
Отчет о научно-исследовательской работе разработать научные основы селекции и семеноводства риса. Создать новые сорта, ресурсосберегающие
| Вид материала | Отчет |
- Удк 635. 649:(631. 52+631. 53) Теоретическое обоснование и практическое использование, 946.29kb.
- Удк (631. 527+631. 531): 635. 34 Научное обоснование и разработка системы методов селекции, 1105.12kb.
- Проект Стратегия развития селекции и семеноводства сельскохозяйственных культур в Российской, 552.1kb.
- Особенности селекции, семеноводства и технологии возделывания сортов нута, адаптированных, 1429.88kb.
- Реферат отчет о научно-исследовательской работе состоит, 61.67kb.
- Отчет о научно-исследовательской работе по теме, 60.42kb.
- Отчёт о научно-исследовательской работе за 2009 год, 851.3kb.
- Отчёт о научно-исследовательской работе за 2011 год, 1208.93kb.
- Отчёт онаучно-исследовательской работе гу нии но ур за 2010 год, 997.69kb.
- Отчет о научно-исследовательской работе профессорско-преподавательского состава, 617.56kb.
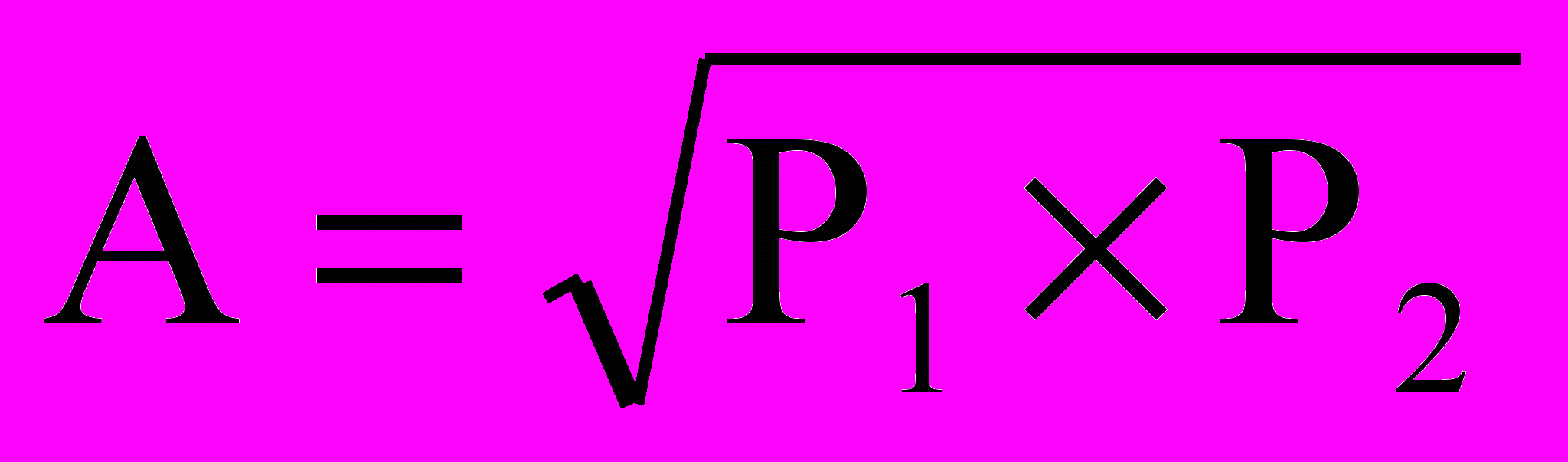 достаточно полно отражает его фактическую величину. Однако при определении теоретического значения величины прогноза конкретного признака не учитывается эффект межаллельной комплементации и взаимодействия генов, а также проявления моногетерозиса и аддитивного эффекта.
достаточно полно отражает его фактическую величину. Однако при определении теоретического значения величины прогноза конкретного признака не учитывается эффект межаллельной комплементации и взаимодействия генов, а также проявления моногетерозиса и аддитивного эффекта.В связи с этим возникает необходимость более точно оценивать величину проявления признака у гибрида в сравнении с родительскими формами. Таким показателем является коэффициент или степень доминирования того или иного признака в гибридах первого и последующих поколений.
