
Курс лекций Составил: к т. н., доцент Тихонов А. И. 2002г
| Вид материала | Курс лекций |
СодержаниеДинамика экосистем 12.1. Виды динамики экосистем 12.2. Закономерности сукцессионного процесса 12.3. Стабильность и устойчивость экосистем |
- Курс лекций по дисциплине «Экономика малых предприятий» Составил: Курочкин, 1063.49kb.
- Курс лекций по предмету «Макроэкономика». Составил: ассистент кафедры, 1929.31kb.
- Курс лекций Барнаул 2001 удк 621. 385 Хмелев В. Н., Обложкина А. Д. Материаловедение, 1417.04kb.
- Книга организация управленческого труда (курс лекций), 2089.08kb.
- Матюшкин Александр Васильевич, кандидат филологических наук, доцент кафедры литературы., 123.47kb.
- Матюшкин Александр Васильевич, кандидат филологических наук, доцент кафедры литературы., 123.48kb.
- Курс лекций по автоматизированному электроприводу для итр проектный организаций с применением, 24.37kb.
- Курс лекций (28 часов) канд филос наук О. В. Аронсон Курс лекций «Математика и современная, 27.49kb.
- Курс лекций в электронной форме содержит все лекции предусмотренные программой дисциплины, 32.88kb.
- В. А. Деденко Л. Г. Караваев В. А. Курс лекций, 48.22kb.
Динамика экосистем
Цель: особенности жизненного цикла надсистем
Несмотря на то, что многие экологи не склонны относить экосистемы к “сверхорганизмам”, между развитием экосистем и развитием живых организмов имеется множество параллелей. Как и любые организмы, экосистемы рождаются, старятся и умирают, точнее, структурно перестраиваются или замещаются другими экосистемами. Выявлена масса закономерностей протекания этих процессов, которые своей повторяемостью свидетельствуют о наличии достаточно однозначных “программ” развития экосистем в том или ином регионе.
На каких носителях “записаны” эти программы? Что является генетическим кодом онтогенеза (индивидуальное развитие) экосистем? Как уже не раз отмечалось, ход развития определяется совокупностью факторов среды. В одинаковых условиях развиваются одинаковые по структуре экосистемы. Это своего рода давление надсистемы на эволюционирующую экосистему (макроуровень). Кроме того, следует учесть и характер законов природы, “зашитых” в формах организации материи, из которых строится экосистема, лежащих в основе всех механизмов природных явлений (микроуровень). Кто-то склонен переоценивать роль надсистемы в эволюции подсистем, считая, что развитие подсистем целиком подчинено требованиям более высокоуровневой организации (холистический подход). Кто-то, наоборот, категорически отрицает роль каких-либо надсистем, сводя всю динамику экосистем к механизмам межвидовых и внутривидовых отношений, генетической изменчивости, к борьбе за выживание и т.п. (редукционистский подход). Ни тот ни другой путь не дает полного понимания происходящих процессов, о чем свидетельствует бесконечный спор между сторонниками обеих концепций (аналогично спору между идеалистами и материалистами в философии).
К счастью, современная наука, похоже, открыла причину своих неудач в понимании мира. Принцип дополнительности находит признание и применение во все более широких областях знания. Только в единстве холизма и редукционизма возможно понимание сути явлений. Каждый подход по-своему прав, но только в их единстве возможно достижение полноты истины.
12.1. Виды динамики экосистем
Любая экосистема постоянно находится в состоянии динамики (изменений), вызванной как изменением факторов среды, то есть внешними возмущениями, так и внутренними процессами. При этом удобно различать циклическую динамику и поступательную. К разряду циклической динамики можно отнести:
1) суточную динамику, связанную с изменением активности растений и животных в течение суток: изменения в фотосинтезе, транспирации (испарение воды с поверхности листьев), у животных это смена периодов активности и сна и т.п.;
2) сезонную динамику (смена периодов вегетации и покоя), связанную с сезонными колебаниями: смена времен года, периодов дождей и засухи, муссонная периодичность и т.п.;
3) многолетнюю цикличность, вызванную, например, периодичностью солнечной активности (период порядка 10-11 лет), или же какими-то другими колебаниями с многолетним периодом, например, цикл Эль-Ниньо, который приводит к усилению ураганов, обрушивающихся на Южную Америку с периодичностью 5-7 лет, и т.п.
