
Биоценотическая активность гликополимеров и состав основных метаболитов пресноводных высших растений в условиях загрязнения водоема 03. 00. 16 экология 03. 00. 04 биохимия
| Вид материала | Автореферат диссертации |
- Лекция 11 Тема: Высшие растения. Происхождение высших наземных растений. Отдел Моховидные, 276.22kb.
- Регенерационная активность разных генотипов пшеницы и эгилопса в культуре in vitro, 499.46kb.
- Культура столонов и регуляция роста растений и клубнеобразования у картофеля in vitro, 342.87kb.
- Изучение биологических особенностей растений в новых условиях, 533.64kb.
- Рабочей программы учебной дисциплины «экология растений, животных и микроорганизмов», 31.83kb.
- Программа курса Методика организации курса \"Экология Тамбовской области\", 78.43kb.
- План по теме «Анатомия и морфология высших растений» Цель курса, 22.96kb.
- Тема : Жизнь пресного водоёма. Цель, 144.23kb.
- Эпигенетическая регуляция фотосинтетической ассимиляции со 2 у хлопчатника (03. 00., 544.35kb.
- Фёдорова Светлана Владиславовна Структура и организация популяций ряда наземно-ползучих, 312.38kb.
ксильных групп с максимумами около 3400 см-1. Полоса при 2923 см-1 во всех спектрах ПС была вызвана валентными колебаниями метильных групп в остатках рамнозы и фукозы (н-ПС-1), а при 1743 см-1 – валентными колебаниями карбонила в сложноэфирной группе. Интересно, что в спектрах к-ПС-3 и к-ПС-4 сложноэфирная группа обнаруживала себя слабой полосой при 1734 см-1, в спектрах остальных ПС эта полоса отсутствовала. Т.к. к-ПС рдеста состоят только из нейтральных сахаров, то, скорее всего, их заряд обусловлен наличием неуглеводных анионных заместителей. В спектре н-ПС-1 в низкочастотной области наблюдается поглощение средней интенсивности с максимумом при 834 см-1, на основе известных корреляций позволяет предположить, что связывание моносахаридных остатков в основном происходит за счет -гликозидных связей (Жбанков, 1972). Нужно сказать, имеются единичные работы относительно ПС состава представителей рдестовых, в частности, определена структура и выявлена иммуномодулирующая активность пектиновых ПС Potamogeton natans L. (Popov et al., 2007). Полученные нами данные расширяют представление о составе растворимых гликополимеров растений семейства рдестовых.
Глава 5. РОЛЬ ПОЛИСАХАРИДОВ РДЕСТА ПРОНЗЁННОЛИСТНОГО В
ФОРМИРОВАНИИ ЕГО БАКТЕРИАЛЬНОГО ОКРУЖЕНИЯ
Существенную роль в самоочищении пресноводных водоемов играют растительно-микробные ассоциации прибрежной мелководной зоны (Van Donk, van de Bund, 2002; Chang et al., 2006), однако, механизм и метаболиты, являющиеся наиболее значимыми для их формирования, остаются малоизученными (Звягинцев и др., 1993; Xie, Yokota, 2005). В этой связи актуальной является оценка роли ПС в формировании макрофитно-бактериальных ассоциаций. Рдест пронзённолистный является одним из основных ценозообразователей среди погруженных пресноводных макрофитов (Лукина, Смирнова, 1988).
Одним из тестов, позволяющих наблюдать взаимодействие бактерий с растениями, является хемотаксис (рис. 2). Из всех микроорганизмов, используемых в работе, индифферентной ко всем ПС (все гликаны вносили в концентрации 10-3 мМ) была культура E.coli K12, поэтому в экспериментах она выполняла роль отрицательного контроля. Следует отметить, что различия диаметров хемотаксических колец, наблюдаемых в экспериментах, были незначительными. Положительную реакцию по отношению к нейтральным ПС проявляли ассоциативные бактерии A. brasilense Sp245 и P. fluorescens B-1471 (диаметр хемотаксических колец от 10 до 12 мм), а к кислым ПС – фитопатоген X. campestris B-610 (диаметр от 8 до 10 мм) (рис. 3).
