
Классические законы г. Менделя 42
| Вид материала | Закон |
- Лекции тема 7, 852.45kb.
- Законы делимости (дискретности) в мире животных и растений. Законы наследственности, 276.87kb.
- Н. брумберг, В. Попов, 78.9kb.
- Решение задач по генетике с использованием законов Г. Менделя, 419.2kb.
- Лекция 18. Генетика. Первый и второй законы Г. Менделя, 108.91kb.
- Темы уирс: Этапы развития медицинской генетики. Наследственно обусловленные патологические, 69.44kb.
- Применение flash – анимаций на уроках биологии, 68.93kb.
- Направление: Искусство и гуманитарные науки, 1316.91kb.
- Основные причины и условия жестокого поведения Введение, 1346.89kb.
- Лекция Классические маркеры I типа, 237.04kb.
Г л а в а IV
ДНК КАК ОСНОВА НАСЛЕДСТВЕННОСТИ
Для психогенетики, главным объектом исследования которой является природа индивидуальных различий, ознакомление со структурой и механизмами функционирования ДНК важно для понимания того, как гены влияют на человеческое поведение. Гены само поведение не кодируют. Они определяют последовательности аминокислот в белках, которые направляют и создают основу химических процессов клетки. Между геном и поведением лежат многочисленные биохимические события, открытие и понимание которых — интереснейшая задача, решаемая разными науками. Вариативность гена, тот факт, что он существует во множественных формах (аллелях), создает основу для формирования индивидуальных различий — соматических, физиологических, психологических. Именно в этом смысле говорят, что ДНК и есть материальная основа наследственности: вариативность генетическая создает, в контексте средовой вариативности, вариативность фенотипическую.
1. НУКЛЕИНОВЫЕ КИСЛОТЫ
Нуклеиновые кислоты являются непериодическими полимерами. Различают два вида нуклеиновых кислот: дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК). ДНК содержится главным образом в составе хромосом клеточного ядра; РНК находится и в ядре, и в цитоплазме.
ДНК
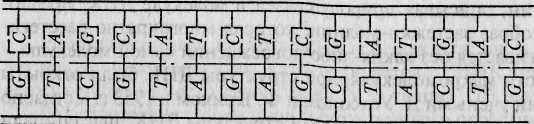
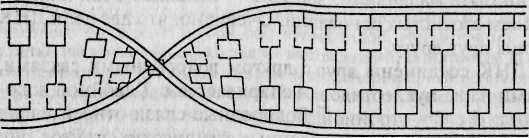
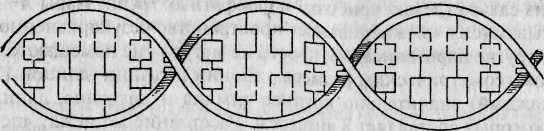
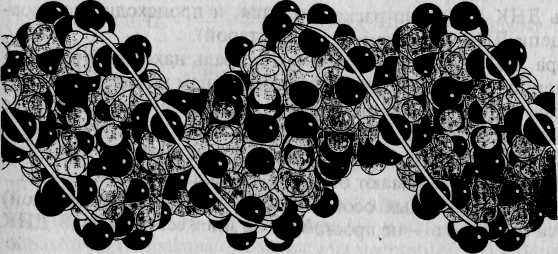
Во всех живых организмах (за исключением некоторых вирусов) наследственная информация передается из поколения в поколение с помощью молекул ДНК. Каждая клетка организма человека содержит примерно 2 метра ДНК. Обычно ДНК состоит из двух комплементарных цепей, формирующих двойную правостороннюю спираль (рис. 4.1а, б). Напомним, что каждая цепь представляет собой линейный полинуклеотид, состоящий из четырех азотистых оснований: аденина (А), гуанина (G), тимина (7) и цитозина (С)*. При формировании двойной спирали ДНК >4 одной цепи всегда спаривается с Гдругой, а G — с С. Эти основания называются комплементарными. Принцип селективности этой связи чрезвычайно прост и определяется принци-
*
 Азотистые основания в соединении с сахаром и фосфатом называются нук-леотидами.
