brevibacterium – В. permense, B. antiquum и В. aurantiacum; выявлены новые виды «Rhodococcus naphthalenivorans» sp. nov., «Thalassospira permensis» sp. nov. и ряд других видов.
Обнаружены и изучены ключевые гены начального этапа разложения бифенила/ПХБ у бактерий-деструкторов родов Arthrobacter, Janibacter, Rhodococcus. Впервые клонированы и изучены fcb-гены A. globiformis КЗТ1, контролирующие гидролитическое дегалогенирование 4ХБК. Путем клонирования fcb-генов в клетки P. putida впервые сконструирован рекомбинантный путь полной деградации 4ХБК, более эффективный в отношении утилизации субстрата в сравнении с природными штаммами. В результате селекции на высоких концентрациях 4ХБК рекомбинантного штамма P. putida КЗ-6R (pPC3) получены супердеструкторы 4ХБК. Впервые у бактерий рода Rhodococcus обнаружены и охарактеризованы fcb-гены, кодирующие компоненты 4ХБК-дегалогеназы. Клонированы и изучены новые ohb-гены P. aeruginosa 142, контролирующие окислительное орто-дегалогенирование 2-/2,4-хлорбензоатов. ohb–Гены фланкированы IS1396-подобными последовательностями, содержащими предполагаемые гены транспозазы А (tnpA) и ДНК топоизомеразы I/III (top).
Впервые выделены и исследованы ассоциации бактерий, способные к росту и утилизации нафталина в условиях высокой солености среды (до 8 - 9 % NaCl). Показано, что в состав ассоциаций входят галотолерантные бактерии-деструкторы ПАУ порядка Actinomycetales, а также галотолерантные и умеренно галофильные бактерии, не способные к разложению нафталина. Экспериментально подтверждено присутствие протокооперативных взаимоотношений в консорциуме SMB3 между деструкторами нафталина и бактериями-спутниками. Впервые получены данные о положительном влиянии экзогенных осмопротекторных соединений, продуцируемых галофильными/галотолерантными бактериями исследуемых микробных ассоциаций, на разложение ПАУ штаммами-деструкторами в условиях повышенной солености среды. Для представителей рода Rhodococcus (деструкторов нафталина из консорциума SMB3) впервые описана комбинация осмопротекторов: глутамат, эктоин и гидроксиэктоин.
На основе выделенных и детально охарактеризованных штаммов-деструкторов ПХБ и ХБК предложены и экспериментально обоснованы новые подходы к созданию бактериальных ассоциаций, способных эффективно осуществлять утилизацию различных конгенеров ПХБ (как индивидуальных, так и в составе коммерческих смесей).
Теоретическое и практическое значение работы. Полученные данные о способности аэробных бактерий разлагать ароматические соединения при повышенной солености среды расширяют представления о системах биодеградации ПАУ и могут быть использованы для создания новых эффективных биотехнологий восстановления окружающей среды. Выделенные и детально охарактеризованные уникальные штаммы-деструкторы ПХБ, в частности, Rhodococcus ruber Р25 (=ИЭГМ 896) (Патент РФ №2262531), могут применяться для восстановления почв и водоемов, загрязненных ПХБ, а также в технологиях детоксикации промышленных смесей, содержащих хлорбифенилы. Полученные путем селекции галотолерантные нафталинметаболизирующие консорциумы являются перспективными для биоремедиации загрязненных ПАУ почв и водоемов в условиях засоления. Данные по механизмам функционирования консорциумов могут быть использованы для усовершенствования методов биоремедиации засоленных экосистем. Результаты изучения D-плазмид галотолерантных бактерий-деструкторов найдут применение при конструировании рекомбинантных штаммов для создания биотехнологий ремедиации антропогенных экосистем.
Конструирование рекомбинантного пути полной деградации 4ХБК в клетках P. putida определило новые подходы к созданию штаммов микроорганизмов, способных к эффективной деструкции и/или детоксикации различных ксенобиотиков-поллютантов. Полученные рекомбинантные штаммы и плазмиды, содержащие fcb- и ohb-гены, были использованы в совместных исследованиях в Центре микробной экологии Мичиганского университета (г. Лансинг, США). На основе fcb- и ohb-генов был создан ряд генноинженерных штаммов, осуществляющих полную утилизацию орто- и пара-хлорбифенилов. Полученные рекомбинантные штаммы-деструкторы хлорированных бензойных кислот и бифенилов могут быть использованы при создании инновационных технологий защиты окружающей среды от токсичных галогенароматических соединений.
Новые данные о разнообразии бактерий и генетическом потенциале микробного сообщества района солеразработок (г. Березники, Пермский край) являются основой для разработки критериев мониторинга этого уникального микробиоценоза. Результатом изучения таксономического разнообразия явились усовершенствованные системы классификации и идентификации бактерий ряда групп - важными для микробиологической практики и в связи с вопросами интеллектуальной собственности.