Микроорганизмы Препараты
аттрактанты
н-ПС-1
A. brasilense Sp245 н-ПС-2
к-ПС-1
н-ПС-2
P. fluorescens B-1471 к-ПС-4
к-ПС-1
к-ПС-3
X. campestris B-610 к-ПС-4
к-ПС-5
к-ПС-3
M. luteus B-109 к-ПС-4
Рис. 3. Схема таксиса бактерий по отношению к полисахаридам рдеста пронзеннолистного
А Б



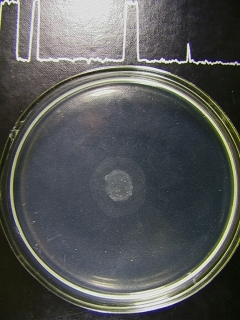

Рис. 2. Хемотаксис A. brasilense Sp245 к
полисахаридам рдеста пронзённолистного:
А
 – наличие таксиса к препарату н-ПС-2 рдеста;
– наличие таксиса к препарату н-ПС-2 рдеста;Б – отсутствие таксиса к препарату к-ПС-4
Стрелкой указана ширина кольца таксиса
Таким образом, выявлена положительная реакция таксиса бактерий по отношению к некоторым из ПС рдеста. Вопрос о механизме таксиса бактерий к исследуемым ПС остается открытым. Возможно, полисахаридсодержащие фракции рдеста могут вызывать конформационные изменения рецепторов или иных структур поверхности клетки при непосредственном связывании с ними. Количество таких «трансформированных» рецепторов является первичным сигналом для хемотаксической системы бактериальной клетки, действие которой отражается на двигательной активности бактерий (Брезгунов и др., 1989).
Взаимодействие ПС с клеточной стенкой может приводить к изменению гидрофобности поверхности микроорганизмов. Гидрофобность является одной из ключевых характеристик клеточной поверхности, которая лежит в основе ряда биологических процессов и может оказать существенное влияние на образование симбиотических и ассоциативных сообществ (Никовская и др., 1985). Для выявления таких изменений была исследована относительная гидрофобность поверхности изучаемых бактерий, обработанных полисахаридными препаратами рдеста, в концентрации 4х10 4 мМ (концентрация ПС была подобрана на основе анализа литературных данных, а также в ходе предварительных экспериментов). Для исследований использовали тест солевой агрегации, который позволяет оценить, главным образом, вклад углеводных компонентов в формирование относительной гидрофобности поверхности бактерий (Dillon et al., 1986). Результаты исследований, представленные в табл. 2, показали, что в 70% экспериментов наблюдалось достоверное изменение относитель ной гидрофобности поверхности бактерий, обработанных раствором ПС.