Азотистые основания в соединении с сахаром и фосфатом называются нук-леотидами.Конец страницы №92
Начало страницы №93
Конец страницы №93
Начало страницы №94
пом наличия свободного пространства. Дело в том, что спиральная лестница ДНК зажата с двух сторон в ограничивающие ее «перила», состоящие из сахара (дезоксирибозы) и фосфатных групп. Пары А— Т и G— С вписываются в «межперильное» пространство безукоризненно, а вот любые другие пары вписаться просто не могут — не помещаются. Так, по своим геометрическим размерам аденин и гуанин (длиной 12 ангстрем* каждый) значительно крупнее тимина и цитозина, длина каждого из которых составляет 8 ангстрем. Расстояние же между «перилами» всюду одинаково и равно 20 ангстремам. Так что пары А—Ти G—C неслучайны: их структура определяется как размером (одно основание должно быть маленьким, а другое — большим), так и химическим строением азотистых оснований. Очевидно, что две цепи ДНК комплементарны друг другу.
Две цепи ДНК соединены друг с другом водородными связями, объединяющими пары нуклеотидов. А спаривается с Гдвойной водородной связью, a G с С — тройной. Водородные связи относительно непрочны; под воздействием определенных химических агентов они легко как разрушаются, так и восстанавливаются. Американский генетик Р. Левонтин, описывая природу связей в молекуле ДНК, предложил удачный образ застежки-молнии, которая многократно расстегивается и застегивается без каких-либо повреждений самой молекулы.
Особенности макромолекулярной структуры ДНК были открыты американскими учеными Д. Уотсоном и Ф. Криком в 1953 г. Согласно разработанной ими трехмерной модели структуры ДНК, шаг спирали ДНК составляет примерно 34 ангстрема, а каждый ее виток содержит 10 нуклеотидов, расположенных на расстоянии 18 ангстремов друг от Друга.
ДНК обладает свойством ковариантной редупликации, т.е. ее молекулы способны копировать сами себя с сохранением возникших в них изменений. Это удвоение происходит в ходе процессов, которые называются митозом и мейозом (см. гл. I). В процессе удвоения (репликации) ДНК, который осуществляется с участием ферментов, двойная спираль ДНК временно раскручивается, и происходит построение новой цепи ДНК (комплементарной старой).
Структура ДНК динамична: двойная спираль находится в постоянном движении. Самые быстрые из известных нам процессов, разворачивающихся в ДНК, связаны с деформацией связей в каждой из ее цепей; эти процессы занимают пикосекунды (10~12 с). Разрушение и создание связей между комплементарными основаниями — процессы более медленные; они занимают от тысячной доли секунды до часа.
Одной из поразительных особенностей ядерной (хромосомной) ДНК является то, что она — не простой набор множества генов. В ДНК
* Ангстрем — десятимиллионная доля миллиметра.
Конец страницы №94
Начало страницы №95
высших организмов много последовательностей, которые ничего не кодируют. В организме человека эти последовательности составляют примерно 80—90% всей ядерной ДНК, так что кодирующие последовательности — скорее исключение, чем правило. Некодирующие последовательности ДНК служат удобным источником так называемых полиморфных маркёров.
Малые ДНК. В клетках ядро является не единственным «местом» в организме, где можно найти ДНК. Митохондрии — органеллы, находящиеся не в ядре клетки, как хромосомы, а в цитоплазме, тоже имеют собственную ДНК, но в целом митохондриальной ДНК в клетке значительно меньше, чем ядерной.
В разных организмах количество митохондриальной ДНК не одинаково, оно изменяется от организма к организму. Например, митохондриальная ДНК человека содержит 16 569 пар нуклеотидов. Несмотря на то что в каждой клетке имеется несколько тысяч митохондрий, а организм высших животных построен из миллиардов клеток, митохондриальный геном каждой конкретной особи, по-видимому, одинаков во всех ее митохондриях. Это позволяет использовать митохондриальную ДНК в популяционных и эволюционных генетических исследованиях.
РНК
Рибонуклеиновая кислота (РНК) содержится как в одно-, так и в двуцепочечных молекулах. РНК отличается от ДНК тем, что она содержит рибозу вместо дезоксирибозы и урацил (Ј/) вместо тимина.
В соответствии с функцией и структурными особенностями различают несколько видов молекул РНК, два из которых — матричная, или информационная, РНК (мРНК, или иРНК) и транспортная РНК (тРНК). Матричная РНК принимает участие в транскрипции гена, транспортная — в его трансляции. Правильная «сборка» последовательности аминокислот в белковую цепь происходит с помощью рибосом — специальных частиц в цитоплазме клеток; они содержат третью форму РНК — рибосомную РНК (рис. 4.2).
