
Отчет о научно-исследовательской работе разработать научные основы селекции и семеноводства риса. Создать новые сорта, ресурсосберегающие
| Вид материала | Отчет |
- Удк 635. 649:(631. 52+631. 53) Теоретическое обоснование и практическое использование, 946.29kb.
- Удк (631. 527+631. 531): 635. 34 Научное обоснование и разработка системы методов селекции, 1105.12kb.
- Проект Стратегия развития селекции и семеноводства сельскохозяйственных культур в Российской, 552.1kb.
- Особенности селекции, семеноводства и технологии возделывания сортов нута, адаптированных, 1429.88kb.
- Реферат отчет о научно-исследовательской работе состоит, 61.67kb.
- Отчет о научно-исследовательской работе по теме, 60.42kb.
- Отчёт о научно-исследовательской работе за 2009 год, 851.3kb.
- Отчёт о научно-исследовательской работе за 2011 год, 1208.93kb.
- Отчёт онаучно-исследовательской работе гу нии но ур за 2010 год, 997.69kb.
- Отчет о научно-исследовательской работе профессорско-преподавательского состава, 617.56kb.
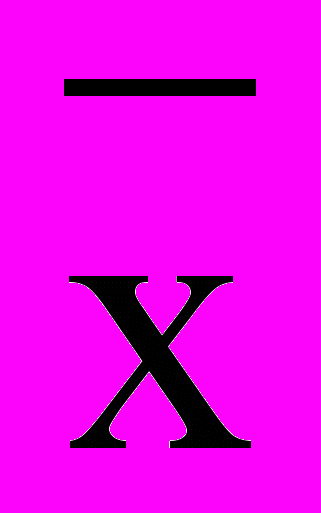 ), лимиты, коэффициенты вариации (V) и корреляции.
), лимиты, коэффициенты вариации (V) и корреляции.Полученные данные подвергались математической обработке на ПК в программе Excel и «Полиген А», разработанной А.Ф. Мережко.
Коллега. Включен в Госреестр по Северо-Кавказскому региону в 2007 году. Рекомендован для возделывания в Центральной и Южно-предгорной зонах Краснодарского края и Горной зоне Ставропольского края.
Разновидность лютесценс. Куст полупрямостоячий - промежуточный. Растение короткое. Восковой налет на колосе, верхнем междоузлии и влагалище флагового листа очень сильный. Колос цилиндрический, средней плотности, белый, средней длины - длинный. Остевидные отростки на конце колоса очень короткие. Опушение верхушечного сегмента оси колоса с выпуклой стороны среднее. Плечо прямое - приподнятое, средней ширины. Зубец прямой - слегка изогнутый, очень короткий - короткий. Нижняя колосковая чешуя на внутренней стороне имеет очень слабое опушение. Зерновка окрашенная. Масса 1000 зерен 37-48 г. Средняя урожайность в регионе - 46,6 ц/га, в Краснодарском крае - на уровне стандарта Палпич, в Ставропольском крае по занятому пару и пропашным культурам прибавка к стандарту Скифянка составила 4,5 ц/га при урожайности 57,0 ц/га. Максимальная урожайность 87,6 ц/га получена в Ставропольском крае в 2005 г. Среднеспелый. Вегетационный период 229-268 дней. Созревает на 3-6 дней позднее стандарта Палпич. По зимостойкости в год проявления признака превышает Скифянку на 0,7-1,0 балла. Высота растений 68-93 см. Устойчивость к полеганию выше, чем у сорта Палпич на 0,6-0,9 балла. Засухоустойчивость на уровне стандарта. В Ставропольском крае максимальные прибавки урожайности обеспечивает при выращивании по занятому пару и пропашным культурам.
Хлебопекарные качества на уровне пшеницы филлера. Умеренно устойчив к бурой ржавчине; умеренно восприимчив к мучнистой росе, септориозу, фузариозу колоса. В полевых условиях стеблевой ржавчиной поражался слабо как и стандарт Скифянка, слабовосприимчив к твердой головне; умеренно восприимчив к желтой ржавчине.
Юнона. Происхождение. Сорт создан методом гибридизации и индивидуального отбора в F3 из комбинации Ейка Лютесценс 5573 h 16.
Общая характеристика. Высота растений 80-90 см. Высокоустойчивый к полеганию. Среднеспелый. Разновидность lutescens.
Урожайность. В конкурсном сортоиспытании на Северокубанской сельскохозяйственной опытной станции в среднем за три года (2002-2004) сорт Юнона при урожайности 72,3 ц с 1 га превысил сорт Скифянку на 9,2 и зимостойкий сорт Победу 50 на 6,0 ц с 1 га.
Мукомольные и хлебопекарные качества. По качеству зерна соответствует сильной пшенице.
Устойчивость к болезням и климатическим условиям. На инфекционном фоне высокоустойчив к пыльной головне, бурой ржавчине, мучнистой росе, устойчив к септориозу. Обладает полевой устойчивостью к желтой ржавчине. Восприимчив к фузариозу колоса и твердой головне. Отличается высокой морозостойкостью (на уровне Краснодарской 39). Засухоустойчив.