Особого внимания заслуживает поступательная динамика экосистем, или развитие. Обычно развитие сопровождается последовательным рядом изменений видовой и трофической структур экосистемы, всей ее организации, что позволяет даже говорить о последовательной смене экосистем, поэтому данный процесс называется сукцессией (от латинского слова сукцессио - преемственность, наследование). В зависимости от причин сукцессии различают экзодинамические (от греческого слова эксо - снаружи) сукцессии, вызванные внешними по отношению к данной экосистеме факторами, и эндодинамические (от греческого слова эндон - внутри) сукцессии, вызванные внутренними механизмами экосистемы (в теории управления различают вынужденную и свободную составляющие динамического процесса, которые соответствуют двум видам сукцессии экосистем).
Экзодинамические сукцессии могут быть вызваны изменениями климата, понижением уровня грунтовых вод, подъемом уровня мирового океана и т.п. Такие смены могут длиться столетиями и тысячелетиями. Они связаны в основном с действием механизмов адаптации экосистемы к факторам внешней среды, которые в свою очередь базируются на механизмах адаптации живых организмов экосистемы.
Эндодинамические сукцессии приводятся в действие особыми законами, механизмы которых до сих пор во многом непонятны. Известно, что на любом, даже абсолютно безжизненном, субстрате типа песчаных дюн или затвердевшей лавы, рано или поздно расцветает жизнь. При этом формы жизни, точнее, типы сообществ, в данном пространстве последовательно сменяют друг друга, постепенно усложняясь и увеличивая видовое разнообразие, формируя так называемый сукцессионный ряд, состоящий из последовательных стадий, отмечающих смену одного сообщества другим. Сукцессионный ряд заканчивается стадией зрелости, на которой экосистема изменяется очень мало. Экосистемы на этой стадии называются климаксными (от греческого слова климакс - лестница).
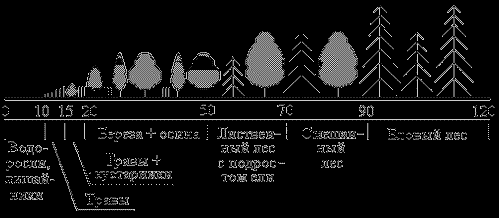 |
| Рис.23 |
Продолжительность сукцессии от зарождения экосистемы до стадии климакса может составлять до сотен и даже до тысяч лет. Столь длительная продолжительность связана в основном с необходимостью накопления питательного вещества в субстрате. Изначально на безжизненном субстрате поселяются так называемые пионерные виды, типа лишайников и корковых водорослей (рис.23). За 5-10 лет они несколько обогащают субстрат питательными веществами, формируя зачатки почвы. Затем на этих еще совсем бедных почвах поселяются травы, еще более обогащая почву. Лет через 15 от начала сукцессии на когда-то безжизненном пространстве поселяются первые кустарники, которые постепенно вытесняются лиственными светолюбивыми деревьями, чаще всего березой и осиной, которые характеризуются быстрым ростом. К 50-летнему возрасту в молодом лиственном лесу выделяются наиболее сильные деревья, которые затеняют более слабую поросль, которая погибает, давая возможность поселиться под пологом лиственного леса поросли ели. Ель более теневынослива, под защитой лиственных деревьев она постепенно догоняет их в росте, отвоевывая у них жизненное пространство. Где-то к 70 годам экосистема достигает стадии смешанного елово-лиственного леса. Лиственные деревья к тому времени успевают состариться, и постепенно ель выходит на первый ярус, затеняя и изреживая всю лиственную растительность. К 90 годам данная экосистема достигает стадии климакса, для которой характерно практически полное отсутствие лиственных деревьев, ель становится доминирующим видом-эдификатором, формируя особым образом всю жизнь населяющего данную экосистему сообщества.
Это типичный пример сукцессии, характерный для южной тайги. Как ни странно, в одном и том же регионе практически все сукцессии развиваются по довольно похожим сценариям и заканчиваются в итоге одним и тем же климаксом. Конечно, в зависимости от конкретных особенностей среды возможны некоторые вариации общего сценария, но они не принципиальны. Таким образом, каждый достаточно крупный регион характеризуется своей программой развития экосистем и своим особым климаксом, который в данном смысле называется моноклимаксом. Это еще один аргумент в пользу биополевого механизма координации жизни в биосфере - каждый регион планеты развивается в соответствии с характерной для него программой и имеет свою характерную структуру, призванную обеспечить определенную функцию, закрепленную за данным регионом в едином организме биосферы (или планеты).