. транскрипция трансляция „
ДНК — ► мРНК — ► Белок
Рис. 4.2. Этапы синтеза белка.
ТРАНСКРИПЦИЯ
При самокопировании ДНК каждая из ее цепочек играет роль образца для создания дополнительной к ней цепочки. Подобным же образом при образовании молекулы матричной РНК одна из цепочек ДНК служит образцом для построения дополнительной к ней цепочки, но уже не из нуклеотидов ДНК, а из нуклеотидов РНК. Иными
Конец страницы №95
Начало страницы №96
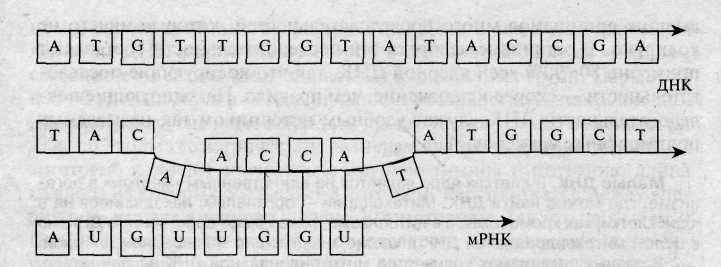
Рис. 4.3. Транскрипция — копирование ДНК молекулой матричной РНК.
словами, последовательность нуклеотидов строящейся цепочки РНК определяется последовательностью нуклеотидов молекулы ДНК.
Синтез молекулы мРНК по матрице ДНК называется процессом транскрипции.
Молекула мРНК имеет одноцепочечную структуру. Механизм ее образования подобен тому, который используется при репликации (самокопировании) ДНК (рис. 4.3). После окончания транскрипции мРНК покидает ядро и выходит в цитоплазму клетки, перемещаясь к рибосомам — «фабрикам» по производству белков. На рибосомах и осуществляется синтез белков.
ТРАНСЛЯЦИЯ
Молекула белка представляет собой цепочку аминокислот. Аминокислотами называются органические (карбоновые) кислоты, содержащие, как правило, одну или две аминогруппы (—NH2) и кислотную группу СООН (отсюда и название — аминокислоты). Друг от друга аминокислоты отличаются химической группой, называемой радикалом (R).
В табл. 4.1 перечислены двадцать аминокислот, являющихся основным «строительным материалом» при создании белков. (Главные аминокислоты распадаются на три класса в зависимости от природы группы R: нейтральные (полярные и неполярные), т.е. не несущие заряда в растворе; основные, положительно заряженные в растворе; и кислотные, отрицательно заряженные в растворе.)
Белки синтезируются с помощью ферментов путем соединения аминокислот так называемой пептидной связью: СООЯ-группа одной молекулы аминокислоты присоединяется к УУ#2-группе другой (при этом выделяется молекула воды). Данный процесс чрезвычайно сложен, но его скорость удивительна — аминокислоты строятся в цепоч-
Конец страницы №96
Начало страницы №97
Таблица 4.1
Названия аминокислот и их краткое обозначение
| Нейтральные ' | Основные (+) | Кислотные (-) | |
| Неполярные | Полярные | ||
| Триптофан (Тгр) Фенилаланин (Phe) Глицин (Gly) Алании (Ala) Валин (Val) Изолейцин (Не) Лейцин (Leu) Метионин (Met) Пролин (Pro) | Тирозин (Туг) Серии (Ser) Треонин (Thr) Аспарагин (Asn) Глутамин (Gin) Цистеин (Cys) | Лизин (Lys) Аргинин (Arg) Гистидин (His) | Глутаминовая кислота (Glu) Аспарагиновая кислота (Asp) |
ки полипептидов (белков) со скоростью примерно 100 аминокислот в секунду. В среднем белки содержат 100—1000 аминокислот, и от того, какова последовательность аминокислот в этих длинных цепях, зависят структура и функция данного белка. Любая аминокислота одинаково хорошо соединяется с любой другой (в том числе и с такой же); при этом взаимодействуют между собой одинаковые у всех аминокислот группы атомов NH2 и СООН. Благодаря этой способности аминокислот могут образовываться длиннейшие цепи.