Зона возделывания. Рекомендуется для испытания в Северо-Кавказском, Центрально-Черноземном, Средневолжском и Нижневолжском регионах.
Сроки сева. Оптимальные для зоны.
Нормы высева. 5 млн. всхожих зерен на 1 га.
Батько. Происхождение. Получен методом сложной ступенчатой гибридизации и индивидуального отбора из гибридной популяции КН 4217Г25908-4228/КН 5126h85/52.
Общая характеристика. Короткостебельный, на 2-3 см выше сорта Юна, устойчив к полеганию. Среднеустойчив к осыпанию. Скороспелый, созревает одновременно или на 1 день позже стандарта. Разновидность lutescens. Зерновка крупная, яйцевидной формы.
Урожайность. Потенциал продуктивности высокий, 100-110 ц с 1 га. В среднем за 3 года изучения в конкурсном сортоиспытании института превысил стандартный сорт Скифянку на 6,4 ц. В 2005 году в опытах по сортовой агротехнике урожай сорта Батько составил 100,0 ц с 1 га, превысив сорт Безостая 1 на 33,4 ц.
Мукомольные и хлебопекарные качества. По всем показателям качества зерна соответствует требованиям, предъявляемым к «сильным» пшеницам.
Устойчивость к болезням и климатическим условиям. Устойчив к твердой головне, бурой и стеблевой ржавчинам. Средне устойчив к септориозу, мучнистой росе и фузариозу колоса. Средне восприимчив к желтой ржавчине. Засухоустойчивость и морозостойкость на уровне стандартного сорта Юна.
Зона возделывания. Допущен к возделыванию в Северо-Кавказском регионе РФ, в Украине. Сроки сева. Оптимальные для зоны.
Норма высева. 4,0-4,5 млн. всхожих семян на 1 га при посеве в оптимальные сроки. В конце оптимальных сроков сева норму высева следует повышать до 5 млн.
Аруана. Сорт зарубежной селекции. Получен для изучения из коллекции ВИР. Разновидность velutinum. Среднерослый сорт, высота растений около 100 см, устойчив к полеганию. Скороспелый сорт, созревает на 1-3 дня раньше Скифянки. Колос цилиндрический, безостый, имеет опушение. Все растение имеет восковой налет и опушение. Средняя урожайность 70 ц/га.
Высокоустойчив к пыльной головне. В слабой степени поражается желтой и стеблевой ржавчинами, мучнистой росой, бурой ржавчиной и септориозом.
Главная роль в создании органического вещества принадлежит фотосинтезу - первоначальному этапу формирования урожая. Важную функцию в накоплении органической массы играют листья. В них протекает процесс фотосинтеза, то есть усвоение из внешней среды углекислоты за счет энергии солнечного света и преобразования ее в химическую энергию органических веществ. В процессе фотосинтеза создаются различные углеводистые соединения, превращающееся затем в крахмал и сахар. При этом углерод и вода, содержащиеся в растении наряду с азотом и другими элементами, участвуют в образовании аминокислот и белков. Таким образом, сухая масса урожая зеленых растений состоит в основном из органических веществ, первично создаваемых растениями в процессе фотосинтеза.
Интенсивность фотосинтеза зависит от площади листовой пластинки. Площадь же листовой пластинки зависит от генотипа сорта, уровня агротехники (правильно подобранные предшественники, удобрения, сроки и способы посева и др.), обеспеченности влагой и других условий.
Селекция пшеницы ведется, в основном, на увеличение площади двух верхних листьев и повышение фотосинтетического потенциала. При этом установлен преимущественный рост фотосинтетического потенциала верхних листьев, снабжающих наливающееся зерно.
В своих опытах мы определяли площадь флагового и подфлагового листа, а также индекс листовой поверхности у сортов озимой мягкой пшеницы и устанавливали их выравненность через коэффициент вариации (V%).
Данные по площади и вариации верхних листьев представлены в таблице 12.
Таблица 12 - Площадь и вариация двух верхних листьев сортов и гибрида озимой пшеницы, 2010-2011 гг.
| родительские формы, гибрид | Густота стояния растений, шт./м2 | Флаговый лист | Подфлаговый лист | Сумма, см2 | ||
| S, см2 | V, % | S, см2 | V, % | |||
| Коллега | 481 | 33,5 | 16,9 | 29,5 | 14,2 | 63,0 |
| Коллега х Юнона | 491 | 30,5 | 24,0 | 30,2 | 16,4 | 60,7 |
| Юнона | 478 | 30,9 | 18,7 | 27,4 | 16,1 | 58,3 |
Из данных таблицы видно, что родительские формы и гибрид Коллега х Юнона сформировали хорошую густоту стояния растений на 1 м2. Сорта, взятые для скрещивания имели примерно одинаковую густоту стояния, а гибрид сформировал на 10 растений больше на 1 м2.
Условия 2010 года оказались благоприятными для развития листовой пластинки. Растения родительских форм и гибрида сформировали довольно крупные листья. Площадь первого и второго листа у материнской формы Коллеги составила соответственно – 33,5 и 29,5 см2, у отцовской формы Юноны – 30,9 и 27,4 см2 соответственно. Гибрид по площади флагового и подфлагового листьев имел промежуточные значения. У всех изучаемых растений сортов и гибрида площадь флагового листа была более вариабельной, чем подфлагового. Коэффициент вариации двух верхних листьев у гибрида был выше, чем у родительских форм.