Создана обширная рабочая коллекция бактерий-деструкторов (галоген)арома-тических соединений и галотолерантных/галофильных бактерий, доступных для фундаментальных и научно-прикладных исследований специалистами разного профиля. Большинство штаммов включены в фонд Всероссийской коллекции микроорганизмов (ВКМ, ИБФМ, г. Пущино) и Региональной профилированной коллекции алканотрофных микроорганизмов (ИЭГМ УрО РАН, г. Пермь). Дубликаты типовых штаммов описанных новых видов депонированы в ВКМ и Биологических ресурсных центрах мира (в Германии, Бельгии, Франции, Японии), а также включены в международную поисковую систему Straininfo (ссылка скрыта). Сведения о последовательностях генов 16S рРНК и функциональных генов (fcb, ohb, har, bph) штаммов-деструкторов включены в GenBank (Национальный центр биологической информации, США; http:/www.ncbi.nlm.nih.gov) и сопряженные базы данных (EMBL, DDBJ, EzTaxon) других стран.
Материалы диссертации используются в лекционных курсах Биологического факультета (на Кафедрах микробиологии и иммунологии; ботаники и генетики растений) Пермского государственного университета.
Основные положения, выносимые на защиту
- Бактерии родов Arthrobacter, Rhodococcus, Bacillus, Paenibacillus и Pseudomonas преобладают среди культивируемых аэробных галотолерантных бактерий-деструкторов ПАУ из микробиоценозов района разработок Верхнекамского месторождения солей (г. Березники, Пермский край). Бактерии содержат разнообразные генетические детерминанты (D-плазмиды, гены/опероны), контролирующие ключевые этапы разложения нафталина и фенантрена.
- Полное разложение ряда моно- и дихлорбифенилов (феномен ранее не известный для грамположительных бактерий) осуществляют штаммы Microbacterium sp. В51 [(орто-моно(ди)ХБ] и Rhodococcus ruber Р25 [(орто)пара-моноХБ и 2,4’диХБ]. Другие активные штаммы бактерий характеризуются разной окислительной способностью по отношению к орто- и пара-хлорированным бифенилам. Бактерии-деструкторы моно(ди)хлорбифенилов, относящиеся к родам Arthrobacter, Rhodococcus, Janibacter и Pseudomonas, содержат структурно различающиеся гены, контролирующие начальный этап разложения бифенила/ПХБ.
- Впервые клонированы и изучены fcb-гены A. globiformis KЗТ1, контролирующие реакцию гидролитического дегалогенирования 4ХБК. В клетках P. putida с использованием fcb-генов сконструирован рекомбинантный путь полной деградации 4ХБК и получены супердеструкторы, характеризующиеся высокой стабильностью признака утилизации этого соединения. Клонированы и охарактеризованы новые ohb-гены P. aeruginosa 142, контролирующие окислительное орто-дегалогенирование 2-/2,4-хлорбензоатов.
- В условиях повышенной солености среды (до 8-9% NaCl) эффективная деструкция и утилизация в качестве источника углерода нафталина может осуществляться консорциумами бактерий, включающими галотолерантные бактерии-деструкторы ПАУ порядка Actinоmycetales, а также галотолерантные и умеренно галофильные бактерии, не способные к разложению нафталина. В галотолерантных нафталинметаболизирующих ассоциациях микроорганизмов присутствуют протокооперативные взаимоотношения.
- В составе микробиоценозов района разработок Верхнекамского месторождения солей присутствуют галофильные и галотолерантные бактерии новых таксонов.
Публикации. По материалам диссертации опубликовано 110 печатных работ, включая 24 экспериментальные статьи в реферируемых зарубежных, центральных и региональных журналах, 1 обзор, 1 патент, 1 учебное пособие (в соавторстве), 12 статей в сборниках научных статей, 70 тезисов докладов и сообщений на российских и международных конференциях и симпозиумах.
Объем и структура диссертации. Работа изложена на 310 страницах машинописного текста, содержит 30 таблиц и 50 рисунков. Диссертация состоит из введения, обзора литературы, описания материалов и методов исследования, пяти глав экспериментальных исследований, отражающих результаты исследований, а также заключения, выводов, списка цитируемой литературы, включающего 560 литературных источников, из них 60 на русском и 500 на английском языках. В Приложении приведены нуклеотидные последовательности генов 16S рРНК и функциональных генов (fcb, ohb, har, bph) исследуемых штаммов, депонированных в международной базе данных GenBank.
Связь работы с крупными научными программами. Работа выполнена в соответствии с планом НИР ИЭГМ УрО РАН, тема «Биохимические и генетические системы трансформации сложных органических соединений у бактерий, перспективных для биотехнологии» (№ гос. рег. 0120.0406511), а также в рамках Программ фундаментальных исследований Президиума РАН «Молекулярная и клеточная биология» и «Фундаментальные основы воспроизводства и оптимизации генофондов растений, животных и человека»; Программы интеграционных фундаментальных исследований Президиума УрО РАН, выполняемых в содружестве с учеными СО РАН; проектов РФФИ №00 04 49056-а, №02-04-49043-а, РФФИ-Урал №02-04-96404_а, №04-04-96042_а, №07 04 96078_а, №07 04 97625 р_офи.
