
Законы делимости (дискретности) в мире животных и растений. Законы наследственности Г. Менделя
| Вид материала | Закон |
- Лекции тема 7, 852.45kb.
- Исследование о влиянии эволюционной теории на учение о политическом развитии народов, 6089.9kb.
- Исследование о влиянии эволюционной теории на учение о политическом развитии народов, 10156.41kb.
- Лекция 18. Генетика. Первый и второй законы Г. Менделя, 108.91kb.
- Н. брумберг, В. Попов, 78.9kb.
- Классические законы г. Менделя, 24318.4kb.
- Программа дисциплины дпп. Р. 01 Мутационные изменения растений и животных цели и задачи, 112.28kb.
- Направление: Искусство и гуманитарные науки, 1316.91kb.
- Решение задач по генетике с использованием законов Г. Менделя, 419.2kb.
- Законы сохранения и принципы симметрии, 283.17kb.
ФГОУ ВПО Курганская государственная сельскохозяйственная академия имени Т.С.Мальцева.
Факультет: экономический.
Отделение: экономическое.
Специальность: экономика и управление на предприятии АПК.
Курсовая работа
Тема: Законы делимости (дискретности) в мире животных и растений. Законы наследственности Г. Менделя.
Выполнил: студент 3 группы 1 курса
экономического факультета
экономического отделения
Ионин А. В.
Проверил: ассистент кафедры
Содержание.
1. Предмет и методы генетики…………………………………………..1
2. Цитологические основы наследственности и развития…………..4
2.1. Клеточное строение организмов……………………………………4
2.2 Строение клетки………………………………………………………4
^ 2.3 Передача наследственной информации в процессе размножения
клеток и при оплодотворении…………………………………………..7
3. Законы наследственности Г. Менделя……………………………..13
4. Выводы…………………………………………………………………24
1. Предмет и методы генетики.
Генетика изучает два свойства живых организмов – наследственность и изменчивость. Под термином «наследственность» понимают свойство родительских особей передавать свои признаки и особенности развития следующему поколению. Под термином «изменчивость» понимают свойство, противоположное наследственности; оно заключается в способности организмов изменяться под действием наследственных и не наследственных факторов.
Основные задачи генетики следующие: изучение механизма изменение гена, репродукции генов и хромосом, действия генов и контроля или процессов образования различных признаков или свойств организма; разработка методов конструирования наследственной программы живых организмов, борьбы с наследственными болезнями, повышения продуктивности животных и урожайности растений.
Генетика служит теоретической основой для совершенствования пород сельскохозяйственных животных, определение потенциальной продуктивности, разработки методов генетической оценки популяций и отдельных особей по качеству потомства. Важное значение имеет генетика и для растениеводства. Знания законов наследования и изменчивости признаков позволяет интенсифицировать селекционный процесс по созданию сортов растений, устойчивых к неблагоприятным условиям произрастания, вредителям и болезням. В селекции растений успешно используют гибридизацию, мутагенез, полиплоидию. Широкие возможности для создания новых форм растений открывают генетическая инженерия, гибридизация соматических клеток, культура клеток и тканей. В последние годы для повышения урожайности применяют различные макро- и микроудобрения, ядохимикаты, гербициды. Многие из них накапливаются в растениях и, попадая в организм животного или человека, вызывают вредные мутационные изменения.
Преемственность между поколениями осуществляется путем полового, бесполого или вегетативного размножения. При половом размножении возникновение нового поколения происходит в результате слияния материнской и отцовской половых клеток, поэтому потомки несут признаки обеих родительских форм. Г. Мендель установил четкие закономерности независимого наследования признаков у гибридного потомства. Открытие явления сцепленного наследования не изменило представления о дискретном характере наследственности.
Признак – одно из главных понятий генетики. Понятием «признак», условно обозначают единицу морфологической, физиологической или биохимической дискретности организма. Каждой особи присущи видовые, породные, и индивидуальные свойства и признаки. Один организм отличается от другого в первую очередь внешними морфологическими признаками, например высоким или низким ростом; наличием или отсутствием рогов у крупного рогатого скота; белой, красной окраской венчика у цветковых растений и т. п.
Признаки условно подразделяют на качественные и количественные. Качественными называют морфологические или биохимические признаки (масть, форма рогов, ушей и др.). Количественными являются признаки не имеющие четкого выражения, их изучают путем измерения, подсчета (масса, длина шерсти, жирность молока и т. д.).