В целом, суммарная площадь двух верхних листьев была довольно высокой, как у родительских форм, так и у гибрида.
Индекс листовой поверхности зависит от площади флагового листа и густоты продуктивного стеблестоя. Этот показатель характеризует степень покрытия листьями площади посева. Продуктивность листьев – отношение массы хозяйственно ценных органов к площади листьев. Зерновая нагрузка листа - отношение количества зерен с колоса к площади листа.
Как установлено работами ТСХА, запасы пластического материала и ассимиляционные возможности листьев и побегов у различных сортов пшеницы хорошо оцениваются в фазе полного формирования зерна, когда прирост вегетативной массы заканчивается.
Переброска ассимилянтов в зерно зависит от аттрактации колоса. Самой высокой аттрактацией из изучаемых растений характеризуется материнская форма Коллега. Для налива зерна используются свежие продукты фотосинтеза из листьев, стебля и колоса. Чтобы обеспечить наилучшие условия для фотосинтетической деятельности посева, селекционеры создают сорта с вертикальным расположением листьев. Компенсация продуктивности фотосинтеза у них осуществляется за счет влагалища листьев, соломины и колоса.
У всех растений индекс листовой поверхности был больше единицы, что говорит о благоприятных погодных условиях для роста и развития озимой пшеницы. ИЛП гибрида Коллега х Юнона имел средние значения по сравнению с родительскими формами (таблица 13). Это связано с меньшей площадью флагового листа в комбинации, нежели у родительских форм. индекс продуктивности листьев гибрида имел снова промежуточное значение по сравнению с родительскими формами. Это и повлияло на формирование невысокой массы зерна у гибрида по сравнению с родителями.
Индекс зерновой нагрузки листа показывает, какое количество зерен «кормит» 1 см2 листа, т.е. косвенно характеризует аттрагирующую способность растения. У изучаемого нами гибрида на 1 см2 приходится более 2 зерен, когда у родительских форм 1,74 и 1,79 шт. соответственно. Однако, растения гибридной комбинации имели средние значения аттрагирующей способности, которая характеризуется индексом продуктивности листа (0,066). наибольшей аттракцией обладал сорт Коллега (0,078), а наименьшей – Юнона (0,056).
Таблица 13 - Индексы, характеризующие продуктивность листьев озимой мягкой пшеницы, 2010-2011 гг.
| Индекс | Коллега | Коллега х Юнона | Юнона |
| ИЛП | 1,61 | 1,48 | 1,50 |
| Продуктивность листьев | 0,078 | 0,066 | 0,056 |
| Зерновая нагрузка листа | 1,74 | 2,08 | 1,79 |
Таким образом, при создании высокоурожайных сортов пшеницы в основу их получения должен быть положен общий объем ассимилянтов, создаваемый листьями, стеблями, колосом и эффективность его реализации в зерне.
Практическое использование эффекта гетерозиса - одно из крупнейших достижений генетики и селекции за последние десятилетия.
Успехи в этой области свидетельствуют о том, что растениеводство с каждым годом будет всё больше опираться на использование гетерозисных гибридов более продуктивных, чем обычные сорта.
При подборе родительских пар для скрещивания, селекционера всегда интересует вопрос, как проявится гетерозис в той или иной гибридной комбинации. Величина прогноза гетерозиса любого признака растений определяется по следующей формуле.
А=√P1×P2, (1)
где P1 и P2 - средние значения признака родительских форм.
В нашем опыте мы изучили шесть наиболее важных количественных признаков, которые влияют на формирование продуктивности пшеничного растения. Нами была подсчитана теоретическая величина (прогноз) признака в F1 и фактические величины гетерозиса в F1 (таблица 14).
Общее количество колосков в колосе предположительно должно было бы быть 19,5, а в действительности сформировалось около 22 штук, то есть реализация признака составила + 2 шт.
Таблица 14 - Прогнозирование значений признаков у гибридов F1 и фактическое проявление эффекта гетерозиса в F1
| Признаки | Родительские формы | Величина прогноза для F1 | Фактическое значение в F1 у гибрида Коллега х Юнона | |
| Коллега | Юнона | |||
| Высота растения, см | 81,2 | 77,2 | 79,2 | 76,7 |
| Длина колоса, см | 10,4 | 8,9 | 9,6 | 11,3 |
| Число колосков в колосе, шт. | 19,8 | 19,2 | 19,5 | 21,5 |
| Число зерен в колосе, шт. | 51,3 | 52,5 | 51,9 | 78,0 |
| Масса зерна с колоса, г | 2,2 | 1,9 | 2,1 | 3,3 |
| Масса 1000 зерен, г | 43,1 | 36,6 | 39,7 | 41,8 |
По признаку число зерен в колосе реализация признака превысила прогнозируемые значения на – 26,1 шт.
По массе зерна с колоса и массе 1000 зерен фактическая величина признака была снова выше прогнозируемой, что опять-таки говорит об эффекте гетерозиса гибрида первого поколения.