Апробация работы и публикации. Материалы диссертации были представлены и обсуждены на международных симпозиумах и конференциях: III Международном симпозиуме «Pseudomonas» (Триест, 1991), Конференции Американского общества микробиологов «Анаэробное дегалогенирование» (Атенс, 1992), Съездах Американского общества микробиологов (1993; 1994; 1996), X Международном конгрессе по бактериологии и прикладной микробиологии (Париж, 2002), I Конгрессе Европейского микробиологического общества (Любляна, 2003), Международных конференциях «Загрязнение окружающей среды» (Москва, 1998; Волгоград-Пермь, 2001; Пермь, 2008), IV и V Международных семинарах-презентациях инновационных научно-технических проектов «Биотехнология-2000» и «Биотехнология-2001» (Пущино, 2000; 2001), Международном семинаре «Научно-технический потенциал Западного Урала в области конверсии военно-промышленного комплекса» (Пермь, 2001), Международном Байкальском симпозиуме «Микроорганизмы в экосистемах озер, рек, водохранилищ» (Иркутск, 2003), Международной конференции «Биохимические взаимодействия микроорганизмов и растений с техногенными загрязнителями окружающей среды» (Саратов, 2003), II и III Международных конференциях «Микробное разнообразие: состояние, стратегия сохранения, биологический потенциал» (Пермь, 2005; 2008), Международной конференции «Проблемы биодеструкции техногенных загрязнителей окружающей среды» (Саратов, 2005), IV и V Международных конгрессах «Биотехнология: состояние и перспективы развития» (Москва, 2005, 2007), Международной научной конференции «Микроорганизмы и биосфера» (Москва, 2007), Международных научных конференциях «Современное состояние и перспективы развития микробиологии и биотехнологии» (Минск, 2004, 2008), V съезде Вавиловского общества генетиков и селекционеров (Москва, 2009), Международной научной конференции «Актуальные проблемы микробиологии и биотехнологии» (Кишинев, 2009) и других.
Благодарность. Автор выражает искреннюю благодарность и признательность научному консультанту, заведующему лабораторией химического мутагенеза ИЭГМ УрО РАН, чл.-корр. РАН В.А. Демакову - за многолетнюю помощь и поддержку при выполнении работы; научному консультанту, заведующей отделом ВКМ Института биохимии и физиологии микроорганизмов им. Г.К. Скрябина РАН, д.б.н. Л.Н. Евтушенко - за консультативную помощь при выполнении таксономических исследований. Автор признателен коллегам лаборатории, участвовавшим в проведении исследований, чей вклад отражен в совместных публикациях - к.б.н. О.В. Ястребовой, к.б.н. Д.О. Егоровой, к.б.н. Л.Н. Ананьиной, к.б.н. Е.С. Шумковой, к.б.н. А.В. Назарову, а также другим сотрудникам ИЭГМ УрО РАН, способствовавшим успешному выполнению работы. Особую благодарность автор выражает Т.В. Цой и О.В. Мальцевой; сотрудникам ИБФМ РАН к.б.н. И.А. Кошелевой, д.б.н. Л.А. Головлевой, д.б.н. И.П. Соляниковой и другим сотрудникам института за искреннюю поддержку и всестороннюю помощь в проведении исследований.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования
Микроорганизмы и плазмиды. Исследуемые бактериальные штаммы были изолированы из образцов почв, грунтов и донных отложений, отобранных с трех локальных территорий, загрязненных отходами предприятий химической промышленности, территории завода конденсаторов г. Серпухова Московской области; территории предприятия ОАО «Галоген» г. Перми; производственных площадок соледобывающего производства ОАО «Уралкалий» (г. Березники, Пермский край). При изучении генетических систем раннего дегалогенирования ХБК использовали штаммы Arthrobacter globiformis КЗТ1, P. putida КЗ6 (Зайцев, Карасевич, 1981, 1984, 1985), P. aeruginosa 142 (Романов и др., 1993), а также P. putida mt-2 (Bagdasarian et al., 1981) и Pseudomonas sp. B13 (Dorn et al., 1974). Для проведения генетических исследований (определение размера D плазмид, конъюгационные эксперименты) штаммы P. putida BS394 (cys-, nah-, sal-), P. putida PpG7 (Dunn, Gunsalus, 1973) и pRK2013, NPL-1, NAH7, pASN1, pDK8 были получены из коллекции лаборатории биологии плазмид ИБФМ РАН. При клонировании и изучении функциональных генов использовали рекомбинантные штаммы E. coli: JM109 (Yanish-Perron et al., 1985), DH5
 F' («Bethesda Research Laboratories», США),
F' («Bethesda Research Laboratories», США),  925 (F+ minA minB thr leu thi) (Stoker et al., 1984) и векторные плазмиды pUC19 (Yanish-Perron et al., 1985), BlueScript («Stratagene», США), pSP329 (Schmidhauser, Helinski, 1985), pRK415(Keen et al., 1988). Для сравнительного изучения использовали типовые культуры из фонда Всероссийской коллекции микроорганизмов (ИБФМ РАН, г. Пущино), Региональной профилированной коллекции алканотрофных микроорганизмов (ИЭГМ УрО РАН, г. Пермь) и из зарубежных коллекций.