Правда, справедливости ради надо сказать, что у автора концепции моноклимакса, американского эколога Клементса, имеется достаточное количество оппонентов. Может быть, это связано с тем, что любая крайне полярная точка зрения всегда оказывается односторонней. Концепция моноклимакса хорошо укладывается в холистический подход, однако сторонники исследования детальных механизмов природных явлений приводят множество примеров, когда в районах с известным моноклимаксом формируются совершенно другие климаксные экосистемы. Например, в южной тайге наряду с еловыми лесами могут формироваться луговые и сосновые климаксные экосистемы. Сторонники моноклимакса склонны рассматривать их как промежуточные стадии, которые рано или поздно обязательно должны прийти к моноклимаксу, если для этого не будет таких препятствий, как, например, вмешательство человека. Так луговые экосистемы, похоже, существуют только за счет регулярного скашивания молодой древесной поросли. Сосновые леса могут возникать и без участия человека, особенно на крайне бедных почвах, типа песчаных и щебнистых почв, куда более требовательная к почвам ель внедриться не может. Однако по мере накопления плодородия почв, ель, как более сильный эдификатор, скорее всего, вытеснит сосну и создаст моноклимаксную экосистему.
Проверить это достаточно сложно, так как подобного рода сукцессии растянуты на тысячи лет. Иногда экологам улыбается удача и они обнаруживают экосистемы, где можно непосредственно “наблюдать” такие тысячелетние сукцессии. Это позволяет выявить некоторые характерные особенности сукцессионного процесса. Так например, замечательной природной лабораторией являются дюны у южной оконечности озера Мичиган (США, штат Индиана). Озеро постепенно отступает, оставляя после себя все более молодые песчаные дюны, абсолютно безжизненные, которые постепенно “осваиваются” жизнью. Процесс этот длится уже тысячи лет, поэтому разноудаленные от кромки воды дюны находятся на разных стадиях сукцессии, что позволяет проследить весь ход сукцессионного процесса. Первые поселенцы на дюнах - это прибрежные злаки, ива, песчаная вишня, хлопковое дерево и животные: пауки, кузнечики и т.п. За ними следуют сухие сосновые леса, затем сухие леса из черного дуба, и, наконец, на самых старых дюнах - влажные леса из дуба, бука и клена, для которых характерна мощная богатая гумусом почва, населенная дождевыми червями и улитками. Для достижения климаксного сообщества на дюнах требуется около 1000 лет, что примерно в пять раз больше, чем это требуется при более лучших, чем дюны, исходных условиях.
В зависимости от исходных условий принято различать первичные сукцессии, которые начинаются на абсолютно безжизненных субстратах, например, на дюнах, и вторичные сукцессии, начинающиеся с более благоприятных стартовых условий, например, после пожара, вырубки леса или на заброшенном поле. Вторичные сукцессии могут начинаться с более поздних пионерных видов, например, с трав или кустарников. Так например, сукцессии на заброшенных полях начинаются с однолетних трав, бывших ранее сорняками. Затем они сменяются многолетними травами, кустарниками и деревьями. Ввиду большего исходного плодородия почвы такие сукцессии достигают климакса гораздо быстрее, чем в случаях первичной сукцессии. В последнее время для биосферы характерны именно вторичные сукцессии, что связано главным образом с деятельностью человека.
Вмешательство человека в природу ведет зачастую также к явлениям дигрессии (от латинского слова дигрессион – отклонение), когда климакс экосистемы (устойчивое равновесное состояние) достигается на более ранних стадиях сукцессии, вследствие чего экосистемы значительно упрощаются. В отличие от моноклимакса эта стадия называется катаценозом (от греческого слова ката – вниз, против). Типичным примером является пастбищная дигрессия, когда вместо леса, на данном пространстве формируется обширная луговая экосистема. Чем дальше отступает пояс лесов, тем реже достигают данного пространства семена кустарников и деревьев. Регулярный выпас скота не дает развиться молодым деревьям. Постепенно на этом месте вдали от лесов формируется особая зона, богатая гумусом, где никогда (точнее, очень долго) не вырастет лес, даже если человек оставит этот район. Так, вероятно, формировались степи, на которых древние кочевые племена пасли свои табуны и стада. Попытки возобновить леса в степной зоне требуют немалых усилий, так как помимо колонии деревьев, лес характеризуется особой структурой симбиотических связей, особенно между конями деревьев и грибницей (микроза). Поэтому лес может постепенно “наступать” на степь. Но, чтобы лес самопроизвольно возник посреди степи - случай практически невозможный. Для того, чтобы лес заменил степи после ухода человека потребуются, вероятно, десятки тысяч лет.
Следует различать автотрофные и гетеротрофные сукцессии. Все описанные выше примеры относятся к автотрофным сукцессиям, так как они протекают в экосистемах, где центральным звеном является растительность (фитоценоз). Динамика гетеротрофов целиком подчинена динамике автотрофов - смена животных сообществ зависит от смены растительных сообществ. Автотрофные сукцессии могут длиться теоретически вечно, так как они постоянно подпитываются энергией Солнца.