Как же осуществляется синтез белков? Оказывается, что для описания строения конкретного белка достаточно указать последовательность аминокислот: какая из них занимает первое место, какая — второе, третье и т.д. Например, строение белка инсулина таково:
аланинлизин — пролин — ... — лейцин — аланин ...
Последовательность нуклеотидов в ДНК, а затем и в мРНК определяет, какой должна быть последовательность аминокислот, т.е. каким будет строение данного белка. Одна цепь ДНК содержит информацию о химическом строении значительного числа различных белков. Таким образом, последовательность оснований мРНК кодирует последовательность аминокислот. Сведения о строении белков — это «зерно» информации, передаваемой потомкам из поколения в поколение; кодирование аминокислот нуклеотидами и называется кодированием наследственной информации.
Всего существует 64 возможных тройки нуклеотидов (43= 64), кодирующих 20 аминокислот. Некоторые из нуклеотидных комбинаций играют роль «дорожных знаков», регулирующих синтез белка (напри-
Конец страницы №97
Начало страницы №98
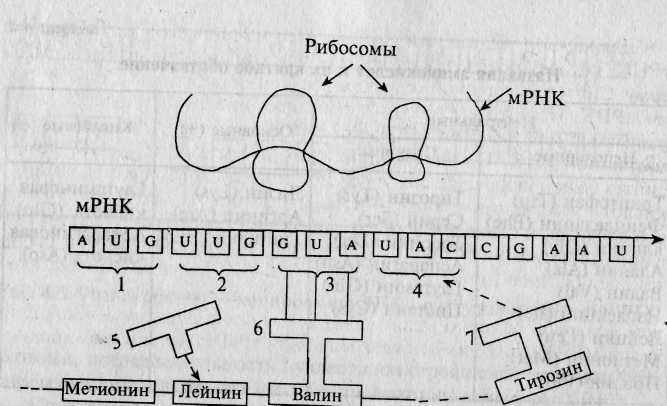
Р
 ис. 4.4. Трансляция мРНК (синтез белка).
ис. 4.4. Трансляция мРНК (синтез белка).1-4 — кодоны: метионина, лейцина, валина, тирозина; тРНК: 5 — отделилась от аминокислоты (лейцина), присоединив ее к белковой цепи; 6 — молекула тРНК, несущая валин, готова присоединить его к растущей белковой цепи; 7 — молекула тРНК, несущая тирозин, подходит к мРНК, определяя кодон тирозина.
мер, кодируя стоп-сигнал — сигнал окончания транскрибированной последовательности). При этом излишние комбинации нуклеотидов могут либо вообще не использоваться при кодировании наследственной информации, либо служить дополнительными (синонимическими) способами записи тех же самых аминокислот.
«Сборка» молекулы белка из аминокислот обеспечивается весьма сложным механизмом, главным образом в рибосомах — особых орга-неллах клетки, находящихся в цитоплазме. Рибосомы примерно наполовину состоят из рибонуклеиновой кислоты (отсюда и их название).
Напомним, что в процессе транскрипции формируется мРНК, которая комплементарна определенному участку ДНК. В ходе трансляции нуклеотидная последовательность мРНК выступает как основа, матрица для синтеза белка. «Считывание» последовательности мРНК происходит группами по 3 нуклеотида. Каждая аминокислота соответствует определенному сочетанию трех оснований — так называемому триплету (отсюда — триплетный код), или кодону.
«Сырье» (аминокислоты), необходимое для синтеза белка, находится в цитоплазме. Доставка аминокислот к рибосомам (рис. 4.4) производится с помощью сравнительно небольших специальных молекул транспортной РНК (тРНК). Небольшими эти молекулы, состоящие примерно из сотни нуклеотидов, можно считать только по сравнению с матричной РНК, состоящей из тысяч нуклеотидов.
Конец страницы №98
Начало страницы №99
Для каждой из двадцати аминокислот имеется свой тип молекулы тРНК, которая обеспечивает доставку данной аминокислоты в рибосому. Синтез белка происходит при движении рибосомы вдоль цепочки мРНК. При этом молекулы тРНК, несущие аминокислоты, выстраиваются, согласно коду молекул мРНК, в цепочку, параллельную матричной РНК. На рис. 4.4 показано, что молекула мРНК начала синтез белка, включающего, в частности, последовательность аминокислот ...«метионин—лейцин—валин—тирозин»... Валин был только что добавлен к белковой цепочке, к которой перед этим были присоединены метионин и лейцин. Кодон мРНК, представляющий собой триплет GUA, соединяется с молекулой тРНК, несущей аминокислоту валин. Молекула тРНК доставляет эту аминокислоту к концу растущей белковой цепочки и присоединяет валин к лейцину. Следующий кодон мРНК, UAC, привлекает молекулу тРНК, несущую-аминокислоту тирозин.