925 (F+ minA minB thr leu thi) (Stoker et al., 1984) и векторные плазмиды pUC19 (Yanish-Perron et al., 1985), BlueScript («Stratagene», США), pSP329 (Schmidhauser, Helinski, 1985), pRK415(Keen et al., 1988). Для сравнительного изучения использовали типовые культуры из фонда Всероссийской коллекции микроорганизмов (ИБФМ РАН, г. Пущино), Региональной профилированной коллекции алканотрофных микроорганизмов (ИЭГМ УрО РАН, г. Пермь) и из зарубежных коллекций. Методы. Для выделения и роста бактерий-деструкторов были использованы жидкие и агаризованные минеральная среда К1 (Зайцев, Карасевич, 1981) и минеральная среда Раймонда (Розанова, Назина, 1982). В качестве полноценных сред использовали среду LB (Маниатис и др., 1984), а также модифицированную среду Раймонда при добавлении 5 г/л триптона и 2.5 г/л дрожжевого экстракта в качестве ростовых субстратов. Чистые культуры бактерий хранили в глицерине при -80°С и в лиофильно высушенном состоянии. Для получения биомассы с целью измерения активностей и очистки ферментов проводили культивирование бактерий в среде К1 методом периодического культивирования, используя нафталин, фенантрен, салицилат, 4ХБК в качестве единственного источника углерода и энергии. Определение количества биомассы проводили спектрофотометрически при λ=540 нм. Количество колониеобразующих единиц (КОЕ) определяли методом серийных разведений с последующим высевом и подсчетом колоний бактерий на чашках с полноценной агаризованной средой. Кинетические характеристики растущей периодической культуры определяли согласно рекомендациям (Перт, 1978).
Морфологические и физиолого-биохимические признаки бактерий изучали по стандартным методикам (Методы общей бактериологии, 1983). Состав аминокислот и сахаров в препаратах клеточных стенок (Schleifer, Kandler, 1972) определяли хроматографически (Suzuki et al., 1993). Определение состава жирных кислот целых клеток и хинонов определяли с использованием методов ГЖХ и масс-спектрометрии (Zhilina et al., 1997; Suzuki et al., 1993).
Выделение тотальной ДНК проводили методом, описанным (Current protocols in molecular biology, 1995). Нуклеотидный состав ДНК определяли по температуре ее тепловой денатурации (Owen, Lapage, 1976). Выявление клонов и предварительную группировку штаммов бактерий осуществляли методами ДНК типирования (REP-ПЦР, BOX-ПЦР и ARDRA) как описано ранее (Versalovic et al. 1994; Scortichini et al., 2002). Амплификацию 16S рРНК генов проводили по описанным методам с использованием универсальных бактериальных праймеров (Weisburg et al., 1991). Определение нуклеотидных последовательностей гена 16S рРНК и функциональных генов проводили с применением набора реактивов CEQ Dye Terminator Cycle Sequencing Kit на автоматическом секвенаторе MegaBASE 1000 в соответствии с рекомендациями производителя («JSC GE Healthcare», США). Для филогенетического анализа использовали последовательности 16S рРНК генов, депонированные в GenBank (http:/www.ncbi.nlm.nih.gov) и EzTaxon (on.org). Последовательности 16S рРНК генов выравнивали с помощью программы Clustal W (Thompson et al., 1994). Построение филогенетических древ производили с помощью пакета программ Treecon (Van de Peer, DeWachter, 1994).
Плазмидную ДНК выделяли стандартными (Marco et. al., 1982; Бабыкин и др., 1984) и оригинальными методами. Электрофорез в агарозном геле, обработку ДНК эндонуклеазами рестрикции, щелочной фосфатазой, нуклеазой Bal31, фрагментом Кленова ДНК-полимеразы I E.coli, лигазой фага Т4 проводили в соответствии с рекомендациями (Маниатис и др., 1984). Коньюгационный перенос плазмидной ДНК осуществляли, используя модифицированный метод (Dunn, Gunsalus, 1973). Элиминация бактериальных плазмид проводилась согласно стандартной методике (Rheinwald et al., 1973). Трансформацию плазмидных ДНК в клетки E. coli и P. putida осуществляли по общепринятой методике (Dagert, Ehrlich, 1979), а также методом электропорации на приборе E. coli Gene Pulser («Bio-Rad Laboratories», США). Клонотеки генов A. globiformis КЗТ1 и P. aeruginosa 142 конструировали при использовании плазмидных векторов pUC19, BlueScript, pSP329 в клетках E. сoli JM109 и E. сoli DH5
F'. Для обнаружения рекомбинантных клонов, содержащих fcb- и ohb-гены, использовали оригинальные подходы с применением методик (Резников и др., 1970; Parke, 1992). Реакцию ник-трансляции, перенос ДНК на нитроцеллюлозные фильтры, гибридизацию ДНК-ДНК осуществляли по (Rigby et al., 1977; Southern, 1975). Получение мини-клеток E.coli и включение 35S-метионина проводили в соответствии с рекомендациями (Stoker et al.,1984). Электрофорез белков проводили по (Laemmli, 1970), используя 12.5% полиакриламидный гель. Структуру ассоциации бактерий SMB3 исследовали методом ДГГЭ фрагментов гена 16S рРНК, амплифицированных с применением эубактериальных праймеров 27F и 518R (Tiirola et al., 2002). Электрофорез был выполнен на DcodeTM Universal Mutation System («Bio-Rad Laboratories», США) согласно протоколу (Muyzer et al., 1993)
Для амплификации fcb-генов использовали праймеры, сконструированные на основе гомологичных последовательностей fcb-генов штамма A. globiformis КЗТ1 (Rodrigues et al., 2001). При изучении bph-генов штаммов деструкторов бифенила/ПХБ применяли вырожденные праймеры, подобранные к участкам генов, кодирующих α-субъединицы известных ферментов подсемейства бифенил/толуол диоксигеназ (Witzig et al., 2006).