Таблица 2
Влияние полисахаридов рдеста пронзённолистного на относительную гидрофобность клеточной поверхности микроорганизмов
| Препараты | Индекс гидрофобности, (%) | |||
| X. campestris B-610 | P. fluorescens B-1417 | A. brasilense Sp245 | M. luteus B-109 | |
| Контроль | 49,0 1,5 | 58,0 4,5 | 34,0 1,5 | 65,5 4,5 |
| н-ПС-1 | 44,5 1,5* | 53,5 3,0 | 43,0 1,5* | 41,5 3,0* |
| н-ПС-2 | 47,5 3,0 | 67,0 1,5* | 37,0 3,0 | 46,0 3,0* |
| к-ПС-1 | 35,5 1,5* | 44,5 1,5* | 40,0 4,5* | 56,5 1,5* |
| к-ПС-3 | 32,5 1,5* | 58,0 3,0 | 19,0 1,5* | 59,5 3,0 |
| к-ПС-4 | 34,0 3,0* | 53,5 1,5 | 26,5 3,0* | 64,0 3,0 |
| к-ПС-5 | 44,5 1,5* | 62,5 1,5 | 22,0 4,5* | 52,0 1,5* |
* отличия параметра от контрольных значений достоверны
Полученные результаты могут свидетельствовать о наличии связывания расти-тельных ПС с бактериальной поверхностью, вследствие чего и наблюдается изменение гидрофобности. Характер изменения гидрофобности зависит от ряда причин, в том числе и от моносахаридного состава используемых в эксперименте ПС. Наблюдаемое в большинстве опытов понижение индекса показало, что относительная гидрофобность поверхности бактерий за счет взаимодействия с ПС повышалась. Возможно, повышение гидрофобности объясняется присутствием в составе большинства исследуемых ПС дезоксисахара – рамнозы. Наибольший эффект наблюдался при воздействии к-ПС-3 на ксантомонады и азоспириллы. Так как увеличение относительной гидрофобности бактериальной поверхности микроорганизмов, как правило, способствует их адгезии, возможно обогащение микроокружения рдеста этими видами микроорганизмов. На следующем этапе работы был проведён анализ влияния ПС рдеста на рост бактерий на агаризованной питательной среде (табл. 3).
Таблица 3
Влияние полисахаридов рдеста пронзённолистного на рост микроорганизмов
| Тест-культуры | Вносимый препарат | Наблюдаемый эффект | Диаметр зоны эффекта, мм |
| X. campestris B-610 | н-ПС-1 | Ингибирование роста | 12,3 ± 0,1 |
| н-ПС-2 | Ингибирование роста | 12,5 ± 0,5 | |
| к-ПС-1 | Усиление роста | 11,0 ± 2,0 | |
| к-ПС-3 | Усиление роста | 7,0 ± 0,5 | |
| к-ПС-4 | Усиление роста | 15,0 ± 0,2 | |
| P. fluorescens B-1417 | н-ПС-2 | Усиление роста | 5,3 ± 0,3 |
| к-ПС-4 | Усиление роста | 12,0 ± 0,6 | |
| к-ПС-5 | Усиление роста | 3,0 ± 0,2 | |
| A. brasilense Sp245 | н-ПС-2 | Усиление роста | 12,0 ± 0,3 |
| к-ПС-4 | Ингибирование роста | 1,5 ± 0,6 | |
| M. luteus B-109 | к-ПС-3 | Усиление роста | 2,0 ± 0,6 |
| к-ПС-4 | Усиление роста | 3,0 ± 0,4 |
Учитывая, что таксис, как правило, обусловлен трофическими связями, мы предположили, что некоторые из исследуемых организмов могут использовать эти ПС в качестве источника углерода. Анализы показали, что н-ПС-1 и н-ПС-2 в концентрации 4х10-4 мМ ингибировали рост фитопатогена X. campestris B-610, для остальных ПС значимого подавление роста бактерий не обнаружено, напротив, наблюдались зоны увеличения плотности роста колоний вокруг места внесения препарата. Следует отметить, что данные таблицы 3, в основном, коррелируют с ранее полученными результатами исследования хемотаксиса (см. рис. 3). Исключением является отсутствие влияния на рост M. luteus B-109 препарата к-ПС-3, и небольшой эффект и к-ПС-4. Особенность этих ПС состоит в том, что оба они, являясь аттрактантами для этого микроорганизма, не влияют на гидрофобность его поверхности. Эффект индуцирования роста бактерий, возможно, объясняется наличием у этих микроорганизмов ферментных систем, позволяющих использовать исследуемые ПС как источник углерода. Механизм подавляющего действия ПС на рост бактерий изучен слабо, однако для некоторых н-ПС показано, что вследствие связывания углеводсодержащих полимеров со специфическими рецепторами клеточной стенки бактерий происходило нарушение проницаемости внешней мембраны микроорганизмов до пределов, несовместимых с жизнью. Возможно, это связывание приводило к существенным изменениям физико-химических свойств поверхности, что, в свою очередь, изменяло конформацию поверхностных белков и, как следствие, нарушало их функции (Vaara, 1992; Gil1 et.al., 2004).