При половом размножении передача признаков от родительских особей потомкам идет через половые клетки, имеющие ядро и цитоплазму. В зависимости от того, ядру или цитоплазме принадлежит ведущая роль в передаче данного конкретного признака, различают ядерную или цитоплазматическую наследственность.
Практически наследование всех признаков определяется ядерной наследственностью, которая закодирована в молекулах ДНК, находящихся в хромосомах ядра клетки. Цитоплазматическая наследственность определяется генами, которые находятся в органоидах клетки – митохондриях, пластидах, кинетосомах, плазмидах. Еще в генетике выделяют переходный тип наследственности, который трудно определить однозначно, так как признак несет в себе черты истинной и ложной наследственности.
Основные объекты генетических исследований – молекулы нуклеиновых кислот – ДНК и РНК. Изучение нуклеиновых кислот вирусов, бактерий, грибов, клеток растений и животных позволяет установить закономерности действия генов в процессе жизнедеятельности клетки и организма.
Генетика имеет свои методы исследования. Одним из важнейших является гибридологический. Это система скрещиваний, с помощью которых устанавливается характер наследования каких-то признаков. Хромосомная теория наследственности является продолжением законов Г. Менделя.
Цитологический метод позволяет изучать строение клеток, органоиды, взаимодействие их при передаче наследственной информации.
Онтогенетический метод применяется для изучения явлений онтогенеза (индивидуального развития) в изменяющихся условиях внешней среды.
Статистический метод изучает статистические закономерности в изменчивости.
^ 2. Цитологические основы наследственности и развития.
2.1. Клеточное строение организмов.
Все населяющие Землю живые организмы состоят из клеток: одноклеточные из одной клетки, многоклеточные – из многих клеток, число которых может достигать нескольких миллионов и даже миллиардов. Из клеток построены все ткани и органы растений и животных.
Клеточная теория включает три главных положения.
Первое положение соотносит клетку с живой природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые формы не принимало, обеспечивается в конечном итоге только клеткой.
Второе положение указывает, что единственным способом возникновения новых клеток является деление предшествующих клеток.
Третье положение клеточной теории соотносит клетку с многоклеточными организмами, для которых характерен принцип целостности и системной организации.
Среди живых организмов встречаются два типа организации клеток: прокариоты (к ним относят бактерии и сине-зеленые водоросли) и эукариоты (к ним относят все одно- и многоклеточные организмы – т. е. растения, грибы и животных).
^ 2.2 Строение клетки.
Прокариотические клетки покрыты цитоплазматической мембраной, которая играет роль активного барьера между цитоплазмой клетки и внешней средой. Вместо клеточного ядра имеется его эквивалент, который называется нуклеоид, лишенный оболочки и состоящий из одной молекулы ДНК. Эти клетки могут содержать плазмиды, очень короткие двойные спирали ДНК, замкнутые в кольцо.
Бактерии размножаются путем простого деления. Деление начинается с мезосомы, после этого две половинки мезосомы расходятся, увлекая за собой ДНК, последняя также делится на две части, из которых в последствии образуются ядерные области двух дочерних клеток.
Клетка эукариот организована сложнее. Несмотря на большое разнообразие растительных и животных клеток, все они состоят из цитоплазмы и ядра, заключенного в оболочку. В состав цитоплазмы входят органоиды и включения. К органоидам относятся: эндоплазматическая сеть, митохондрии, комплекс Гольджи, рибосомы и пластиды. Из включений можно назвать капли жира, гранулы белков, витамины, пигменты, вакуоли.
Ядро занимает патую часть клетки. От цитоплазмы ядро отделяется мембраной, через поры которой через цитоплазму проникают все виды РНК. В растительных клетках имеются пластиды. Зеленые пластиды, содержащие хлорофилл называются хлоропластами. В них под действием солнечных лучей из углекислого газа и воды идет первичный синтез органических веществ. В отличии от животных, растительные клетки покрыты цилюлозной оболочкой, который защищает их и предает им прочность.
Основным компонентом ядра является хроматин. Он окрашивается основными красителями, что указывает на его кислотные свойства. В состав хроматина входит ДНК в комплексе с белком.
Мембраны цитоплазмы представляют собой тонкие пласты липопротеидной природы. Основными компонентами мембраны являются липиды (40%), белки (60%) и углеводы. Смеси разных липидов высокой концентрации дают бимолекулярный слой, или мембрану.
Плазматические мембраны ограничивают клетку снаружи и контактируют с внеклеточной средой.