Продукты бактериального разложения ПАУ и 4ХБК определяли методами ТСХ и ВЭЖХ. Анализ метаболитов деструкции ПХБ проводили спектрофотометрически при длине волны 390 450 нм и методом ВЭЖХ (Maltseva et al., 1999). Динамику дегалогенирования ПХБ и ХБК контролировали по содержанию ионов хлора в культуральной жидкости, освобожденной от клеток (Резников и др., 1970).
Определение активности ферментов проводили спектрофотометрически. Удельные активности нафталин и фенантрен диоксигеназ определяли согласно (Dua, Meera, 1981), салицилат и 1 гидрокси-2-нафтоат гидроксилаз - (Кiyohara, Nagao, 1978), катехол 2,3- и 1,2-диоксигеназ - (Feist, Hegeman, 1969 и Ornston, 1966), гентизат диоксигеназы - (Crawford et al., 1975). Единицу активности определяли как количество фермента, катализирующее превращение 1 мкМ субстрата или образование 1 мкМ продукта в минуту. Относительную активность рассчитывали, принимая за 100% активность с незамещенным субстратом. Концентрацию белка определяли по модифицированному методу Брэдфорда (Schlomann et al., 1990).
Выделение осмопротекторов проводили из галофильных и галотолерантных бактерий, выращенных в минеральной среде Раймонда с разной соленостью и глюкозой в качестве субстрата. Спектры ЯМР регистрировали на импульсном Фурье-спектрометре ЯМР типа WP80SY («Bruker», Германия) (Хмеленина и др., 2000).
Повторность экспериментов, как минимум, трехкратная. При статистической обработке определяли среднюю арифметическую, стандартное отклонение, доверительный интервал, достоверность результата. Для обработки результатов использовали статистический модуль Exсel 5.0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Бактерии-деструкторы полициклических ароматических углеводородов
Методом накопительного культивирования из техногеннозагрязненных почв, грунтов и донных отложений было выделено 240 штаммов-деструкторов нафталина и фенантрена, из них детально охарактеризовано 48 штаммов. Данные анализа нуклеотидных последовательностей генов 16S рРНК, морфологических, физиолого-биохимических и хемотаксономических особенностей изолятов позволили установить их принадлежность родам Arthrobacter, Brevibacterium, Dietzia, Kocuria, Rhodococcus, Streptomyces, Janibacter, Bacillus, Paenibacillus и Pseudomonas. Ниже приводятся характеристики 3-х групп бактерий-деструкторов ПАУ: актинобактерий порядка Actinomyсetales (роды Arthrobacter и Rhodococcus); спорообразующих бактерии родов Bacillus и Pаenibacillus; протеобактерий рода Pseudomonas. Ряд этих бактерий входит в состав нафталинметаболизирующих комплексов (см. раздел 4.1.).
- Бактерии-деструкторы рода Arthrobacter
Результаты генотипирования 9 штаммов рода Arthrobacter, выделенных из почв района солеразработок, показали, что штаммы B901, B904, B905 составляют одну геномогруппу. Штаммы SF27, DF14, SN17, SMB145 и SMB11 отличаются как от данной группы, так и между собой по BOX-ДНК профилям. Пять штаммов (B905, SMB11, SMB145, SF27 и DF14) были наиболее близки по нуклеотидным последовательностям 16S рРНК генов с типовым штаммом вида A. сrystallopoietes (уровень сходства 99.7%), а SN17 - с типовым штаммом вида A. arilaitensis (уровень сходства 99.8%). На рисунке 1 представлена дендрограмма, отображающая положение исследуемых штаммов в системе рода Arthrobacter.
Большинство исследуемых артробактерий являются галоалкалотолерантными организмами: растут при щелочных значениях рН (до 9.0), а также в присутствии до 110 г/л NaCl на полноценных средах и до 60 г/л соли на минеральной среде с нафталином в качестве единственного источника углерода и энергии. Штаммы SMB145 и В905 растут на нафталине в присутствии до 90 г/л NaCl, а штамм SF27 - на фенантрене при содержании до 60 г/л соли.

Рис. 1. Филогенетическое древо, построенное с использованием метода “neighbor-joining”, отображающее положение исследуемых штаммов в системе рода Arthrobacter. Масштаб соответствует 2 нуклеотидным заменам на каждые 100 нуклеотидов. Цифрами показана статистическая достоверность порядка ветвления, определенная с помощью “bootstrap” анализа 1000 альтернативных деревьев (приведены значения выше 50%).