Различная величина реализации того или иного количественного признака связана как с модификационной изменчивостью (в большинстве случаев), так и возможно с выживаемостью растений, поскольку гибель отдельных растений имела место.
В целом можем сделать вывод, что в результате проведенного скрещивания в первом поколении наблюдалось превышение родительских форм гибридами F1 по всем признакам, кроме высоты растения. Превышение гибридами первого поколения исходных родительских форм вызвано эффектом гетерозиса, который был различной силы по изучаемым нами признакам.
Исходя из значений прогнозируемых признаков у родительских форм, высота растений в первом поколении должна была составить 94,1 см, фактическая же составила 97,1 см, а во втором поколении она составила 100,2 см – в гибридах F1 и F2 разница (реализация признака - + 3 см и + 6,2 см соответственно). По длине колоса разница между прогнозируемой и фактической величинами составила у гибридов + 1 см в первом поколении и –1,9 см во втором поколении.
Тaблица 15 - Прогнозирование значений признаков у гибридов F1 и фактическое проявление эффекта гетерозиса в F1 и F2
| Признаки | Родительские формы | Величина прогноза для F1 | Фактическое значение гибрида | ||
| Батько | Аруана | F1 | F2 | ||
| Высота растения, см | 83,7 | 105,8 | 94,1 | 97,1 | 100,2 |
| Длина колоса, см | 11,3 | 11,6 | 11,4 | 12,4 | 9,5 |
| Число колосков в колосе, шт. | 21,8 | 19,1 | 20,4 | 19,8 | 19,7 |
| Число зерен в колосе, шт. | 48,2 | 37,8 | 42,7 | 52,4 | 35,6 |
| Масса зерна с колоса, г | 2,09 | 2,15 | 2,1 | 2,75 | 1,68 |
| Масса 1000 зерен, г | 43,2 | 58,8 | 50,4 | 51,8 | 41,7 |
Общее количество колосков в колосе предположительно должно было бы быть 20,4, а в действительности сформировалось 19,8 штук, то есть реализация признака составила для гибрида F1 - 1,4 шт., а для гибрида F2 - 1,5 шт.
По признаку число зерен в колосе реализация признака в комбинации F1 превысила прогнозируемые значения на 9,7 шт., а в F2 снизилась на 7,1 шт. и составила 52,4 и 35,6 соответственно.
По массе зерна с колоса и массе 1000 зерен фактическая величина признака была снова выше прогнозируемой у гибрида F1, а у гибрида F2 эти показатели оказались ниже.
Различная величина реализации того или иного количественного признака связана как с модификационной изменчивостью (в большинстве случаев), так и возможно с выживаемостью растений, поскольку гибель отдельных растений имела место.
В целом можем сделать вывод, что в результате проведенного скрещивания в первом поколении наблюдалось превышение родительских форм гибридами F1 по всем признакам, кроме числа колосков в колосе. Превышение гибридами первого поколения исходных родительских форм вызвано эффектом гетерозиса, который был различной силы по изучаемым нами признакам. Во втором же поколении F2 наблюдалось снижение значений по всем признакам гибрида, кроме высоты растения.
Данные таблицы 15 свидетельствуют о том, что у полученного гибрида в первом по отношению к материнской форме гетерозис отмечен по признакам высота растений, длина колоса, число зерен и масса зерна с колоса, а также масса 1000 зерен. По отношению в отцовской родительской форме гетерозис наблюдался по всем изучаемым признакам, кроме высоты растений и массы 1000 зерен. Это объясняется тем, что сорт Аруана довольно высокорослый. Наибольшим эффект гетерозиса был по признакам длина колоса, число зерен в колосе и масса зерна с колоса. У гибрида Батько х Аруана гетерозис по отношению к средним значениям родительских форм наибольший был по числу зерен в колосе и их массе.
В F2 отмечено общее снижение эффекта гетерозиса по всем изучаемым признакам. Гетерозис по отношению к обоим родительским формам Батько и Аруана, а также к их среднему значению был получен по высоте растений, длине колоса и числу колосков в колосе. В F2 наблюдалось широкое варьирование признаков, вызванное расщеплением, что делает возможным проведение отборов необходимых перспективных форм.
Изучение наследования количественных признаков на фоне разнообразных экологических условий представляет собой важнейшую фундаментальную и прикладную проблему. В тех случаях, когда удается расшифровать генетическую составляющую во взаимодействии растений со средой, решение селекционных задач многократно упрощается.
Таблица 16 - Фактические величины гетерозиса у гибрида Батько х Аруана
| Признаки | F1 | F2 | ||||
| величина гетерозиса в % к: | ||||||
| ♀ | ♂ | МР | ♀ | ♂ | МР | |
| Высота растения, см | 114,4 | 80,3 | 94,4 | 101,9 | 109,5 | 105,6 |
| Длина колоса, см | 109,8 | 106,9 | 108,4 | 86,1 | 100,4 | 92,7 |
| Число колосков в колосе, шт. | 90,5 | 103,5 | 96,6 | 106,3 | 99,1 | 102,5 |
| Число зерен в колосе, шт. | 108,7 | 138,6 | 121,8 | 98,4 | 88,2 | 93,1 |
| Масса зерна с колоса, г | 131,6 | 127,8 | 129,7 | 78,2 | 89,3 | 83,3 |
| Масса 1000 зерен, г | 120,0 | 88,1 | 101,6 | 78,8 | 98,5 | 87,6 |
Принципы менделевской генетики достаточно широко используются при описании наследования количественных признаков. Разработанные на их основе методологии различаются по характеру генетических и технических ограничений, затратам труда и времени, точности и правдоподобности получаемых результатов.