В настоящее время исследуется физиологическая активность растительных ПС по отношению к про- и эукариотическим клеткам. Влияние растительных ПС на скорость прорастания семян и клеточных культур растений широко обсуждается в литературе (Гюнтер, Оводов, 2001; Елькина и др., 2002). Мало известно о влиянии ПС ВВР на микроорганизмы. Нами на примере бактерий A. brasilense Sp245 показана важная роль капсульных ПС для выживания бактерий в неблагоприятных условиях. Проведенные эксперименты показали, что добавление препаратов ПС к бескапсульным клеткам A. brasilense Sp245, существенно снижало воздействие экстремальных факторов окружающей среды (температуры, рН, высушивания) и увеличивало выживаемость бактерий на 17 – 50%.
Исследование динамики роста трёх видов бактерий в присутствии ПС рдеста в концентрации 4х10-4 мМ показало, что к-ПС-4 в среде выращивания X. campestris B-610 стимулирует накопление биомассы в среднем на 26%, а для A. brasilense Sp245 наблюдается подавление накопления биомассы на 27%. Достоверного влияния к-ПС-4 на динамику роста M. luteus B-109 не выявлено. Н-ПС-2 подавлял накопление биомассы ксантомонад и незначительно, на некоторых этапах, стимулировал деление клеток азоспирилл. Присутствие к-ПС-5 сокращало для M. luteus B-109 время достижения культурой фазы окончания экспоненциального роста. Все обнаруженные эффекты наиболее ярко проявляются после 11 – 13 часов выращивания культур. Полученные результаты этого эксперимента согласуются с данными по хемотаксису и росту клеток на агаризованной среде. На основании результатов проведенных опытов можно говорить об участии выделенных ПС рдеста пронзённолистного в формировании видового состава его бактериального окружения.
Глава 6. ИЗУЧЕНИЕ ВЛИЯНИЯ ПОЛИСАХАРИДА РДЕСТА ПРОНЗЁННЛИСТНОГО IN VITRO НА ИНДУКЦИЮ СИНТЕЗА ЦИТОКИНОВ ФАГОЦИТИРУЮЩИМИ МАКРОФАГАМИ
Известно, что ПС растительного происхождения являются индукторами ответных реакций макроорганизма, в том числе и синтеза цитокинов. В настоящее время в медицинской практике в качестве иммуностимуляторов и иммуномодуляторов используются препараты на основе растительных ПС (Ройт и др., 2002). Механизм действия ПС на организм человека и животных еще не достаточно изучен. Есть лишь некоторые отрывочные сведения о спектре биологической активности ПС различных растений (Турова, Гладких, 1965; Хотимченко и др., 2005). Исследование влияния растительных ПС на про- и эукариотические клетки остается весьма актуальным.
Проведено изучение влияния ПС рдеста in vitro на активность фагоцитоза эшерихий и индукцию провоспалительных эндогенных цитокинов макрофагами мышей. В качестве объекта исследования использовали н-ПС-1 рдеста пронзённолистного. Сложный моносахаридный состав ПС позволяет предположить возможность взаимодействия этого гликополимера с соответствующими рецепторами на фагоцитах.