Процессы транскрипции и трансляции можно описать, использовав метафору французского ученого проф. К. Элена. На «фабрике» (в клетке) чертежи хранятся в «библиотеке» (в ядре), а для «выпуска продукции» (белков) используются не сами «чертежи» (ДНК), а их «фотокопии» (мРНК). «Копировальная машина» (РНК — полимера-за) выпускает или по одной «страничке фотокопии» (ген), или сразу целую «главу» (набор генов с близкими функциями). Изготовленные «копии» выдаются через специальные «окошки» (поры ядерной мембраны). Затем их используют на «монтажных линиях» (рибосомы) с «дешифратором» (генетический код) для получения из «заготовок» (аминокислот) окончательной «продукции» (белка), i /
2.БИОХИМИЧЕСКИЙ КОД НАСЛЕДСТВЕННОСТИ
РАЗНООБРАЗИЕ БЕЛКОВ
Белки выполняют в организме самые различные функции. В качестве ферментов они служат катализаторами химических реакций; в роли гормонов они, наряду с нервной системой, управляют работой различных органов, передавая химические сигналы. Белки используются в организме и как строительный материал (например, в мышечной ткани), и как транспортные средства (гемоглобин крови переносит кислород).
Размах синтеза белка, происходящего в клетке, огромен. Геном человека (набор последовательностей ДНК, определяющих генетическую индивидуальность человека) содержит порядка 6 биллионов нуклеотидов, из которых сформировано примерно 100 000 генов, чьи размеры варьируют в пределах от 1000 до 2 миллионов нуклеотидных пар. Если бы мы захотели описать эти 6 биллионов азотистых оснований и предположили, что на одной странице можно уместить около
Конец страницы №99
Начало страницы №100
3000 нуклеотидов, то нам понадобилось бы примерно 2 000 000 страниц — «многотомное собрание» нуклеотидов (и это для генома только одного человека)!
Описание всех генов человека и расшифровка соответствующих последовательностей ДНК — основная задача международного исследовательского проекта «Геном Человека», который является самым крупным генетическим проектом в мире. Благодаря усилиям многих генетических лабораторий мира ученые будут иметь в своем распоряжении полное описание генома человека.
ТИПЫ И СТРУКТУРА ГЕНОВ
До конца 80-х — начала 90-х годов геном называли сегмент ДНК, кодирующий полипептидную цепочку или определяющий функциональную молекулу РНК. Однако современные молекулярные исследования коренным образом изменили наше представление о структуре гена. Сегодня понятием «ген» обозначается сегмент геномной ДНК или РНК, выполняющий определенную функцию (причем выполнение этой функции вовсе не означает, что ген должен быть транскрибирован и транслирован).
В настоящее время разделяют три типа генов: гены, кодирующие белки, которые транскрибируются в РНК и затем транслируются в белки; гены, кодирующие РНК; и регуляторные гены, которые содержат нетранскрибируемые последовательности. Гены, кодирующие белки и РНК, называются структурными генами; их активность, «включение» и «выключение» определяются генами-регуляторами.
По мере проникновения в молекулярную структуру генетического материала все труднее становится находить в молекулах ДНК границы того, что обозначается понятием «ген». Это связано с тем, что процессы транскрипции (на ДНК) и трансляции (на мРНК) прямо не совпадают ни по локализации, ни по составу нуклеотидов. Наконец, постоянно увеличивается число открываемых генетических единиц. Так, наряду со структурными и регуляторными генами обнаружены, например, участки повторяющихся нуклеотидных последовательностей, функции которых мы только начинаем понимать, и мигрирующие нуклеотидные последовательности (мобильные гены).