Как отмечали М.Е. Лобашев (1967), М.М. Тихомирова (1990), Э.Х. Гинзбург (1984) и др., четкой границы между количественными и качественными признаками не существует. Сами количественные признаки различаются между собой как по характеру варьирования, так и по фенотипическому разнообразию в пределах вида.
Средние величины характеризуют всю выборку в целом. Но основное свойство ее членов – свойство изменяться от особи к особи – остается при этом нераскрытым.
Для суждения о степени изменчивости или вариабельности признаков в биометрии наиболее часто используются следующие показатели:
- лимит, или размах изменчивости;
- среднее квадратическое, или стандартное отклонение;
- коэффициент вариации или изменчивости
- дисперсия.
Результаты анализа гибридных популяций и их родительских форм по высоте растения представлены в таблице 17.
Таблица 17 – Характеристика гибрида и родительских форм по высоте
растений
| Гибрид, родительские формы | Х+Sx, см | Lim, см | V, % | σ2 |
| Р.ф. Коллега | 78,6 +0,80 | 65-90 | 8,0 | 6,3 |
| Коллега х Юнона | 93,3 + 0,78 | 55-127 | 16,7 | 240,8 |
| Р.ф. Юнона | 74,4 +1,25 | 67-81 | 5,4 | 4,0 |
Обе родительские формы имели среднюю высоту растения – 78,6 и 74,4 см соответственно. Пределы изменчивости признака (Lim) у Коллеги составили 65-90 см, при коэффициенте варьирования (V) 8,0 %, у Юноны изменчивость в пределах 67-81 см, коэффициент варьирования 5,4 %. У гибридов средняя высота растений была выше, чем у родительских форм.
Размах изменчивости высоты растений у полученных гибридов второго поколения оказался выше, чем у родительских форм (55-127 см). Cсоответственно и коэффициент вариации был достаточно высоким (16,7%). Среди изучаемых растений встречались трансгрессивные формы, которые были выше или ниже родительских форм. Такой большой размах варьирования признака в гибридной популяции дает возможность проведения отборов наиболее интересных растений с точки зрения селекции.
Обе родительские формы имели среднюю высоту растения – 83,7 и 120,3 см соответственно, при ошибке средней арифметической 1,98 и 2,66. Пределы изменчивости признака (Lim) у Батько составили 62–99,5 см, при коэффициенте варьирования (V) 11,4%, у Аруаны изменчивость в пределах 101-140 см и коэффициент варьирования 10,4%. У гибрида первого поколения средняя высота растений составила 97,1 см, а коэффициент изменчивости составил 11,3%.
Таблица 18 – Характеристика гибридов и родительских форм по высоте
растений
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, см | Lim, см | V,% | Х+Sx, см | Lim, см | V,% | |
| Р.ф. Батько | 83,7+1,98 | 62–99,5 | 11,4 | 91,3+0,71 | 83-96 | 3,9 |
| Батько х Аруана | 97,1+2,18 | 64-121 | 11,3 | 100,2+0,96 | 59–134,5 | 15,6 |
| Р.ф. Аруана | 120,3+2,66 | 101-140 | 10,4 | 98,0+3,14 | 67-121 | 16,0 |
Во втором поколении размах изменчивости высоты растений у полученного гибрида увеличился. Среди изучаемых растений встречались трансгрессивные формы, которые были выше или ниже родительских форм. Такой большой размах варьирования признака в гибридной популяции дает возможность проведения отборов наиболее интересных растений, то есть имеется широкий выбор.
Результаты анализа гибридных популяций и их родительских форм по длине колоса представлены в таблицах 19 и 20.
Таблица 19 – Характеристика гибридов и родительских форм по длине
колоса
| Гибрид, родительские формы | Х+Sx, см | Lim, см | V,% | σ2 |
| Р.ф. Коллега | 10,6 + 0,22 | 8,0-12,5 | 10,3 | 2,8 |
| Коллега х Юнона | 9,6 + 0,09 | 6,0-14,0 | 17,9 | 2,9 |
| Р.ф. Юнона | 9,3 + 0,17 | 7,5-11,0 | 9,2 | 0,9 |
По признаку длина колоса средние показатели у гибридных растений второго поколения занимали промежуточное положение по отношению к сортам Юнона и Коллега. Наиболее длинный колос был у сорта Коллега (10,6 см), а короткий у – Юноны (9,3 см).