В гетеротрофных сукцессиях участвуют только животные (гетеротрофы, консументы). В этот процесс могут быть также вовлечены и мертвые растения, например, поваленные деревья, пни и т.п., которые являются, как правило, источником энергии для гетеротрофной сукцессии. Примером такой сукцессии может служить разрушение поваленного дерева. Первыми на мертвом или ослабленном дереве поселяются насекомые-короеды, которые повреждают кору, давая возможность проникнуть в места повреждений спорам грибов. Грибы-пионеры размягчают древесину, окрашивая ее в разные цвета. Это облегчает появление следующей волны насекомых, которые питаются уже древесиной (ксилофаги). В поврежденной ими древесине развиваются грибы-деструкторы, способствующие появлению мягкой гнили. Затем их сменяют грибы-гумификаторы, превращающие гнилую древесину в гумус. На всех стадиях гетеротрофной сукцессии также идет соответствующая смена видов микроорганизмов. В конечном итоге все органическое вещество разлагается до конечных продуктов: минеральных веществ и углекислого газа. Аналогичные процессы идут и при разложении трупов животных, а также в экскрементах животных, в лесной подстилке, в загрязненных водах и т.д.
 |
| Рис.24 |
Гетеротрофная сукцессия предполагает обязательное наличие определенного запаса энергии, аккумулированной в органическом веществе. Она заканчивается вместе с исчерпанием ресурса энергии, то есть после полного разложения исходного субстрата. После этого экосистема перестает существовать. То есть для нее не определено понятие климакса. В отличие от биогеоценозов, такие экосистемы смертны.
Гетеротрофная сукцессия хорошо ассоциируется с обществом, зависящим от ископаемого горючего. Динамика гетеротрофной сукцессии описывается кривой с быстрым ростом числа организмов, пока не будет достигнут некоторый максимум, затем количество организмов постепенно уменьшается по мере исчерпания энергетического ресурса (рис.24). Какого-то устойчивого состояния (климакса) добиться не удается. Такое общество быстро прогрессирует, но тем не менее оно изначально обречено на вымирание. Мы уже “сняли сливки” с большинства месторождений. Дальнейшая их эксплуатация со временем будет требовать все больших вложений энергии. Поэтому эффективность добычи полезных ископаемых будет неуклонно падать. Вместе с этим будет падать и жизнеспособность цивилизации, построенной на гетеротрофной сукцессии, если, конечно, еще раньше не произойдет катастрофических изменений, например, массового психоза - людям свойственно искать виновников своих проблем. Поэтому мы огромное количество усилий направляем на поиск новых источников энергии. Но даже если мы научимся управлять термоядерным синтезом, это не изменит нашей деструктивной сущности. Мы только несколько отодвинем в будущее развязку, как это мы делали уже не раз, но стабилизировать гетеротрофную сукцессию нам не удастся. Это даже если не учитывать, что в случае успеха в области энергетики перегрев биосферы неизбежен.
12.2. Закономерности сукцессионного процесса
В любом сукцессионном процессе можно выявить ряд закономерностей:
1. Изменение энергетики.
На ранних стадиях автотрофной сукцессии скорость образования валовой первичной продукции (П) превышает скорость дыхания сообщества (Д) (расходы на жизнеобеспечение плюс потребление гетеротрофами), то есть П/Д > 1. В результате биомасса (Б) будет постоянно накапливаться. Поэтому по мере сукцессии и первичная продуктивность и расходы на дыхание сообщества растут, но при этом отношение П/Д стремится к балансу П/Д = 1, характерному для стадии климакса. На этой стадии вся первичная продукция расходуется на дыхание и потребление гетеротрофами. Это значит, что на стадии климакса чистая продукция экосистемы теоретически равна нулю, а количество биомассы максимально.
Гетеротрофная сукцессия, наоборот, начинается с отношения П/Д < 1, но также по мере развития сукцессии стремится к П/Д = 1. То есть в обоих случаях наблюдается тенденция к равновесию между связанной энергией и энергией, затрачиваемой на поддержание биомассы. Поэтому отношение П/Д = 1 может служить показателем зрелости системы.
2. Изменение разнообразия.
На ранних стадиях сукцессии видовое разнообразие незначительно, но по мере развития сукцессии, оно нарастает, меняется видовой состав сообщества, усложняются и удлиняются жизненные циклы, увеличивается величина организмов, развиваются взаимовыгодные симбиозы, усложняется трофическая структура экосистемы. Однако пик разнообразия приходится вовсе не на стадию климакса, а либо на середину сукцессионного ряда, либо ближе к климаксу.
На ранних стадиях сукцессии преобладают виды с высокой скоростью размножения и роста, но низкой способностью к индивидуальному выживанию. На стадии равновесия давление естественного отбора благоприятствует видам с низкой скоростью роста, но большей способностью к выживанию. Таким образом, в процессе сукцессии происходит, по-видимому, переход от количества продукции к качеству, сопровождаемый генетическими изменениями, охватывающими всю биоту.