У бактерий выявлены разнообразные пути метаболизма ПАУ. Штаммы B901, B904, B905, SMB11 и SMB145 осуществляют катаболизм нафталина с образованием салицилата и катехола как ключевых интермедиатов. Анализ деструктивных особенностей другой группы штаммов (штаммов SF27, DF14 и SN17) указывает на наличие у них двух путей разложения салицилата - через катехол и гентизат (Yen, Serdar, 1988; Grund et al., 1992). Штаммы SF27 и DF14 эффективно растут на фенантрене и возможных продуктах его метаболизма – 1 гидрокси-2-нафтоате и орто-фталате. Три других штамма B901, B904 и SMB11 способны к слабому росту на фенантрене и не растут на орто-фталате. На основании анализа метаболитов и изучения ферментативных активностей можно предположить, что утилизация фенантрена штаммами SF27 и DF14 идет по орто фталатному пути, как описано для нескольких деструкторов рода Arthrobacter (Бабошин и др., 2005; Seo et al., 2006), а штаммы SMB11, B901 и B904 осуществляют деструкцию фенантрена с образованием 1-гидрокси-2-нафтоата, 1,2 дигидрокси-нафталина и салицилата при участии ферментов пути утилизации нафталина (Iwabuchi, Harayama, 1998). Штаммы SN17, В905, B901 и B904 обладают широкой субстратной специфичностью и способны к деструкции не только нафталина, фенантрена, но и бифенила, фенола, алифатических углеводородов.
Результаты исследований на наличие экстрахромосомальной ДНК показали, что штамм SN17 содержит плазмиду размером ~85 т.п.н., в штаммах DF14 и SF27 зарегистрированы плазмиды размером ~120 т.п.н., в штаммах B901, B904 и B905 – размером ~100 т.п.н. При обработке клеток штамма SF27 митомицином С были получены элиминанты, которые не росли на нафталине, фенантрене и 1 гидрокси 2-нафтоате, но сохраняли способность к росту на салицилате. Полученные данные позволяют предположить, что ферменты первичной атаки ароматического кольца фенантрена и 1-гидрокси-2-нафтоата, а также ферменты утилизации нафталина до салицилата у штамма SF27, кодируются плазмидными генами. Факт локализации генов, ответственных за разложение ароматических соединений, в плазмидах описан для многих бактерий (Hayatsu et al., 1999; Eaton, 2001; Sandu et al., 2005).


Рис. 2. Электрофореграммы плазмидных ДНК: (А) 1 – рВ905; 2 – NAH7 (83 т.п.н.); 3 – –pB904; 4 – pB901. (Б) 1 – рSF27; 2 – рDF14; 3 – рSN17; 4 – NAH7 (83 т.п.н.).
1 2 3 4 1 2 3 4
А В
1.2. Бактерии-деструкторы рода Rhodococcus
Исследовано двенадцать бактерий-деструкторов рода Rhodococcus, одиннадцать из которых выделены из района солеразработок. По результатам типирования выявлено пять геномогрупп, и у десяти штаммов определены нуклеотидные последовательности 16S рРНК генов. Анализ фрагментов генов 16S рРНК размером около 560 п.н. этих бактерий с аналогичными последовательностями типовых штаммов рода Rhodococcus показал 100% сходство со штаммами R. wratislaviensis 13082 T (штаммы G10, B13, В14), R. ruber DSM 43338T (штаммы DB11, SN31), R. erythropolis ATCC 4277T (штамм I25). Штаммы SMB37, B2-1 и B1-4, по всей вероятности, относятся к новому виду, наиболее близкому к R. pyridinivorans (уровень сходства 97.3%), а штамм SMB38 скорее всего представляет новый вид, близкий к R. marinonascens (уровень сходства 98.9%).
Все родококки, выделенные из накопительных культур при повышенной солености среды (30-60 г/л NaCl), способны расти при содержании хлорида натрия до 90-120 г/л в полноценной среде и до 60-90 г/л - в минеральной среде с нафталином. Штаммы SMB37, SMB38, B2-1, B13, B14 и B1-4 растут в пределах рН 6.0-9.0, штаммы G10, I17, I17а - при рН 7.0-9.0, штаммы I25, I31 - при рН 7.0-8.0 и штамм SN31 – при рН 6.0-7.0.
Бактерии росли на нафталине, бифениле, салицилате, гентизате и протокатехате в качестве источника углерода, большинство из них способны расти на бензоле, толуоле, феноле, бензоате и дизельном топливе. Штаммы SMB37, B2 1, B13, B14 и B1 4 осуществляли разложение хлорароматических соединений (ХБК и 2,4-Д). Рост штаммов на салицилате и гентизате дает основание предполагать, что разложение нафталина изученными штаммами осуществляется по гентизиновому пути, описанному для ряда представителей этого рода (Grund et al., 1992; Di Gennaro et al., 2001; Kulakov et al., 2005).