Добавление н-ПС-1 в концентрации 0,01 мкг/мл в культуру макрофагов приводило к незначительному увеличению количества активированных клеток по сравнению с контролем в интервале 1 – 4 ч фагоцитоза (на 10% для АМФ и на 15 – 18% для ПМФ), к 6 ч значения фагоцитарного индекса (ФИ) соответствовали контрольным. Препарат в дозе 0,1 мкг/мл усиливал по сравнению с контролем активность только АМФ на 11 и 13% к 2 и 6 ч процесса фагоцитоза соответственно. Внесение н-ПС-1 в дозе 1 мкг/мл приводило к значительной активации АМФ и ПМФ в течение всего процесса фагоцитоза. ФИ на всех этапах фагоцитоза E. сoli были выше по сравнению с контролем для АМФ на 14 – 20%, а для ПМФ – на 23 – 33%. Динамика фагоцитоза эшерихий АМФ и ПМФ на фоне действия ПС в дозе 0,01 и 1 мкг/мл была сходной.
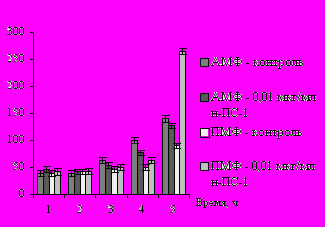
В ходе исследования влияния препарата на индукцию синтеза цитокинов показано, что добавление н-ПС-1 в концентрации 0,01 мкг/мл в культуру макрофагов приводило к синтезу интерлейкина-1-β (ИЛ-1-β) в пределах контроля. Препарат н-ПС-1 в дозах 0,1 и 1 мкг/мл существенно индуцировал продукцию этого цитокина, и к 4, 6 и 12 ч концентрация ИЛ-1-β повышалась в 5 – 10 раз по сравнению с контролем для АМФ и ПМФ (рис. 4).
Содержание фактора некроза опухоли-a (ФНО-a) в опытных пробах при всех исследуемых дозах ПС было выше в 2,5 – 5 раз по сравнению с контрольными значениями. Однако зависимость индукции ФНО-a фагоцитирующими макрофагами от концентрации ПС не была выявлена.
Таким образом показано влияние н-ПС-1 рдеста пронзённолистного in vitro на активность процесса фагоцитоза эшерихий и индукцию синтеза провоспалительных цитокинов – ИЛ-1-β и ФНО-a макрофагами мыши. Проведённые эксперименты свидетельствуют о том, что этот препарат может обладать способностью к индукции неспецифической резистентности теплокровных животных и человека.
А Б
1 2 4 6 12
1 2 4 6 12
Концентрация,
пг/мл
Концентрация,
пг/мл
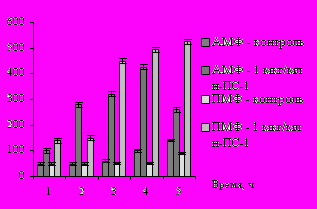
Рис. 4. Синтез ИЛ-1-β макрофагами мышей при добавлении полисахарида рдеста пронзённолистного н-ПС-1 in vitro в дозе 0,01 и 1 мкг/мл в ходе фагоцитоза клеток E. coli Ca 52: А – ИЛ-1-β (0,01 мкг/мл н-ПС-1); Б – ИЛ-1-β (1 мкг/мл н-ПС-1)
Выводы
- Присутствующие в городских стоках соединения азота, хлориды, фториды и углеводороды приводят к существенным изменениям химического состава биомассы погруженных и полупогруженных высших водных растений.
- Выявлено изменение соотношения липидов и жирных кислот высших водных растений в условиях стресса, вызванного загрязнением воды: увеличение количества липидов у элодеи канадской и рдеста пронзённолистного ~ в 1,25 раза; у рдеста – повышение содержания непредельных кислот на 34%, а у элодеи и тростника – понижение на 45 и 24% соответственно. Изменение состава и соотношения ключевых липидов в биомассе растений и содержание октадекадиеновой (С18:29,12) кислоты может иметь биоиндикационное значение как показатель первичных реакций водных растений на присутствие выявленного спектра загрязнителей.