3. Совершенствование круговоротов биогенных элементов.
По мере усложнения трофической структуры экосистемы увеличивается время оборота биогенных веществ. Кроме того, циклы многих биогенных элементов, таких как азот, фосфор и кальций, замыкаются в пределах данной экосистемы. Все это способствует накоплению запаса биогенных элементов. То есть в процессе сукцессии количество этих элементов на входе оказывается большим, чем их количество на выходе. В зрелой климаксной системе вход и выход биогенных элементов почти уравновешиваются, однако некоторое накопление все же происходит. Таким образом, равновесная система более независима от внешних поступлений вещества.
4. Относительность неизменности климаксных стадий.
После достижения климакса динамические процессы вовсе не прекращаются, но динамика экосистемы изменяет свой характер. Она становится более замедленной. Одни организмы сменяют другие, происходит внедрение в экосистему новых видов, формируются новые экологические ниши, совершенствуются адаптации, происходит образование новых видов, формируются новые симбиозы и т.д. Особенно явно эти процессы можно наблюдать в регионах с относительно стабильной средой. Возраст экосистем в этих районах может исчисляться миллионами лет, сложность их организации превышает сложность организации любого организма.
Однако очень большое влияние на динамику климаксных систем оказывают внешние воздействия, которые способны отбросить всю экосистему или отдельные ее районы на более ранние стадии сукцессии. Это могут быть пожары, сильные бури, наводнения, горные лавины и другие природные катаклизмы.
Если вы войдете в густой еловый лес, то увидите, что под пологом леса царит полумрак. В нижнем ярусе леса многие молодые ели погибают раньше, чем успевают пробиться в верхний ярус. Нижние ветви старых деревьев также давно посохли. Многие старые деревья ослаблены болезнями, много деревьев, поваленных сильным ветром. Если в какое-то из деревьев ударит молния, то пожар в таком лесу неминуем. Обычно в лесном массиве области, достигшие стадии климакса чередуются с более молодыми областями (мозаичный климакс), которые более устойчивы к пожарам, опасными для них разве что только в наиболее засушливые годы. Пожар, как правило, выжигает именно области, достигшие климакса, где преобладают старые и сухие деревья. В образовавшиеся прогалы вторгаются виды, свойственные ранним стадиям сукцессии. Одновременно здесь развивается поросль из находящихся в подстилке семян. То же самое можно сказать и о действии сильных ураганов.
В результате действия подобных периодических возмущений лесной массив находится в постоянной динамике, которую можно назвать циклической сукцессией, или циклическим климаксом, или мозаичным климаксом. Взгляд сверху на такой лес позволяет выявить области, находящиеся на разных стадиях сукцессии, которые со временем постепенно развиваются по направлению к климаксу, а затем попеременно возвращаются в одно из промежуточных состояний. В таком лесном массиве в любой момент времени обитает множество самых разнообразных видов растений и животных, характерных для разных стадий сукцессии.
В настоящее время роль наиболее сильного катастрофического фактора, способствующего омоложению экосистем выполняет человек, регулярно вырубающий часть леса на свои хозяйственные нужды. Лесники обычно следят за ходом вырубки, определяя участки, наиболее удачные с точки зрения наименьшего ущерба для леса. В некотором смысле это даже играет положительную роль. Правда, в отличие от естественных факторов, например пожара, с территории леса удаляется солидная часть биомассы, а значит, биогенные элементы безвозвратно изымаются из экосистемы. Поэтому вторичная сукцессия начинается с более ранних этапов и лес вынужден заново наращивать плодородие почвы. Однако темпы сукцессии не укладываются в ритм современной цивилизации. Лес еще не успевает достигнуть стадии зрелости, но уже приходят лесорубы и требуют от него “дань”. В результате почвы постепенно истощаются. Это в лучшем случае. В худшем случае лес сводится полностью и больше уже не восстанавливается, превращаясь в поля, луга, болота, или пустыни.