Структура гена сложна, и в данном учебнике она подробно рассматриваться не будет. Отметим только наиболее важные моменты. В основном гены высших организмов имеют прерывистую структуру. Обычно они состоят из блоков (экзонов) — транслируемых участков, которые копируются в мРНК, переносимую в цитоплазму, и других блоков (интронов) — нетранслируемых участков, которых в мРНК нет. На начальном этапе транскрипции ген копируется полностью в пре-мРНК вместе с нитронами, которые затем «вырезаются», образуя зрелую мРНК. Так, некодирующая ДНК присутствует даже внутри самих генов.
Конец страницы №100
Начало страницы №101
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНОВ
В каждый конкретный момент клетка не использует всю содержащуюся в ее хромосомах генетическую информацию. Например, клетки печени вырабатывают специфические ферменты, которые не синтезируются, скажем, клетками почек, хотя те и другие содержат в своих ядрах одну и ту же ДНК. Кроме того, гены включаются и выключаются на разных стадиях онтогенеза: например, организм человека производит разные типы белка гемоглобина на разных этапах созревания организма (ранний эмбриогенез, развитие плода, детство, взрослый возраст). Синтез этих белков контролируется разными генами, которые включаются и выключаются на разных этапах онтогенеза.
Регуляция генной экспрессии осуществляется на нескольких уровнях при помощи целого набора клеточных механизмов. Общая задача процесса регуляции — избежать напрасных затрат энергии и создать условия для того, чтобы клетка производила наиболее эффективным образом все, в чем она нуждается. Процесс регуляции разворачивается в соответствии с заданной генетической программой или в ответ на изменения как во внутренней, так и во внешней среде организма. Считается, что в геноме человека количество регуляторных генов примерно соответствует количеству структурных генов.
3. ИЗМЕНЧИВОСТЬ НА УРОВНЕ ДНК
До сих пор мы преимущественно говорили об изменчивости генотипа в его широком определении. В последних разделах этой главы речь пойдет об изменчивости на уровне ДНК.
МУТАЦИИ ДНК
В главах I и III были даны определение мутаций и их классификации. Здесь мы рассмотрим только один из видов мутаций — так называемые точковые мутации, т.е. мутации, вовлекающие отдельно взятые нуклеотиды. Точковые мутации представляют собой вставки или выпадения, а также изменения (разные типы замен одного азотистого основания на другое) пары нуклеотидов ДНК (или нуклеотида РНК). В результате мутирования возникают альтернативные формы генов (аллели) —ген становится полиморфным. Одни из этих мутаций являются вредоносными, т.е. вызывающими развитие наследуемых заболеваний (главы II, III), а другие — нейтральными, не вызывающими никаких существенных изменений в синтезируемых белках.
Точковые мутации можно разделить на два больших класса. К первому классу относятся те, которые связаны с заменой основания. Мутации второго класса обусловлены так называемым сдвигом рамки считывания.
Конец страницы №101
Начало страницы №102
Замена одного основания в цепи ДНК может привести к тому, что в синтезируемый белок будет встроена «неправильная» аминокислота (пример такой трансформации: Мама мыла раму ==> Мама рыла раму). В результате функция белка может быть нарушена. Например, если первый кодон мРНК (рис. 4.4) скопирован неправильно и вместо A UG в последовательности мРНК записана последовательность AGG, то вместо метионина будет синтезирован аргинин. Подобная замена единственной аминокислоты в цепочке сотен аминокислот, составляющих белок, может проявиться по-разному. Спектр этих проявлений — от нулевых до летальных — зависит от структуры и функции синтезируемого белка.
ТИП МУТАЦИОННОГО СОБЫТИЯ: СДВИГ РАМКИ СЧИТЫВАНИЯ
Мутации, которые приводят к выпадению или вставке одного и более нуклеотидов, вызывают так называемый сдвиг рамки считывания. В среднем они более вредоносны, чем мутации замены нуклеоти-да. Примеры подобных трансформаций: Мама мыла раму => Ммам ылар аму — выпадение нуклеотида; Мама мыла раму => Мама мыла драму — вставка основания. Сдвигом рамки этот тип мутаций называется потому, что в результате выпадения (или случайного добавления) одного нуклеотида изменяется считывание (трансляция) кодо-нов в молекуле мРНК и, начиная с точки, соответствующей положению мутации, синтезируется искаженная последовательность аминокислот. Например, если в результате мутации теряется второй нуклеотид в последовательности ТАС-ААС-САТ, то эта цепочка счи-тывается как ТСА-АСС-АТ. В результате произведенный белок будет содержать не метионин (ТАС) и лейцин (AAQ, а серии (ТСА) и триптофан (ACQ, что приведет к нарушениям последующих биохимических процессов.