- Показано нарастание в биомассе суммарного количества свободных аминокислот в ответ на загрязнение воды: у рдеста пронзённолистного на 16%, у тростника обыкновенного на 36%, у элодеи канадской на 28%. Повышение содержания аминокислот в биомассе растений происходит, в основном, за счет дикарбоновых кислот, пролина и монокарбоновых аминокислот с гидрофобными радикалами.
- Обнаружено влияние загрязнения воды на моносахаридный состав высокомолекулярных углеводсодержащих веществ биомассы растений. Впервые из биомассы рдеста пронзённолистного выделены и охарактеризованы шесть гетерополисахаридов, различающиеся по молекулярной массе, моносахаридному составу и активности в отношении бактерий.
- Впервые показано, что полисахариды рдеста влияют на жизнеспособность, гидрофобность поверхности и таксис ряда потенциальных симбионтов из числа грамотрицательных и грамположительных бактерий. Показано участие исследуемых полисахаридов высших водных растений в формировании бактериоценоза.
- Впервые обнаружено влияние полисахарида рдеста пронзённолистного in vitro на активность процесса фагоцитоза эшерихий и индукцию синтеза провоспалительных цитокинов – ИЛ-1-β и ФНО-α. Показано, что полисахарид рдеста усиливает синтез ИЛ-1-β в 5 – 10 раз, а ФНО-α в 2,5 – 5 раз по сравнению с контролем.
Список публикций по теме диссертации
* – публикация в изданиях перечня ВАК РФ
1. *Коннова С.А., Брыкова О.С., Сачкова (Фучеджи) О.А, Егоренкова И.В., Игнатов В.В. Исследование защитной роли полисахаридсодержащих компонентов капсулы бактерий Azospirillum brasilense // Микробиология. – 2001. – Т. 70, №4. – С. 503 – 508.
2. Konnova S.A., Brykova O.S., Sachkova (Fuchedzhi) O.A., Ignatov V.V. Role of Azospirillum capsular material in bacterial resistance to extreme environmental factors // Abstract Book of 13th International Conference on Nitrogen Fixation. Hamilton Canada. – 2001. – P. 70.
3. Коннова С.А., Сачкова (Фучеджи) О.А., Игнатов В.В. Исследование углеводов и свободных аминокислот рдеста пронзённолистного // Фундаментальные и прикладные аспекты функционирования водных экосистем: проблемы и перспективы гидробиологии и ихтиологии в ХХI веке: сборник тезисов / Всероссийская научная конф. Саратов, 27 – 30 августа 2001. – Саратов, 2001. – С. 81 – 84.
4. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Влияние загрязнения воды на биохимический состав макрофитов // Биология – наука XXI века: сборник тезисов / 6-я Пущинская школа - конф. молодых ученых. Пущино, 20 – 24 мая 2002. – Пущино, 2002. – С. 332 – 333.
5. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Полисахариды и свободные аминокислоты растений // Химия и технология растительных веществ: сборник тезисов / II Всеросийсская конф. Казань, 24 – 27 июня 2002. – Казань, 2002. – С. 89 – 90.
6. Sachkova (Fuchedzhi) O.A., Konnova S.A. , Ignatov V.V. Change in the glycan content of Potamogeton perfoliatus as a response to water pollution // Abstract Book of International Symposium on Biochemical interactions of Microorganisms and Plants with technogenic environmental pollutants. Saratov Russia. – 2003. – P. 37.
7. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Бактерицидные свойства полисахаридов Potamogeton perfoliatus // Биология – наука XXI века: сборник тезисов / 8-я Пущинская школа - конф. молодых ученых. Пущино, 17 – 21 мая 2004. – Пущино, 2004. – С. 30.
8. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Исследование активности полисахаридов Potamogeton perfoliatus по отношению к микроорганизмам // Биология – наука XXI века: сборник тезисов / 9-я Пущинская школа – конф. молодых ученых. Пущино, 18 – 22 апреля 2005. – Пущино, 2005. – С. 211.