Интересное влияние на ход сукцессии оказывает постоянный приток биогенных элементов в экосистему, неважно естественным или антропогенным путем. Это может быть промышленный сброс в озеро биогенных элементов, или же естественный приток вещества, принесенного реками с водосборного бассейна, в экосистемах суши это может быть внесение удобрений и т.п. В сукцессионном отношении такая экосистема отбрасывается назад к более молодому состоянию с наиболее интенсивной динамикой. В агроценозах, например, это позволяет искусственно поддерживать господство однолетних и двулетних трав. Однако если приток биогенных веществ превышает влияние на сукцессию внутренних процессов, то экосистема не только не в состоянии как-то стабилизироваться, но и “угасает”, то есть заполняется органическим веществом, которое вместо положительного фактора становится загрязнителем. Например в озере, в которое сбрасываются промышленные органические отходы, происходит активизация и рост количества микроорганизмов, потребляющих органику, при этом растут их затраты на дыхание, то есть потребление О2. Если это потребление существенно превысит поступление О2 в процессе фотосинтеза, то в озере возникает дефицит кислорода, а потом вообще формируются анаэробные условия, в которых гибнет сначала рыба, а потом все более мелкие живые организмы, вплоть до тех микроорганизмов, по чьей “вине” это произошло. В результате продолжающееся поступление в озеро промышленных отходов постепенно превращает его в болото, где могут существовать только анаэробные бактерии.
Подводя итог разговору о закономерностях скцессионного процесса, необходимо ответить на вопрос: в чем причина сукцессий, почему одни виды в определенной последовательности замещают другие, почему климаксные экосистемы в данной местности так похожи друг на друга, кто управляет всем этим?
В качестве ответа на этот вопрос можно привести две теории:
1) виды ранних сукцессионных стадий меняют условия существования и подготавливают почву для последующих поселенцев;
2) первые виды противостоят вторжению других видов (ингибируют их) и сохраняются до тех пор, пока не будут вытесненными в результате конкуренции, выедания гетеротрофами, самоотравления продуктами своей жизнедеятельности и т.п.
Каждая из моделей имеет свои достоинства и недостатки. Так первая модель хорошо объясняет первичные сукцессии, но плохо работает в случае вторичных сукцессий. Например, почему бы после пожара не развиться сразу же молодой поросли ели. Опыт показывает, что подобные эксперименты выращивания климаксных лесов, минуя предыдущие стадии сукцессии, обречены на неудачу. Попытки объяснить это тем, что молодые ели требуют затенения для нормального роста, не выдерживает критики, так как в городских условиях, например, ель хорошо развивается и на открытом месте. Вторая модель также имеет свои слабые места. Например, внедрение ели под полог лиственного леса происходит как раз в тот момент, когда почвенные условия наиболее благоприятны для лиственных деревьев.
С точки зрения холизма, сукцессия является частью процесса развития экосистемы как единого целого. Например Клементс, развивая теорию моноклимакса, считал, что сообщество в своем развитии повторяет последовательные стадии развития экосистемы, как своеобразного организма (биогенетический закон). По сути дела, это генетическая теория сукцессии. Сейчас эта теория сильно модифицирована, но не утратила своей актуальности.
Какими бы ни были конкретные механизмы сукцессии, в любом случае этот процесс подчиняется второму закону термодинамики, который в данном случае по предложению Ю. Голдсмита читается как второй закон экодинамики: любая экосистема стремится к климаксу, как состоянию с наибольшим значением энтропии (кстати, первый закон экодинамики Ю.Голдсмита по аналогии с первым законом термодинамики называется законом сохранения структуры биосферы). Действительно, именно климакс, а точнее, даже мозаичный климакс, отвечает условию наличия в системе наибольшей неопределенности, накопленной в видовом разнообразии, в сложности и запутанности трофической структуры, в огромном количестве взаимоотношений организмов друг с другом и с факторами среды и т.п. При прочих равных условиях более сложные экосистемы успешно конкурируют с более простыми и замещают их. Они также оказываются более независимыми от внешней среды как в отношении потоков энергии, так и в отношении поступлений вещества. Это говорит о том, что такие экосистемы имеют меньше противоречий с окружающей средой, то есть с надсистемой, что говорит о их гармоничности.
Конечно реализация второго закона термодинамики в данном смысле имеет свои особые механизмы, которые пытаются вскрыть сторонники редукционизма и выдать их за причины сукцессий. Однако ни одна такая теория не способна охватить всего многообразия явлений сукцессии. Истина, вероятно, как всегда лежит в дополнительности редукционистского и холистского подходов. Таков один из самых главных законов природы.
12.3. Стабильность и устойчивость экосистем
Важнейшими показателями динамики экосистем являются устойчивость и стабильность. Определение устойчивости экосистем уже давалось - это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия. Под стабильностью экосистемы понимают ее способность сохранять свою структуру и функциональные свойства при воздействии на нее внешних факторов. Иногда понятия устойчивость и стабильность рассматриваются как синонимы, но тогда следует различать два вида устойчивости: резидентная устойчивость (стабильность) - способность оставаться в устойчивом (равновесном) состоянии под нагрузкой, и упругая устойчивость (собственно устойчивость) - способность быстро восстанавливаться при снятии нагрузки. В разных терминах имеются различные смысловые оттенки, которые нужно учитывать.