длина колоса у родительских форм была довольно стабильной, коэффициент вариации сортов составил 9,2 и 10,3 % соответственно. Пределы изменчивости данного признака у гибрида во втором поколении колебались от 6 до 14 см. Исходя из данных показателей, коэффициент варьирования был довольно высоким – 17,9%. В гибридных популяциях имелись растения, как с коротким колосом, так и с очень длинным. По данному изучаемому признаку снова проявился отцовский эффект.
По признаку длина колоса средние показатели у гибридных растений первого поколения были выше, чем у родительских форм, а второго поколения – промежуточными между ними. В F1 была отмечена средняя изменчивость данного признака у полученного гибрида.
Таблица 20 – Характеристика гибридов и родительских форм по длине
колоса
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, см | Lim, см | V,% | Х+Sx, см | Lim, см | V,% | |
| Р.ф. Батько | 11,3±0,23 | 9–14 | 10,1 | 9,8±0,19 | 8–12,3 | 9,8 |
| Батько х Аруана | 12,4±0,31 | 9–15 | 12,8 | 9,5±0,11 | 5–14,5 | 18,1 |
| Р.ф. Аруана | 11,6±0,41 | 7–15 | 16,6 | 11,5±0,32 | 9–14,5 | 14,2 |
Пределы изменчивости данного признака во втором поколении колебались у гибрида Батько х Аруана от 5 до 14,5 см. Исходя из данных показателей, коэффициент варьирования также был выше и составил 18,1%. В гибридной популяции имелись растения, как с коротким колосом, так и с очень длинным. Однако, эти значения не выходили за пределы размаха варьирования родительских форм, т.е. длина колоса у полученных гибридов была промежуточной по отношению к родительским формам. Причем по данному изучаемому признаку проявился материнский эффект.
Пределы изменчивости длины колоса во втором поколении колебались у гибрида от 5,7 до 15,2 см. В гибридных популяциях имелись растения, как с коротким колосом, так и с очень длинным. Однако, эти значения выходили за пределы размаха варьирования родительских форм. Это еще раз подтвердило наше предположение, о том, что на длину колоса в большей степени оказывает влияние материнская форма, взятая для скрещивания.
Результаты анализа гибридных популяций и их родительских форм по количеству колосков в колосе представлены в таблицах 21 и 22.
Таблица 21 – Характеристика гибридов и родительских форм по количеству колосков в колосе
| Гибрид, родительские формы | Х+Sx, шт. | Lim, шт. | V, % | σ2 |
| Р.ф. Коллега | 21,5 + 0,22 | 19,0-23,0 | 5,0 | 1,0 |
| Коллега х Юнона | 18,7 + 0,09 | 16,0-23,0 | 9,3 | 3,10 |
| Р.ф. Юнона | 19,0 + 0,24 | 17,0-22,0 | 6,4 | 1,2 |
Данные таблицы говорят о том, что Коллега имеет более плотный колос, чем Юнона и формирует большее число колосков в колосе. Полученный гибрид близок по своим средним значениям к родительской форме Юнона. Размах варьирования составил 16,0-23,0 колосков в колосе. Коэффициент вариации у гибрида Коллега х Юнона оказался выше, чем у родительских форм. Таким образом, полученные данные свидетельствуют о том, что в полученной гибридной популяции можно вести отбор особенно ценных форм с плотным колосом.
В целом, по признаку число колосков в колосе положительная трансгрессия наблюдалась по отношению к родительской форме Юнона, т.к. она имела невысокие показатели числа колосков в колосе. Благодаря высокому числу колосков в колосе у Коллеги, в потомстве гибрида были обнаружены растения с числом колосков больше 22 шт.
Таблица 22 – Характеристика гибридов и родительских форм по количеству колосков в колосе
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, шт. | Lim, шт. | V,% | Х+Sx, шт. | Lim, шт. | V,% | |
| Р.ф. Батько | 21,9±0,35 | 17-25 | 7,7 | 19,8±0,20 | 19-22 | 5,1 |
| Батько х Аруана | 19,8±0,41 | 14–24 | 10,3 | 19,7±0,11 | 15-24 | 8,8 |
| Р.ф. Аруана | 19,1±0,49 | 12-22 | 12,2 | 18,4±0,31 | 15-22 | 8,4 |
Данные таблицы говорят о том, что Батько имеет более плотный колос, чем Аруана и формирует большее число колосков в колосе. Полученные гибриды и в первом и во втором поколениях близки по своим средним значениям к родительской форме Батько. Размах их варьирования и в первом и во втором поколениях составил примерно одинаковые значения и составил для F1 14–24 шт., для F2 15,6–24,6 шт. Соответственно коэффициент вариации у гибрида F2 оказался выше, чем в первом поколении. У полученных нами гибридов первого поколения наблюдалась положительная трансгрессия по отношению к родительской форме Аруана.
По признаку число колосков в колосе положительная трансгрессия наблюдалась по отношению к родительской форме Аруана, а наибольшее влияние на формирование данного признака оказала материнская родительская форма – Батько.
Таким образом, полученные данные свидетельствуют о том, что в полученных гибридных популяциях можно вести отбор особенно ценных форм с плотным колосом.
Данные анализа гибридных популяций и их родительских форм по числу зерен в колосе представлены в таблицах 23 и 24.
Средние значения родительских форм по этому признаку составили 58,2 и 61,3 шт. Наибольший размах вариации был отмечен у Юноны.