Степень доминирования hp у гибридов F1 определяется по формуле А. Густафсона и И. Дормлинга:
hp = F1 – Pmin / Pmax – Pmin, (2)
При скрещивании растений, различающихся между собой по продуктивности, высоте стебля, длине колоса и т. д., у гибридов F1 обычно проявляется промежуточный характер наследования, а в F2 наблюдается плавный переход между крайними вариантами, когда очень трудно разграничить фенотипические классы с различной выраженностью соответствующего признака.
В основе этого явления лежит полимерный характер наследования признаков, определяемых большим числом пар генов. Трансгрессии и новообразования у гибридов возникают наиболее часто при использовании этого метода потому, что многие хозяйственно-полезные признаки у растений обусловлены несколькими полимерными генами. В результате генетической рекомбинации при скрещивании в отдельных генотипах происходит трансгрессивное сочетание в одном генотипе полимерных генов аддитивного действия, что обусловливает более сильное выражение признака по сравнению с обеими родительскими формами.
В нашей работе мы рассчитывали частоту и степень трансгрессии.
Данные проведенного расчета степени доминирования, частоты и степени трансгрессии по изучаемым признакам представлены в таблицах 30 и 31.
Данные таблицы 30 показывают, что по признаку высота растения согласно расчетам у гибрида имеем сверхдоминирование. Длина колоса наследуется как неполное доминирование родителя с меньшей выраженностью признака – Юноны.
По общему количеству колосков в колосе, числу зерен и его во втором поколении массе отмечена депрессия генов. Масса 1000 зерен наследуется по типу неполного доминирования родителя с большей выраженностью признака - Коллеги.
По всем изучаемым признакам, кроме высоты растений отмечены невысокие значения степени трансгрессии (2,5-12,0%). В то же время по высоте растений частота возникновения трансгрессивных форм была довольно высокой (82,9%). На втором месте по этому показателю оказались длина колоса и масса 1000 зерен (27,3 и 28,3% соответственно). Полученные данные подтвердили наше предположение о том, что на наследование изучаемых количественных признаков оказывают влияние не менее двух генов. Так, число колосков в колосе и масса 1000 зерен контролируется тремя генами, а остальные признаки – двумя генами.
Таблица 30 – Степень доминирования в гибридной комбинации Коллега Х Юнона F2 по изучаемым признакам
| Признаки | hp | Степень доминирования | Трансгрессия, % | Число генов (i) | |
| степень | частота | ||||
| Высота растения, см | 8,1 | сверхдоминирование | 41,1 | 82,9 | 2 |
| Длина колоса, см | -0,6 | неполное доминирование Юноны | 12,0 | 27,3 | 2 |
| Число колосков в колосе, шт. | -1,2 | депрессия | 6,9 | 5,7 | 3 |
| Число зерен в колосе, шт. | -3,5 | депрессия | 2,5 | 2,7 | 2 |
| Масса зерна с колоса, г | -2,4 | депрессия | 5,7 | 5,7 | 2 |
| Масса 1000 зерен, г | 0,6 | неполное доминирование Коллеги | 3,3 | 28,3 | 3 |
Полученные данные свидетельствуют о том, что у гибрида обнаружена различная степень доминирования по исследуемым количественным признакам.
По признаку высота растения согласно расчетам у гибрида Батько х Аруана имеем сверхдоминирование родителя с большей выраженностью признака, то есть отцовской формы Аруана. По длине колоса и массе зерна с колоса отмечена - депрессия.
Характер наследования общего количества колосков колосе - неполное доминирование родителя с большей выраженностью признака – материнской формы Батько. Число зерен в колосе наследуется по типу полного доминирования Аруаны, а масса 1000 зерен – по неполному доминированию Батько.