У двух изученных штаммов были обнаружены плазмиды разного размера (рис. 4): у SN31 – одна плазмида (~100 т.п.н), а у G10 - две плазмиды (~85 т.п.н. и 70 т.п.н.). В то же время, необнаружение плазмид у других изученных штаммов родококков не дает основания считать, что экстрахромосомальная ДНК у них отсутствует, так у представителей этого рода плазмиды часто имеют значительно большую молекулярную массу и для их выявления необходимо использование специальных методов выделения и детекции (Shimizu et al., 2001).
1.3. Спорообразующие бактерии родов Bacillus и Paenibacillus
Филогенетический анализ нуклеотидных последовательностей 16S рРНК генов семи спорообразующих штаммов из района солеразработок показал, что штаммы I12b, I23 и I27 являются представителями разных 16S рРНК-групп рода Bacillus (Ash et al., 1991) (рис. 3). Штаммы I23 и I27 входят в кластер «B. pumilus» (уровни сходства 100% и 99.3%, соответственно, с B. pumilus DSM27T). Штамм I12b наиболее близок к B. hwajinpoensis SW-72T (уровень сходства 98.8%). Полученные данные не позволяют однозначно отнести ни один из вышеперечисленных штаммов к какому-либо известному виду; два из них (I12b и I27), по всей вероятности, представляют собой новые виды. Анализ генов 16S рРНК штаммов I10b и SN501 выявил принадлежность штаммов к роду Paenibacillus (рис. 3), виды которого ранее входили в одну из филогенетических групп Bacillus (Ash et al., 1993). Штамм I10b наиболее близок к P. amylolyticus NBRC 15957T (уровень сходства 16S рРНК генов 99.8%), а штамм SN501 - c P. naphthalenovorans PR-N1T (уровень сходства 99%).
Бактерии являются галоалкалотолерантными организмами, способны к росту в щелочных условиях (до pH 9.0) и при содержании 90-110 г/л NaCl в среде культивирования. Утилизируют дизельное топливо, ряд индивидуальных ароматических и алифатических углеводородов. Paenibacillus sp. SN501 обладает широкой субстратной специфичностью: растет на нафталине, фенантрене, салицилате, орто-фталате, а также феноле и октане. Так как штаммы-деструкторы SN501 и I10b не растут на гентизате и 2 метилсалицилате, и не образуют полуальдегид оксимуконовой кислоты (продукт мета-расщепления катехола), можно предположить, что деструкция салицилата данными штаммами осуществляется с образованием катехола и расщеплением последнего по орто-пути (Mutzel et al., 1996; Alquati et al., 2005). Paenibacillus sp. SN501 растет в минеральной среде с нафталином как единственном источнике углерода и энергии при концентрации соли в среде культивирования до 50 г/л.

Рис. 3. Филогенетическое древо, построенное с использованием метода “neighbor-joining”, отображающее положение исследуемых штаммов в системе родов Bacillus и Paenibacillus. Цифрами показана статистическая достоверность порядка ветвления, определенная с помощью “bootstrap анализа” 1000 альтернативных деревьев (приведены значения выше 80%).
1.4. Бактерии-деструкторы рода Pseudomonas
Грамотрицательные бактерии-деструкторы (штаммы SN11, SN21, SN101, DN13, G51) были выделены при культивировании на минеральной среде К1 с нафталином в качестве источника углерода, без добавления NaCl. У штаммов G51, DN13 и SN11 определены нуклеотидные последовательности фрагментов гена 16S рРНК размером около 600 п.н., анализ которых подтвердил принадлежность штаммов к роду Pseudomonas. Наибольшее сходство сравниваемых нуклеотидных последовательностей с таковыми типовых штаммов рода Pseudomonas: для штамма G51 - с P. putida (98%), для штамма DN13 - с P. plecoglossicida (98%) и для штамма SN11 - с P. oryzihabitans (99%).
Все штаммы росли на нафталине и салицилате - промежуточном продукте разложения нафталина, и были способны к росту на 5метилсалицилате - что предполагает наличие у них ферментов орто- и мета-пути расщепления катехола (Timmis, Lehrbach, 1985; Yen, Serdar, 1998). Следует также отметить, что после продолжительного культивирования на фенантрене, изученные псевдомонады приобретали способность к росту на этом субстрате, а также на 1 гидрокси 2 нафтоате (промежуточном продукте биодеградации фенантрена). Возможность модификации генетических систем биодеградации нафталина при длительном инкубировании на фенантрене была показана для штамма P. putida BS3703, который приобретал способность к конститутивному синтезу ферментов "верхнего пути" окисления ПАУ и к росту на фенантрене и 1-гидрокси-2нафтоате (Балашова и др., 1997).