^ Эндоплазматическая сеть была открыта в 1945 г. К. Портером. Он изучал фибропласты цыплят с помощью электронного микроскопа и обнаружил вакуоли, канал или канальцы. Вся эта сеть усыпана гранулами (рибосомами), которые присоединены к мембране своей большой субчастицей.
^ Аппарат Гольджи выводит вещества за пределы клетки. Он представлен сетчатыми образованиями, которые встречаются во всех животных клетках. Благодаря аппарату Гольджи идет синтез полисахаридов и осуществляется взаимодействие с белками. С их же помощью идет выведение готовых секретов за пределы клетки.
Лизосомы – это сферические частицы, окруженные липопротеиновой мембраной. Они содержат большое количество кислых гидролаз. В растительных клетках лизосомы участвуют в мобилизации путем гидролиза питательных веществ эндосперма, а также в растворении и выделении отмирающих структур во время дифференциации и деления клеток.
Пластиды имеются буквально во всех клетках зеленых растений. Их три разновидности: лейкопласты, хлоропласты и хромопласты. Хлоропласты состоят из хлорофилла, белков, липидов и некоторого количества РНК. В лейкопластах образуется крахмал, а в хромопластах накапливаются окрашенные в желтый, красный или оранжевый цвет биологически важные вещества – каротиноиды. Все пластиды размножаются путем деления.
^ 2.3 Передача наследственной информации в процессе размножения клеток и при оплодотворении.
Митоз представляет собой способ упорядоченного деления клетки, при котором каждая из двух дочерних клеток получает такое же число и те же типы хромосом, какие имела материнская клетка.
Промежуток времени между окончанием одного клеточного деления и окончанием последующего называют митотическим циклом, который подразделяют на митоз и интерфазу.
Интерфаза включает три периода. В первом периоде – пресинтетическом, идущем вслед за митозом, осуществляется синтез белков и РНК. Затем следует период синтеза ДНК, в течение которого количество ДНК в ядре клетки удваивается. В постсинтетическом периоде происходит синтез РНК и белков и накапливается энергия для следующего митоза. По окончании интерфазы наступает непосредственно митоз – непрямое деление ядра.
Митоз подразделяют на четыре стадии: профазу, метафазу, анафазу и телофазу.
В профазе происходит формирование хромосом, хроматиды укорачиваются и утолщаются в результате процесса внутренней спирализации. Именно здесь формируется веретено деления, а скорость спирализации достигает максимума – 11 – 27 тыс. оборотов в минуту. Это первая фаза подготовки ядра к делению, в которой сетчатая структура ядра постепенно превращается в хромосомы. В профазе растворяется оболочка ядра, образуется веретено деления, ядрышко исчезает.
В метафазе хромосомы располагаются по экватору, а цитоплазма имеет меньшую вязкость. В этой фазе можно идентифицировать хромосомы и проводить их подсчет. Нити веретена прикрепляются к центромере с обоих полюсов клетки. Таким образом, к каждой хромосоме прикрепляется по две нити ахроматинового веретена.
В анафазе хроматиды отходят к полюсам в результате разрыва центромеры пополам. Сначала отталкиваются центральные участки. Каждый член разъединяющейся пары отходит к донному из полюсов. Это приводит к увеличению числа хромосом.
Телофаза начинается с момента достижения хромосомами полюсов. Хромосомы достигают такого состояния, когда видны лишь хромотиновые нити или гранулы. Вокруг каждого дочернего ядра образуется ядерная оболочка. На этом завершается деление ядра, за которым следует деление тела клетки.
При делении клеток животных на поверхности клетки в плоскости экватора образуется борозда. Она постепенно углубляется и разделяет цитоплазму на две дочерние клетки, в каждой из которых имеется ядро. У большинства типов клеток весь процесс митоза занимает 1 – 2 часа.
Регулярный и упорядоченный митотический процесс обеспечивает передачу генетической информацией каждому из дочерних ядер; в результате каждая клетка содержит генетическую информацию обо всех признаках организма.
В основе полового размножения лежат процессы образования исходных половых клеток. За этим процессом следует особый вид деления незрелых половых клеток- мейоз, а после него- не менее сложные биологические процессы формирования женского и мужского гаметофита у растений и животных.
В процессе размножения высших растений наблюдается смена полового и бесполого поколений. Обычно цикл развития растений состоит из двух периодов- диплоидного и гаплоидного. Диплоидный принято называть спорофитом, а гаплоидный- гаметофитом. Диплоидный период в онтогенезе характеризуется 2n числом хромосом и длится от начала образования зиготы до мейоза. Гаплоидный включает сам мейоз, гаметогенез и заканчивается периодом, предшествующим слиянию гамет.