9. Сачкова (Фучеджи) O.A., Koннова С.A., Игнатов В.В. Изменение состава гликанов тростника обыкновенного в ответ на загрязнение воды // Проблемы биодеструкции техногенных загрязнителей окружающей среды: сборник тезисов / Международная конф. Саратов, 14 – 16 сентября 2005. – Саратов, 2005. – С. 145 – 146.
10. Бойко А.С., Сачкова (Фучеджи) О.А. Исследование активности гликанов рдеста пронзённолистного во взаимодействии с мироорганизмами // Исследования молодых ученых и студентов в биологии: Сборник научных статей. – Саратов: Изд-во СГУ, 2005. – Вып. 3. – С. 19 – 22.
11. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В., Мельников Г.В. Исследование углеводов и свободных аминокислот высших водных растений // Известия Саратовского университета. Серия Химия. Биология. Экология. – 2005. – Т. 5. – Вып. 2. – С. 3 – 7.
12. Коннова С.А., Сачкова (Фучеджи) О.А., Коннова О.Н., Федоненко Ю.П. Выделение и анализ гликополимеров растительного и бактериального происхождения / Учебное пособие для студентов биологического факультета. – Саратов: Изд-во СГУ, 2005. – 39с.
13. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Бактерицидные свойства гликополимеров рдеста пронзённолистного во взаимодействии с микроорганизмами // Стратегия взаимодействия микроорганизмов с окружающей средой: Сборник научных статей. Саратов: Изд-во Научная книга, 2005. – С. 75 – 81.
14. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Влияние загрязнения воды на состав жирных кислот высших водных растений // Биология – наука XXI века: сборник тезисов / 10-я Пущинская школа – конф. молодых ученых. Пущино, 17 – 21 апреля 2006. – Пущино, 2006. – С. 311.
15. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Изменение химического состава биомассы высших водных растений в ответ на загрязнение воды / IX Съезд Гидробиологического общества РАН. Тольятти, 18 – 22 сентября 2006. – Тольятти, 2006. – С. 142.
16. Сачкова (Фучеджи) О.А., Коннова С.А., Игнатов В.В. Исследование влияния полисахаридов Potamogeton perfoliatus на рост микроорганизмов // Химия и технология растительных веществ: сборник тезисов / IV Всероссийская научная конф. Сыктывкар, 26 – 30 июня 2006. – Сыктывкар, 2006. – С. 42.
17. Фомина А.А., Коннова О.Н., Фучеджи О.А. Влияние полисахаридов растительного и бактериального происхождения на индукцию цитокинов фагоцитирующими макрофагами // Исследования молодых ученых и студентов в биологии: Сборник научных статей. Саратов: Изд-во СГУ, 2006. – Вып. 4. – С. 107 – 110.
18. Фомина А.А., Фучеджи О.А. Влияние полисахаридов Potamogeton perfoliatus in vitro на активность фагоцитоза бактерий // Материалы I конференции молодых ученых медико-биологической секции поволжской ассоциации государственных университетов. Ульяновск, 27 апреля 2007. – Ульяновск, 2007. – С. 75 – 77.
19. *Фомина А.А., Фучеджи О.А., Коннова С.А., Тихомирова Е.И. Влияние полисахарида Potamogeton perfoliatus in vitro на активность фагоцитоза и индукцию цитокинов макрофагами // Медицинская иммунология. - 2007. – Т. 9, №2-3. – С. 168 – 169.
20. *Фучеджи О.А., Коннова С.А., Бойко А.С., Игнатов В.В. Участие гликополимеров в формировании бактериального гидроценоза рдеста пронзённолистного // Микробиология. – 2008. – Т. 77, №1. – С. 96 – 102.
__________________________________________________________________________
Подписано в печать 2008. Формат 60×84/16. Бумага офсетная.
Гарнитура Times. Печать RISO. Объем 1,0 печ. л. Тираж 100 экз. Заказ №
Отпечатано с готового оригинал-макета