Таблица 23 – Характеристика гибридов и родительских форм по числу зерен в колосе
| Гибрид, родительские формы | Х+Sx, шт. | Lim, шт. | V,% | σ2 |
| Р.ф. Коллега | 58,2 + 2,31 | 37,0-76,0 | 19,8 | 11,5 |
| Коллега х Юнона | 54,4 + 0,66 | 35,0-82,0 | 24,5 | 123,97 |
| Р.ф. Юнона | 61,3+ 2,73 | 30,0-80,0 | 20,9 | 14,5 |
Гибриды второго поколения имели показатели числа зерен в колосе меньше родительских форм, а размах их варьирования оказался схожим с материнской и отцовской формой. Наибольшее влияние на наследование этого признака у гибрида оказала материнская форма Коллега. Коэффициент вариации и дисперсия у гибрида по изучаемому признаку дает возможность на отбор хорошо озерненных колосьев для дальнейшей селекции.
Таблица 24 – Характеристика гибридов и родительских форм по числу зерен в колосе
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, шт. | Lim, шт. | V,% | Х+Sx, шт. | Lim, шт. | V,% | |
| Р.ф. Батько | 48,2±2,07 | 31-70 | 20,6 | 41,5±1,16 | 30-55 | 14,0 |
| Батько х Аруана | 52,4±2,03 | 31-70 | 19,4 | 35,2±0,61 | 25-58 | 20,1 |
| Р.ф. Аруана | 40,1±2,24 | 24-54 | 20,4 | 35,7±1,94 | 25-50 | 19,8 |
Средние значения родительских форм по этому признаку в первом поколении составили 40,1 и 48,2 шт. Наибольший размах вариации был отмечен у Батько. Гибрид первого поколения сформировал в среднем больше зерен в колосе, чем его родительские формы. Размах варьирования этого признака был довольно большим и составил 19,4%. однако, варьирование значений родителей в опыте было даже выше, чем у гибрида.
При изучении второго поколения у всех исследуемых форм среднее число зерен в колосе оказалось ниже, по сравнению с предыдущим годом. Это связано с неблагоприятными погодными условиями, сложившимися во время налива и созревания зерна озимой пшеницы.
Наибольший размах вариации по числу зерен в колосе был отмечен у гибрида Батько х Аруана. Гибриды второго поколения имели показатели числа зерен в колосе близкие к Аруане, а размах их варьирования оказался даже выше, чем у родительских форм. По данному признаку была отмечена положительная трансгрессия по сравнению с родительской формой Аруана.
Результаты анализа гибридных популяций и их родительских форм по массе зерна с колоса представлены в таблицах 25 и 26.
Таблица 25 – Характеристика гибридов и родительских форм по массе зерна с колоса
| Гибрид, родительские формы | Х+Sx, г | Lim, г | V, % | σ2 |
| Р.ф. Коллега | 2,54 +0,12 | 1,30-3,50 | 22,5 | 0,70 |
| Коллега х Юнона | 1,71 + 0,03 | 1,00-3,70 | 30,3 | 0,33 |
| Р.ф. Юнона | 2,05 +0,11 | 0,70-3,10 | 26,5 | 0,50 |
Данные таблицы показывают, что масса зерна с колоса у родительских форм при изучении F2 была выше 2,0 грамм, причем родительская форма Коллега имела наибольшее значение (2,6 г). У Коллеги отмечен высокий размах изменчивости (1,3-3,5 г), и коэффициент вариации (22,5%). Родительская форма Юнона характеризовалась лимитами варьирования от 0,7 до 3,1 г при коэффициенте вариации 26,5%.
Полученный гибрид второго поколения имел значения массы зерна с колоса ниже, чем его родители. Верхние значения лимита у него достигали 3,7 г, что говорит о достаточной крупности зерна у некоторых растений. Коэффициент изменчивости данного гибрида составил 30,3%, что позволит выделить нам наиболее интересные растения для селекции.
Таблица 26 – Характеристика гибридов и родительских форм по массе зерна с колоса
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, г | Lim, г | V,% | Х+Sx, г | Lim, г | V,% | |
| Р.ф. Батько | 2,09±0,11 | 1,24–3,35 | 25,2 | 1,72±0,06 | 1,30–2,60 | 18,8 |
| Батько х Аруана | 2,75±0,14 | 1,18–3,90 | 26,0 | 1,68±0,03 | 1,00–2,90 | 24,2 |
| Р.ф. Аруана | 2,15±0,13 | 1,11–3,04 | 28,5 | 1,85±0,08 | 1,00–2,70 | 22,6 |
Данные таблицы показывают, что масса зерна с колоса у родительских форм при изучении F1 была ниже, чем у гибридной комбинации Батько х Аруана, родительские формы имели примерно одинаковые значение (2,09 и 2,15 г). У Аруаны же отмечен и самый высокий коэффициент вариации (28,5%).
Полученный гибрид второго поколения имел значения массы зерна с колоса ниже, чем у родителей. Верхние значения лимита у него достигали 2,9 г, что говорит о достаточной крупности зерна. Коэффициент изменчивости данного гибрида составил 24,4 %, что позволило выделить нам наиболее интересные растения для селекции. Полученный гибрид во втором поколении показал промежуточные значения массы зерна, чем у родительских форм.