У растений в мейоз вступают материнские клетки макроспор и мегаспор. Первые образуются из мужского генеративного органа –пыльника, состоящего из четырех лопастей, в которых закладываются микроспорангии. Они делятся и образуют клетки археспория, которые превращаются в материнские клетки микроспор.
Материнские клетки мегаспор развиваются из женского генеративного органа- пестика. Пестик состоит из рыльца, столбика и завязи. В завязи образуются семяпочки, а в одном из слоев нуцеллуса закладывается археспориальная клетка семяпочки, которая митотически делится и превращается в материнскую клетку мегаспор.
У животных из нескольких зародышевых клеток образуются половые клетки- гонии. Первичные половые клетки сначала одинаковы для мужских и женских особей, а позднее происходит половая дифференциация, в результате которой формируются первичные оогонии и первичные сперматогонии.
Таким образом, заканчивается период формирования незрелых половых клеток, предшествующий мейозу. По числу хромосом они не отличаются от соматических.
Следовательно в мейоз вступают незрелые половые клетки.
Мейоз в отличие от митоза состоит из двух делений. Первое деление называется редукционным, а второе, протекающее по типу митоза, эквационным. В каждом делении одинаковые фазы., однако после названия фазы в первом делении принято ставить римскую цифру I.
После названия фазы второго деления ставят римскую цифру II. Каждому делению предшествует интерфаза, однако перед вторым делением ее называют интеркинезом. Интеркинез отличается от интерфазы тем, что в нем не наблюдается удвоение ДНК.
В профазе наблюдается спирализация хромосом. Профаза разбита на пять стадий: лептотена, зиготена, пахитена, диплотена и диакинез.
Лептотена – это стадия тонких нитей. В этой стадии спирализация только начинается, но у хромосом уже видна двойственная структура, то есть они уже состоят из двух хроматид.
Зиготена – это вторая стадия профазы II. Здесь скорость спирализации начинает нарастать, хромосомы еще более «взрослеют» и в результате этого образуют гомологичные пары. Представители каждой пары сначала робко прикасаются, а затем сливаются по всей длине.
Пахитена – это стадия толстых нитей. Силы взаимного притяжения в результате конъюгации настолько велики, что гомологичная пара образует бивалент, в недрах которого закладываются точки будущего обмена (кроссинговера).
Диплотена – это стадия, в которой наблюдается явление, противоположное конъюгации. Вместо взаимного притяжения между хромосомами здесь наблюдается их отталкивание. Каждая из гомологичных хромосом бивалента расщепляется на две хроматиды, так что биваленты содержат по четыре хроматиды.
Диакинез – это последняя стадия профазы I. В этой стадии биваленты приобретают равновесное состояние, то есть силы отталкивания уравновешиваются силами сцепления между хромосомами.
Метафаза – I. Биваленты собираются в экваториальной пластинке, и к ним подходят две нити веретена деления, по одной от каждого полюса клетки. Каждая центромера присоединена к своей нити веретена деления .
Анафаза – I. Целые хромосомы расходятся к полюсам, что приводит к ложной редукции, поскольку наследственный материал остается удвоенным, несмотря на то, что число хромосом уменьшается вдвое.
Телофаза – I. У разных организмов образование телофазы I идет по-разному. У них идет образование клеток, то есть протекает цитокенез. Это тип, который характерен для ржи, пшеницы, кукурузы. У других организмов, как у гороха, подсолнечника, после телофазы цитокенез отсутствует, и клетки не образуются, а в конце мейоза все четыре микроспоры находятся в общей оболочке.
Интеркинез клетки проходит быстро без всяких изменений, после чего наступает второе деление мейоза, которое идет по типу митоза. Оно необходимо, чтобы довести концентрацию ДНК до нормы.
В метафазе – II в экваториальной пластинке располагаются хромосомы, число которых после первого деления уменьшено в два раза. К каждой хромосоме с разных полюсов клетки подходят нити веретена деления.
В анафазе – II центромеры разрываются пополам и отдельные хроматиды отходят к полюсам, а так как каждая монада несет двойную нагрузку ДНК, то незамедлительно превращается в целую хромосому. На этом мейоз заканчивается, и далее наступает гаметогенез.
Гаметогенез. Микроспоры растений одеты общей оболочкой, которая потом распадается на отдельные микроспоры с гаплоидным числом хромосом. Каждая микроспора превращается в пыльцевое зерно. Его первичное ядро делится, и в результате этого образуются две клетки, из которых одна вегетативная, а вторая генеративная. Генеративная клетка расположена в цитоплазме внутри вегетативной клетки. Последняя делится и образуется два спермия.