У Pseudomonas sp. SN11 были определены активности ферментов деградации нафталина и фенантрена. В связи с тем, что ферменты катаболизма нафталина у большинства бактерий индуцируются салицилатом (Barnsley, 1975), измерение активности ключевых ферментов биодеградации проводили как в отсутствие, так и в присутствии салицилата. Полученные данные подтверждают факт утилизации нафталина по пути диоксигеназного расщепления ароматического кольца с образованием салицилата и последующим его расщеплением через катехол. Индукция салицилатом фенантрен диоксигеназной активности и 1 гидрокси-2-нафтоат гидроксилазы свидетельствует о возможности утилизации фенантрена штаммом Pseudomonas sp. SN11 с участием ферментов биодеградации нафталина (Балашова и др., 1997). Низкая активность гентизат 1,2-диоксигеназы при индукции салицилатом свидетельствует о возможности окисления данным штаммом салицилата до гентизиновой кислоты, как описано на примере штамма Pseudomonas sp. U2 (Fuenmayor et al., 1998). Установлено, что салицилат является индуктором ферментов пути деградации нафталина: нафталин диоксигеназы, салицилат гидроксилазы и катехол 1,2-диоксигеназы, а также индуктором фенантрен диоксигеназной активности и 1-гидрокси-2-нафтоат гидроксилазы, необходимых для утилизации фенантрена с участием ферментов деградации нафталина. Фермент мета-пути расщепления катехола – катехол 2,3 диоксигеназа имеет высокую конститутивную активность и не индуцируется салицилатом.
Все исследованные штаммы рода Pseudomonas содержат плазмиды размером ~85 т.п.н. (рис. 4). Для доказательства плазмидной локализации генов, участвующих в разложении ПАУ, проведены эксперименты по элиминации плазмид из штамма SN11 и конъюгационному переносу. Так, перенос плазмиды рSN11 в реципиентный штамм P. putida BS394 (cys- nah- sal-) приводил к образованию трансконъюгантов с фенотипом Cys Nah+ Sal+ Phn+. Полученные результаты свидетельствуют, что плазмида рSN11 является конъюгативной и содержит гены разложения нафталина и салицилата; деградация фенантрена штаммом SN11 также контролируется плазмидой. Аналогичные эксперименты по конъюгационному переносу плазмид были проведены с использованием штаммов SN101 и G51. Как и в случае плазмиды pSN11, плазмиды из этих штаммов являются конъюгативными и содержат гены деградации нафталина. Рестрикционный анализ с использованием эндонуклеазы рестрикции EcoRI показал, что плазмиды из штаммов SN11 и G51 имеют одинаковый набор рестрикционных фрагментов, что позволяет предположить идентичность данных плазмид.
1 2 3 4 5 6 7
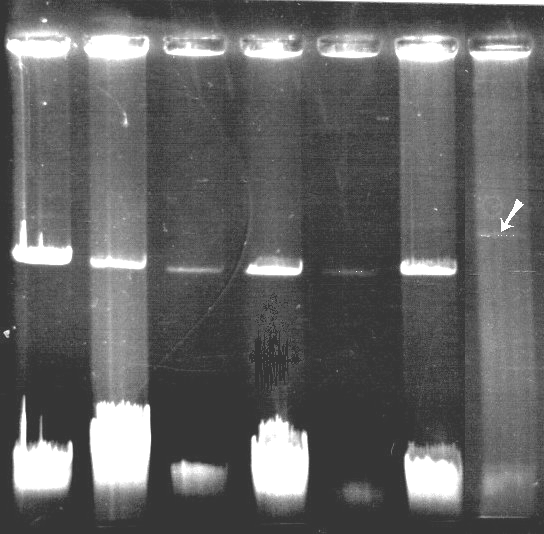
Рис. 4. Электрофореграмма плазмидных ДНК штаммов-деструкторов ПАУ: (А) 1 – NAH7 (83 т.п.н.); 2 – рDN13; 3 – рSN11; 4 – рSN21; 5 – рSN101; 6 – рG51; 7 – рSN31.
1.5. Галотолерантные свойства бактерий-деструкторов ПАУ
Бактерии-деструкторы ПАУ, выделенные из района солеразработок, способны как к росту на средах, не содержащих хлорид натрия, так и на средах с повышенным содержанием соли. Большинство штаммов росли на полноценных средах при концентрации NaCl выше 90 г/л. Кроме того, все исследованные бактерии, кроме Paenibacillus sp. SN501 (растет в присутствии 50 г/л NaCl), способны к росту на агаризованной среде с нафталином в качестве единственного источника углерода и энергии, при 60 г/л NaCl. Ряд штаммов росли в присутствии более высоких концентраций соли (см. раздел 4.1.). Полученные результаты позволяют отнести бактерии к умеренно галотолерантным микроорганизмам (Кашнер, 1981).
Следующим этапом изучения являлось установление некоторых механизмов галотолерантности у исследуемых бактерий. Полученные нами трансконъюгантные штаммы (см. раздел 1.4.), содержащие плазмиду из галотолерантного штамма Pseudomonas sp. SN11, были проверены на способность к росту на средах с повышенным содержанием соли (табл. 2).
Таблица 2. Рост рекомбинантных бактерий при повышенном содержании NaCl
Штаммы | Полноценная среда Раймонда, содержание NaCl (г/л) | Среда Раймонда с нафталином, содержание NaCl (г/л) | ||||
| 40 | 60 | 90 | 25 | 40 | 60 | |
| Pseudomonas sp. SN11 | ++ | ++ | ++ | ++ | ++ | + |
| P.putida BS394 | + | | | | | |
| P.putida BS394-(SN11-81) | ++ | + | | ++ | + | |
| P.putida BS394-(SN11-82) | ++ | + | | ++ | + | |