Таблица 31 - Характер доминирования в гибридной комбинации Батько Х Аруана F2 по изучаемым признакам
| Признак | hp | Характер доминирования | Трансгрессия, % | Число генов | |
| степень | частота | ||||
| Высота растения | 1,65 | сверхдоминирование Аруаны | 11,2 | 55,7 | 2 |
| Длина колоса | -1,38 | депрессия | 26,1 | 11,1 | 2 |
| Количество колосков в колосе | 0,86 | неполное доминирование Батько | 9,1 | 60,3 | 2 |
| Число зерен в колосе | -1,00 | полное доминирование Аруаны | 5,5 | 19,1 | 2 |
| Масса зерна с колоса | -1,71 | депрессия | 7,4 | 17,9 | 2 |
| Масса 1000 зерен | -0,96 | неполное доминирование Батько | 17,4 | 7,3 | 2 |
По всем изучаемым признакам отмечены невысокие значения степени трансгрессии. В то же время по высоте растений и числу колосков в колосе частота возникновения трансгрессивных форм была довольно высокой (55,7 и 60,3% соответственно). Полученные данные подтвердили наше предположение о том, что на наследование изучаемых количественных признаков оказывают влияние не менее двух генов.
Проведенный анализ наследования важнейших количественных признаков при скрещиваниях двух сортов озимой пшеницы, принадлежащих к разным разновидностям, позволяет в дальнейшем более точно проводить подбор родительских пар для гибридизации.
Вычисление индексов имеет преимущества перед пользованием абсолютными величинами только в тех случаях, когда такие индексы выявляют известную закономерность, незаметную на абсолютных величинах, или когда они оказываются менее изменчивыми, чем абсолютные величины.
Цель построения и использования индексов в селекции:
- для идентификации и отбора физиолого-генетических систем, повышающих урожай (аттракции, микрораспределений, адаптивности, горизонтального иммунитета, «оплаты» лим-фактора почвенного питания, толерантности к загущению, генетической вариабельности длин фаз онтогенеза).
Эти системы нельзя количественно описать «абсолютными величинами», они не являются признаками и проявляют себя лишь в определенных признаковых координатах, где обнаруживается разнонаправленность их реагирования на экологические и генетические воздействия. Поскольку эти системы быстро и просто без дорогостоящего оборудования и приборов могут быть изучены в виде индексов, они используются селекционерами и довольно активно внедряются в селекционные технологии.
Для элиминации средовых (модификационных) эффектов, отраженных на компонентных признаках индекса, но исчезающих при построении индекса.
Перечислим периодически используемые в генетико-селекционной практике и литературе индексы:
1. «Полтавский индекс» - отношение массы зерна с колоса (г) к длине верхнего междоузлия (см);
2. «Мексиканский индекс» - отношение массы зерен с колоса (г) к высоте растения (см);
3. «Индекс интенсивности» - отношение массы стебля (г) к высоте растения (см);
4. «Индекс продуктивности колоса» - отношение массы зерен с колоса (г) к массе колоса с семенами и мякиной (г);
5. «Индекс линейной плотности колоса» - отношение числа зерен с колоса к длине колоса (см);
6. «Индекс потенциальной продуктивности колоса» - отношение массы зерен с колоса (г) к массе колоса с семенами (г), умноженное на число зерен в колосе;
7. «Канадский индекс (удельный урожай колоса)» - отношение массы зерен с колоса (г) к длине колоса (см);
8. «Индекс микрораспределений» - отношение массы зерен c колоса (г) к массе мякины (половы) с колоса (г).
Существуют и другие, менее распространенные и редко употребляемые индексы, построенные из морфологических мерных и счетных признаков.
Для селекции наиболее важно создание экспрессных методов точной идентификации главных физиолого-генетических систем, повышающих урожай в данной конкретной среде, а не генетическая характеристика количественного признака, которая почти обязательно изменится в другой среде. На пригодность того или иного индекса в селекции в конкретной зоне селекционеры обычно выходят «на ощупь», т.е. методом проб и ошибок.
В связи с этим, мы с своей работе изучали индексы для использования их в отборах ценных форм для селекции. Полученные данные представлены в таблице 32.