Данные анализа гибридных популяций и их родительских форм по массе 1000 зерен представлены в таблицах 27 и 28.
Таблица 27 – Характеристика гибридов и родительских форм по массе 1000 зерен
| Гибрид, родительские формы | Х+Sx, г | Lim, г | V, % | σ2 |
| Р.ф. Коллега | 44,5 + 1,62 | 35,9-79,0 | 18,3 | 11,8 |
| Коллега х Юнона | 41,8 + 0,33 | 26,8-63,0 | 16,0 | 44,87 |
| Р.ф. Юнона | 31,6 + 0,70 | 22,0-37,3 | 11,1 | 3,5 |
Наибольшая масса 1000 зерен оказалась у сорта Коллега (44,5 г), у родительской формы Юнона она была значительно ниже – 31,6 г. Полученный гибрид второго поколения имел массу 1000 зерен, близкую к Коллеге (41,8 г). Поэтому можно сделать вывод, что он оказал большее влияние на наследование этого признака у гибрида. Коэффициент изменчивости данного признака был сравнительно небольшим – 16 % и имел промежуточные значения между родительскими формами. Наименьшей изменчивостью этого признака обладала Юнона (11,1% при размахе варьирования от 22,0 г до 37,3 г). У коллеги крайние значения предела варьирования были довольно высокими 35,9-79,0 г. Это и способствовало появлению в потомстве гибрида растений с массой 1000 зерен более 60 г.
Таблица 28 – Характеристика гибридов и родительских форм по массе 1000 зерен
| Гибрид, родительские формы | F1 | F2 | ||||
| Х+Sx, г | Lim, г | V,% | Х+Sx, г | Lim, г | V,% | |
| Р.ф. Батько | 43,2±0,76 | 34,9-48,9 | 8,5 | 41,5±0,96 | 32,1-51 | 11,6 |
| Батько х Аруана | 51,8±1,06 | 38,1–58,4 | 10,3 | 41,7±0,51 | 30,0-64,9 | 16,6 |
| Р.ф. Аруана | 53,1±4,21 | 39,6-66,2 | 12,3 | 51,6±0,56 | 45,0-55,3 | 5,4 |
Родительские формы имели среднюю массу 1000 зерен выше 40,0 г, причем у Аруаны она была значительно выше (53,1 г). Полученный гибрид первого поколения имел массу 1000 зерен, близкую к сорту Аруана. Поэтому можно сделать вывод, что она оказала большее влияние на наследование этого признака. Коэффициент изменчивости данного признака у полученного гибрида первого поколения был небольшим (10,3%).
Во втором поколении средняя масса 1000 зерен полученного гибрида была ниже по сравнению с первым поколением. У гибрида второго поколения отмечен и самый высокий размах изменчивости (30,0–64,9 г), и коэффициент вариации (16,6%).
Вариабельность гибрида второго поколения была выше, чем у первого поколения изучаемого гибрида, это связано с расщеплением в потомстве.
Абсолютные величины элементов продуктивности сортов пшеницы подвержены изменчивости по годам, однако, характер их соотношения сохраняется. В связи с этим необходимо было выяснить степень сопряженности отдельных признаков между собой у гибрида второго поколения на примере комбинации Коллега Х Юнона (таблица 29).
Таблица 29 – Корреляционные взаимосвязи некоторых признаков гибрида озимой пшеницы Коллега х Юнона
| Признаки | Высота растения | Длина колоса | Число колосков в колосе | Число зерен в колосе | Масса зерна с колоса | Масса 1000 зерен |
| Высота растения | 1 | | | | | |
| Длина колоса | 0,40 | 1 | | | | |
| Число колосков в колосе | 0,18 | 0,63 | 1 | | | |
| Число зерен в колосе | 0,25 | 0,70 | 0,62 | 1 | | |
| Масса зерна с колоса | 0,29 | 0,72 | 0,62 | 0,93 | 1 | |
| Масса 1000 зерен | 0,25 | 0,41 | 0,31 | 0,35 | 0,64 | 1 |
Для гибрида Коллега х Юнона характерна высокая сопряженность (r> 0,5) массы зерна и числа колосков и зерен с одного колоса между собой и с длиной колоса. Следует отметить, что связь между признаками: длина колоса и высота растения, длина колоса и масса 1000 зерен, число колосков в колосе и зерен в колосе и масса 1000 зерен слабая.
Таким образом, продуктивность колоса сортов озимой мягкой пшеницы взаимосвязана с массой колоса, массой зерна и числом зерен в нем. высокая взаимосвязь отмечена между длиной колоса и числом зерен в нем, а также массой зерна с колоса. Это дает возможность для отбора лучших перспективных форм по признаку «длина колоса» для дальнейшего изучения.
Для селекционной работы большое значение имеет определение характера наследования, установление количества генов, обуславливающих определенные количественные признаки и используемые в качестве их источников, взаимодействие этих генов, их эффективность при разных условиях внешней среды.
Формула прогноза гетерозиса