Системы с высокой резидентной устойчивостью способны воспринимать значительные воздействия, не изменяя существенно своей структуры, то есть практически не выходя за пределы равновесного состояния. Поэтому понятие упругой устойчивости для них не определено (если система не выходила за пределы равновесия, то как можно говорить о возвращении в равновесное состояние после снятия возмущения). Если внешнее воздействие превышает определенные критические значения, то такая система обычно разрушается. В технике подобное качество называется жесткостью. Предельные значения внешних воздействий, которые система способна выдержать без разрушения соответствуют запасу жесткости. Когда говорят о высокой резидентной устойчивости, то имеется в виду именно высокий запас жесткости данной системы. Это несколько отличается от понятия высокой стабильности, так как здесь в первую очередь внимание обращается на неизменность структуры. Тундра, например, обладает высокой стабильностью, но она очень ранима, у нее малый запас жесткости, то есть малая резидентная устойчивость. Экосистему тундры очень легко разрушить. Достаточно проехать вездеходу. Колеи, которые он оставляют за собой, сохраняются десятилетиями. Такие экосистемы по аналогии с техникой можно назвать хрупкими.
Системы с малой резидентной устойчивостью для нормального существования должны обладать высокой упругой устойчивостью. Они более чувствительны к внешним возмущениям, под действием которых они как бы “прогибаются”, частично деформируя свою структуру, но после снятия или ослабления внешних воздействий быстро возвращаются в исходное равновесное состояние. При превышении пороговых воздействий такая система теряет устойчивость, то есть все дальше удаляется от состояния равновесия. Диапазон воздействий, которые может выдержать система без разрушения, в технике соответствует запасу упругости. Таким образом, степень упругой устойчивости можно оценить как упругостью, определяющей степень сопротивления внешнему воздействию и скорость возврата в исходное состояние после снятия воздействия, так и запасом упругости. В отличие от упругих систем, пластичные системы после снятия внешнего воздействия не возвращаются в исходное состояние, а приходят к какому-то другому равновесному состоянию. Так согласно точке зрения оппонентов теории моноклимакса, для экосистем характерно не одно, а несколько состояний равновесия (климакса). Таким образом, для пластичных экосистем характерна малая упругая и малая резидентная устойчивость.
Похоже на то, что резидентная и упругая устойчивости взаимоисключают друг друга, точнее, экосистеме трудно развивать оба вида устойчивости. Например, одни леса состоят из деревьев с толстой корой, обладающих повышенной резидентной устойчивостью к пожарам. Но если такой лес все-таки сгорит, то его восстановление, как правило, крайне проблематично. Напротив, многие леса очень часто горят (низкая резидентная устойчивость), но быстро восстанавливаются (высокая упругая устойчивость). Ориентация экосистем на один из видов устойчивости определяется, как правило, изменчивостью среды: при стабильных условиях экосистемы склонны к более высокой резидентной устойчивости, при изменчивых условиях предпочтение отдается упругой устойчивости.
 |
| Рис. 25 |
Природа обоих видов устойчивости состоит в наличии в экосистеме разного рода обратных связей. В основном это отрицательные обратные связи, которые направлены на стабилизацию параметров экосистемы, возвращая их значения к какой-то изначально заданной величине. Однако немаловажную роль играют и положительные обратные связи, усиливающие благоприятные для системы изменения, например, в плане роста и выживаемости организмов. Однако деятельность положительных обратных связей обязательно должна быть ограничена соответствующими отрицательными обратными связями, иначе ничем не контролируемая экспансия жизни может привести экосистему к гибели. В обоих случаях схема управления выглядит одинаково (рис.25).
Механизмы реализации обратных связей называются механизмами гомеостаза (от греческих слов homoios - подобный, одинаковый и ststis - состояние). Обычно они достаточно сложны, так как их компоненты связаны в единые информационные сети факторами среды, а также различными физическими и химическими агентами - “посредниками”, подобно тому, как нервная или гормональная системы связывают в одно целое части организма. Примером может служить сильная реакция организмов в экосистеме на слабые концентрации некоторых веществ. С энергетической точки зрения можно выделить одну закономерность, о которой уже говорилось ранее: обладатели высококачественной энергии, оставаясь в меньшинстве, управляют большими потоками энергии более низкого качества. Например, хищники управляют численностью травоядных животных. В то же время активность хищников и их численность управляется численностью их жертв по цепи обратной связи. По этой цепи на более высококачественный энергетический уровень (к хищникам) подается небольшая часть низкокачественной энергии - хищники уничтожают не всех жертв, а лишь небольшой их процент. Причем влияние этой части энергии на управление всей системой существенно усиливается деятельностью хищников.