Таблица 32 - Селекционно-генетические и физиологические индексы продуктивности растений озимой пшеницы (на примере комбинации Коллега Х Юнона)
| № | Индекс | Коллега | Коллега х Юнона | Юнона |
| 1 | «Полтавский индекс» | 0,096 | 0,051 | 0,075 |
| 2 | «Мексиканский индекс» | 0,033 | 0,018 | 0,028 |
| 3 | «Индекс продуктивности колоса» | 0,80 | 0,89 | 0,76 |
| 4 | «Индекс линейной плотности колоса» | 5,48 | 5,67 | 6,91 |
| 5 | «Индекс потенциальной продуктивности колоса» | 46,81 | 48,67 | 49,10 |
| 6 | «Канадский индекс (удельный урожай колоса)» | 0,25 | 0,18 | 0,22 |
| 7 | «Индекс микрораспределений» | 4,11 | 8,50 | 3,24 |
Указанные индексы можно условно разделить на три группы: 1. характеризующие продуктивность листа, 2. аттрагирующую способность колоса и 3. продуктивность колоса.
Полтавский, канадский и мексиканский индексы демонстрируют аттрагирующую способность растений. Судя по этим показателям, наибольшей аттрагирующей способностью обладают родительские формы, особенно Коллега, у гибрида она невысокая. Однако, что касается индекса микрораспределений, то здесь картина иная. Гибрид имел наивысшую способность усваивать питательные вещества из половы колоса, нежели родительские сорта.
В то же время, индексы, характеризующие продуктивность колоса указывают на способность гибрида формировать хорошо озерненный колос с выполненным зерном. Индексы линейной плотности, потенциальной продуктивности колоса и индекс микрораспределений у гибрида оказались выше, чем у родительских форм. Это дает надежду на отбор ценных форм из популяции гибрида.
Проведенный анализ наследования важнейших количественных признаков при скрещиваниях двух сортов озимой пшеницы, принадлежащих к разным разновидностям, позволяет в дальнейшем более точно проводить подбор родительских пар для гибридизации.
Выводы:
- Растения родительских форм и гибрида сформировали довольно крупные листья. Гибрид по площади флагового и подфлагового листьев имел промежуточные значения. У всех изучаемых растений сортов и гибрида площадь флагового листа была более вариабельной, чем подфлагового. Коэффициент вариации двух верхних листьев у Коллега х Юнона был выше, чем у родительских форм.
- ИЛП гибрида Коллега х Юнона имел средние значения по сравнению с родительскими формами. Это связано с меньшей площадью флагового листа в комбинации, нежели у родительских форм. У индекса продуктивности листьев гибрида отмечена такая же тенденция. Это и повлияло на формирование невысокой массы зерна у гибрида по сравнению с родителями.
- растения гибридной комбинации имели средние значения аттрагирующей способности, которая характеризуется индексом продуктивности листа (0,066). наибольшей аттракцией обладал сорт Коллега (0,078), а наименьшей – Юнона (0,056).
- в результате проведенного скрещивания в первом поколении наблюдалось превышение родительских форм гибридами F1 по всем признакам, кроме высоты растения (у гибрида Коллега Х Юнона) и числа колосков в колосе (у гибрида Батько Х Аруана). Во втором поколении у гибрида наблюдалось снижение значений по всем признакам, кроме высоты растения. В F2 отмечено общее снижение эффекта гетерозиса по всем изучаемым признакам. Гетерозис по отношению к обоим родительским формам Батько и Аруана, а также к их среднему значению был получен по высоте растений, длине колоса и числу колосков в колосе. Во втором поколении наблюдалось широкое варьирование признаков, вызванное расщеплением, что делает возможным проведение отборов необходимых перспективных форм.
- Размах изменчивости высоты растений у полученных гибридов второго поколения оказался выше, чем у родительских форм (55-127 см). Среди изучаемых растений встречались трансгрессивные формы, которые были выше или ниже родительских форм. На формирование признака высоты растений в большей степени повлияла отцовская родительская форма Юнона, нежели материнская.
- в популяции растений гибрида Коллега Х Юнона имелись растения с длиной колоса 13,1-14,7 см (5,7%), что способствовало отбору элитных растений с длинным колосом. Размах варьирования у гибрида был близок к родительской форме Юноне, которая в большей степени оказала влияние на наследование этого признака. На длину колоса гибрида Батько Х Аруана в большей степени оказывает влияние материнская форма, взятая для скрещивания. Однако, были выделены формы с длиной колоса более 12 см, благодаря сорту Аруана, взятому для скрещивания в качестве отцовской форме.