Часто мутации оказываются гораздо сложнее описанных выше. Один и тот же ген может мутировать в нескольких местах. Например, известно более 60 разных мутаций одного гена ФКУ, каждая из которых приводит к развитию фенилкетонурии (гл. II), причем некоторые из этих мутаций соответствуют разным степеням тяжести заболевания. Мутации, происходящие в экзонах (кодирующих участках гена), как правило, вредоносны. К счастью, большинство мутаций в организме происходит в интронах (некодирующих участках гена). Эти мутации не транскрибируются мРНК и, следовательно, фенотипически не проявляются.
Замечательная особенность мутаций состоит в том, что их действие может быть различным в разных организмах и фенотипические проявления одной и той же мутации у разных особей могут быть очень разнообразными. Так, обладание мутантным аллелем у одной особи
Конец страницы №102
Начало страницы №103
может фенотипически проявиться в форме тяжелого заболевания, а у другой — в форме легкой симптоматики или даже полного ее отсутствия. Два ключевых понятия, описывающих изменчивость проявления одной и той же мутации в популяции как совокупности организмов (подробнее о генетике популяций в гл. V), — упоминавшиеся (гл. II) понятия пенетрантности и экспрессивности.
Пенвтрантностъю называется частота проявления аллеля определенного гена у особей данной популяции. Различают пенетрантность полную (аллель проявляется у всех особей) и неполную (аллель не проявляется у части особей). Количественно ее выражают в процентах особей, у которых данный аллель фенотипически проявляется (100% — полная пенетрантность).
Приведем пример. Известно, что не все носители мутации гена ФКУ страдают фенилкетонурией. Пенетрантность мутантного аллеля ФКУ высока и составляет примерно 99%. Это означает, что среди каждых 100 носителей аллеля-мутанта в среднем будет один носитель, не имеющий фенотипичес-ких признаков заболевания — среди 100 мутировавших генов один ген-мутант не проявится, т.е. не вызовет развитие заболевания.
Экспрессивностью называется степень фенотипической выраженности одного и того же аллеля определенного гена у разных особей. Если фенотипический признак, контролируемый данным аллелем, в популяции не варьирует, то говорят о постоянной экспрессивности, в противоположном случае — об изменчивой (вариабельной) экспрессивности.
Различия экспрессивности означают, во-первых, разную степень пораженности носителей мутации (например, больные ФКУ — носители одной и той же мутации — могут страдать умственной отсталостью разной степени), а во-вторых, разные формы фенотипического проявления одной и той же мутации (например, предполагается, что один и тот же ген-мутант вызывает один тип психического расстройства — синдром Туретта — у мужчин и другой тип — синдром навязчивых идей — у женщин).
Новые мутации — важнейший источник генетической изменчивости, являющейся основой биологической эволюции. Частота мутирования отдельного гена чрезвычайно мала, но генов в организме много, а каждый биологический вид представлен множеством особей. Так что, когда организм или биологический вид рассматривается как целое, мутация выглядит не как редкое, а как вполне регулярное событие. Предположим, что геном человека насчитывает 100 000 пар генов, а средняя частота мутации на один ген составляет 10~5. Тогда среднее число мутаций в одном поколении составит (2-105 генов) х х (10~5 мутаций на ген) = 2 мутации на зиготу человека. На Земле живет около 4-Ю9 людей. Если у каждого человека возникает по 2 мутации, то общее число новых мутаций у ныне живущего населения земного шара составляет 8-109.
Конец страницы №103
Начало страницы №104
Мутации — основной источник генетического полиморфизма, т.е. наличия в популяции нескольких аллелей одного локуса. Полиморфная природа ДНК позволила разработать системы методов генетического и психогенетического анализа, которые позволяют определить и картировать целый ряд генов, вовлеченных в формирование индивидуальных различий по исследуемым поведенческим признакам. Так, например, использование полиморфных маркёров ДНК позволило картировать ген на коротком плече хромосомы 4, ответственный за развитие хореи Гентингтона.