Помимо отрицательной обратной связи устойчивость экосистемы может быть обеспечена избыточностью компонентов. Например, если в системе имеется несколько автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры. То же самое можно сказать и про гетеротрофов. Поэтому видовое разнообразие способствует повышению устойчивости экосистем (закон Эшби). При этом как в случае резидентной, так и в случае упругой устойчивости расширяется диапазон пороговых воздействий, которые способны выдержать экосистема без разрушения (запас жесткости и запас упругости).
Так, например, для тундры с ее бедным разнообразием характерны такие проявления низкой упругой устойчивости, как периодические резкие увеличения численности мелких грызунов - леммингов. О малой резидентной устойчивости тундры мы уже говорили. Экосистема тундры очень ранима и очень долго восстанавливается. В то же время тропические леса с их богатым разнообразием способны достаточно быстро залечивать свои раны. Легенды повествуют о целых городах, поглощенных джунглями, население которых не могло справиться с их натиском и вынуждено было уходить в другие районы.
Если разные стадии сукцессии экосистемы считать квазистатическими состояниями, то здесь можно отметить ряд закономерностей. На первых стадиях сукцессии экосистемы способны очень быстро возобновляться. Например, мы можем распахать по весне все поле , но уже к осени оно будет укрыто травами. Это говорит о достаточно высокой упругой устойчивости на начальных стадиях сукцессии, которая по мере движения к климаксу постоянно снижается. То есть на каждой следующей стадии для восстановления исходного состояния требуется гораздо больше времени. Климаксный лес характеризуется очень малой упругой устойчивостью.
Для резидентной устойчивости тенденция обратная. На первых стадиях сукцессии резидентная устойчивость остается достаточно невысокой - системы очень чувствительны к внешним воздействиям, откликаясь на них повышенным динамизмом. Например, заросли кустарника или молодой березняк может быть легко поврежден вездеходом (малая резидентная устойчивость), однако вторичная сукцессия достаточно быстро “затянет” повреждение (высокая упругая устойчивость). По мере роста видового разнообразия резидентная устойчивость непрерывно растет. При этом растет одновременно и жесткость системы (стабильность) и запас жесткости (взрослый лес вездеход так просто не одолеет). Однако по мере приближения к климаксу, как уже было показано, экосистема становится более чувствительна к действию некоторых катастрофических факторов среды (бури, пожары и т.п.). То есть на фоне продолжающегося роста стабильности (жесткости), запас жесткости экосистемы при движении ее к климаксу, по-видимому, начинает уменьшаться, что можно объяснить преобладанием в ней старых деревьев. Это говорит о том, что она становится все более хрупкой.
Как уже было сказано, для реальных лесов характерен, как правило, мозаичный климакс. По-видимому, именно такой лес имеет наилучший комплекс показателей устойчивости и стабильности. Периодические сильные воздействия на него в целом лес выдерживает достаточно жестко, жертвуя лишь наиболее старыми и хрупкими участками, которые, даже если они превращаются после пожара в абсолютно пустые пространства, в условиях непосредственной близости неповрежденного леса восстанавливаются сравнительно быстро.
Примером абсолютно нежизнеспособных экосистем являются агроценозы, создаваемы человеком из однолетних и двулетних культурных растений, то есть продукты сельскохозяйственной деятельности человека. Многие экологи даже не считают их экосистемами, хотя есть все основания считать их экосистемами с искусственно поддерживаемыми начальными стадиями сукцессии. Заброшенные поля сразу же втягиваются в естественный ход сукцессии. Что касается возделываемых полей, то они существуют только за счет человека. Свидетельством их нежизнеспособности является крайне малая устойчивость как упругая, так и резидентная, что является следствием очень скудного видового разнообразия как флоры (все сорняки подавляются гербицидами), так и фауны (“вредных” насекомых мы травим пестицидами). Малая упругая устойчивость проявляется, например, в резких вспышках численности конкретных видов вредителей. Подобные вспышки в естественной природе наблюдаются очень редко. Малая резидентная устойчивость проявляется, например, в повышенной чувствительности посевов к природным условиям, таким как град, засуха, повышенное количество осадков и т.д.
Осваивая все большие территории планеты, человек продолжает стратегию обеднения видового разнообразия. Этим он все более подрывает способность природы сопротивляться внешним воздействиям и возвращаться в исходное состояние. Как знать, может быть роковая черта, за которой начинаются необратимые изменения, уже пройдена. Это значит, что биосфера никогда больше не придет к исходному состоянию. Это значит, что мы вступили на путь глобальных перемен, которые постепенно изменят жизнь на Земле самым коренным образом. Будет ли в этом мире место человеку?