- по числу колосков в колосе положительная трансгрессия наблюдалась по отношению к родительской форме Юнона. Благодаря высокому числу колосков в колосе у Коллеги, в потомстве гибрида были обнаружены растения с числом колосков больше 22 шт. Наибольшее влияние на формирование данного признака у гибрида Батько Х Аруана оказала материнская родительская форма – Батько. В полученной гибридной популяции можно вести отбор особенно ценных форм с плотным колосом.
- Наибольшее влияние на наследование числа зерен в колосе у гибрида Коллега Х Юнона оказала материнская форма Коллега. Коэффициент вариации и дисперсия у гибрида по изучаемому признаку дает возможность на отбор хорошо озерненных колосьев для дальнейшей селекции.
- Полученный гибрид Коллега Х Юнона второго поколения имел значения массы зерна с колоса ниже, чем его родители. Верхние значения лимита у него достигали 3,7 г, что говорит о достаточной крупности зерна у некоторых растений. Коэффициент изменчивости данного гибрида составил 30,3%. Наибольший родительский эффект у гибрида Батько Х Аруана имел Батько. В комбинации встречались растения с массой зерна от 2,6 г (6,1%) до 3,1 г (0,4%). Это дало возможность для отбора наиболее ценных растений для дальнейшего изучения.
- на формирование массы 1000 зерен у гибрида Коллега х Юнона оказала влияние материнская форма Коллега. В популяции были выделены растения с массой 1000 зерен более 59,5 г. Во втором поколении у гибрида Батько Х Аруана средняя масса 1000 зерен полученного гибрида была ниже по сравнению с первым поколением. В F2 отмечен самый высокий размах изменчивости (30,0–64,9 г), и коэффициент вариации (16,6%). Это связано с расщеплением в потомстве. у гибрида Батько х Аруана получены растения с массой 1000 зерен выше 50 г, что позволило выделить интересные с точки зрения селекции формы.
- продуктивность колоса сортов озимой мягкой пшеницы взаимосвязана с массой колоса, массой зерна и числом зерен в нем. высокая взаимосвязь отмечена между длиной колоса и числом зерен в нем, а также массой зерна с колоса. Это дает возможность для отбора лучших перспективных форм по признаку «длина колоса» для дальнейшего изучения.
- по признаку высота растения у гибрида Коллега х Юнона имеем сверхдоминирование. Длина колоса наследуется как неполное доминирование родителя с меньшей выраженностью признака – Юноны. По общему количеству колосков в колосе, числу зерен и его во втором поколении массе отмечена депрессия генов. Масса 1000 зерен наследуется по типу неполного доминирования родителя с большей выраженностью признака - Коллеги.
- у гибрида Батько Х Аруана обнаружена различная степень доминирования по исследуемым количественным признакам. По признаку высота растения имеем сверхдоминирование родителя с большей выраженностью признака, то есть отцовской формы Аруана. По длине колоса и массе зерна с колоса отмечена – депрессия, по общему количеству колосков колосе и массе 1000 зерен – неполное доминирование Батько. Число зерен в колосе наследуется по типу полного доминирования Аруаны.
- По всем изучаемым признакам у гибрида Батько Х Аруана отмечены невысокие значения степени трансгрессии. В то же время по высоте растений и числу колосков в колосе частота возникновения трансгрессивных форм была довольно высокой (55,7 и 60,3% соответственно). Полученные данные подтвердили наше предположение о том, что на наследование изучаемых количественных признаков оказывают влияние не менее двух генов.
- Индексы, характеризующие продуктивность колоса указывают на способность гибрида формировать хорошо озерненный колос с выполненным зерном. Индексы линейной плотности, потенциальной продуктивности колоса и индекс микрораспределений у гибрида оказались выше, чем у родительских форм. Гибрид демонстрировал хорошую способность усваивать питательные вещества из половы колоса, нежели родительские сорта. Это дает надежду на отбор ценных форм из популяции гибрида.
ЗАКЛЮЧЕНИЕ
1. В отчетном году выполнен полный комплекс работ по изучению созданного селекционного материала.