В качестве примера рассмотрим два типа ДНК маркёров: полиморфизм длины рестрикционных фрагментов (/fFZP-полиморфизм) и полиморфизм повторяющихся комбинаций нуклеотидов (STR-no-лиморфизм). Для изучения полиморфности (этот процесс также называется тайпингом ДНК) ДНК выделяется из клеток крови или любых других клеток организма, содержащих ДНК (например, берется со-скоб с внутренней стороны щеки). При использовании технологии RFLP, ДНК, под воздействием ферментов, распознающих специфические последовательности нуклеотидов в ДНК и избирательно разрушающих ее цепь в определенных местах, разрезается на куски-фрагменты. Такие ферменты впервые были найдены в бактериях, которые производят их с целью защиты от вирусной инфекции.
Существуют сотни таких «рестрицирующих» ферментов, каждый из которых разрезает ДНК в определенном месте, распознавая определенную последовательность оснований; этот процесс называется рестрикцией. Например, один из часто используемых ферментов, EcoRI, распознает последовательность GAA ТТС и разрезает молекулу ДНК между основаниями GhA. Последовательность GAA ТТС может быть представлена в геноме несколько тысяч раз. Если в определенном локусе эта последовательность различна у разных людей, то у тех из них, которые являются носителями измененной последовательности, фермент в данном локусе ее не разрежет. В результате ДНК геномов, несущих нестандартные последовательности, разрезана в данном локусе не будет и, следовательно, образует более длинный фрагмент. Таким способом распознается разница в структуре ДНК. В результате разреза «рестрицирующими» ферментами могут получиться два типа фрагментов, соответствующих данному локусу, — длинный и короткий. Их также называют аллелями. По аналогии с «обычными» генами полиморфизмы могут быть гомозиготными по короткому фрагменту, гомозиготными по длинному фрагменту или гетерозиготными по длинному и короткому фрагментам.
Несмотря на то что существуют сотни «рестрицирующих» ферментов, распознающих различные последовательности ДНК, они, как выяснилось, способны отыскать только примерно 20% полиморфных
Конец страницы №104
Начало страницы №105
участков ДНК. Были разработаны несколько других типов ДНК-маркёров, распознающих полиморфизмы других типов. Широко используется, например, полиморфизм повторяющихся комбинаций нуклеотидов (5ТЛ-полиморфизм). Как уже упоминалось, по неизвестной пока причине в ДНК присутствуют повторяющиеся последовательности, состоящие из 2, 3 или более нуклеотидов. Количество таких повторов варьирует от генотипа к генотипу, и в этом смысле они также обнаруживают полиморфизм. Например, один генотип может быть носителем двух аллелей, содержащих по 5 повторов, другой — носителем двух аллелей, содержащих по 7 повторов. Предполагается, что геном человека содержит примерно 50 000 локусов, включающих подобные повторяющиеся последовательности. Хромосомные координаты многих локусов, обнаруживающих 5ТЛ-полиморфизм, установлены и теперь используются для картирования структурных генов, служа координатами на хромосомных картах.
Таким образом, генетический полиморфизм, связанный с присутствием так называемых нейтральных (не изменяющих синтезируемый белок) мутаций, плодотворно используется в молекулярно-гене-тических, в том числе психогенетических, исследованиях, поскольку генетическую изменчивость, выявленную молекулярными методами, можно сопоставлять с изменчивостью фенотипов. Пока этот перспективный путь используется в подавляющем большинстве случаев для исследования разных форм патологии, дающих четко очерченные фенотипы. Однако есть все основания надеяться, что он будет включен и в изучение изменчивости нормальных психических функций.
* * *
Одним из наиболее замечательных биологических открытий XX столетия стало определение структуры ДНК. Расшифровка генетического кода, открытие механизмов транскрипции, трансляции и некоторых других процессов на уровне ДНК являются фундаментом в строящемся здании психогенетики — науки, одна из задач которой состоит в раскрытии секретов соотношения генов и психики. Современные представления о структуре и функциях ДНК коренным образом изменили наши представления о структуре и функционировании генов. Сегодня гены определяются не как абстрактные «факторы наследственности», а как функциональные отрезки ДНК, контролирующие синтез белка и регулирующие активность других генов.
Одним из основных источников изменчивости являются генные мутации. Своими успехами современная молекулярная генетика обязана открытию и использованию закономерностей мутирования ДНК с целью обнаружения и картирования генетических маркёров. Именно они позволят психогенетике перейти от популяционных характеристик к индивидуальным.,
Конец страницы №105
Начало страницы №106