Из четырех мегаспор тетрады развивается только одна, а три остальные отмирают. Оставшаяся мегаспора формирует одноядерный зародышевый мешок. В результате митоза образуется двуядерная, затем четырехъядерная и восьмиядерная клетка, которую называют зародышевым мешком. Затем из восьмиядерной группы отделяется по одному ядру, которые продвигаются в центр зародышевого мешка, образуя центральное ядро с числом хромосом, равным диплоидному. Когда же произойдет оплодотворение, то центральное ядро зародышевого мешка становится триплоидным эндоспермом. Второй спермий сливается с яйцеклеткой, образуя диплоидный зародыш.
У животных оогонии и сперматогонии вступают в мейоз. После прохождения интерфазы образуются ооцит и сперматоцит первого порядка. В конце первого деления формируются ооцит сперматоцит второго порядка, а рядом образуется одно полярное тельце.
После второго деления из каждой клетки образуется по две гаплоидные клетки. При этом, у женской особи- одна ооцида и три направительных тельца, у мужской- четыре сперматиды. Из оотиды образуется яйцеклетка, а из сперматид – сперматозоиды.
Значение мейоза в жизни организмов заключается в следующем. При оплодотворении происходит слияние спермия с яйцеклеткой и геномы мужского и женского организма объединяются. Геном – это гаплоидный набор хромосом. Мейоз выступает в качестве механизма, регулирующего постоянство хромосом, сохраняет видовую специфичность кариотипа. В мейозе наблюдаются следующие важные явления: уменьшается число хромосом вдвое, диплоидное ядро превращается в гаплоидное, через кроссинговер реализуется наследственная изменчивость организмов.
^ 3. Законы наследственности Г. Менделя.
Чтобы установить, как передаются по наследству индивидуальные признаки у живых организмов, он начал экспериментировать с горохом и в 1866 г. опубликовал результаты экспериментов в «Трудах» общества естествоиспытателей г. Брно.
Г. Мендель впервые сформулировал идею о существовании наследственных факторов. Он создал также гибридологический метод, связанный с изучением характера наследования отдельных признаков и свойств организмов.
К основным особенностям гибридологического метода изучения наследственности относят:
1) Использование в качестве исходных форм для скрещивания растений, отличающихся друг от друга сравнительно небольшим числом (одна, две или три пары) контрастных признаков, и тщательный учет характера наследования каждого из них;
2) Точный количественный учет гибридных растений, различающихся по отдельным признакам, в ряде последовательных поколений;
3) Индивидуальный анализ потомства от каждого растения в ряде последовательных поколений;
4) Недопустимость влияния чужеродного генетического материала на родительские расы и гибриды;
5) Сохранение способности к размножению у гибридов и их потомков.
Мендель изучает наследственность дискретно, то есть по частям. Для своих опытов он подбирает 34 сорта гороха, но впоследствии оставляет только 7 пар, различающихся между собой четкими альтернативными признаками (желтые – зеленые, гладкие – морщинистые, красные – белые и т. д.). Пары подобраны, но Мендель упорно не начинает опытов еще целых три года, поскольку он хочет убедиться, что сорта отличаются только по одной паре признаков. Но вот сомнения развеяны, сорта оказались чистолинейными, опыты можно начать.
Мендель берет растения красноцветковые и скрещивает их с белоцветковыми. Осенью получает семена нулевого поколения.
Нулевым называется потому, что по семенам пока еще невозможно судить о цвете венчика у растений, которым предстоит вырасти из этих семян. Растения выросли с красными цветками.
При изучении формы горошины и цвета в соответствующих парах не нужно ждать следующего года, поскольку результат скрещивания виден уже к осени, когда горошины первого поколения созрели. Во всех семи парах гибриды первого поколения несли признаки одного из родителей. Следовательно, один признак оказался сильнее второго и Мендель его называет доминантным. Слабый признак назвали рецессивным. Но самое интересное заключалось в том, что во втором поколении наблюдалось расщепление по фенотипу 3:1.
Г. Мендель выдвигает следующие доводы, которые следует считать основными положениями:
- каждый признак определяется наследственным задатком, передающимся потомству через половые клетки;
- задатки не утрачивают своей индивидуальности в ряду поколений, то есть не смешиваются;
- оба пола в равной мере участвуют в передачи своих наследственных свойств потомству;
- в половых клетках число факторов уменьшается вдвое;
- факторы парные, а у гибридов один фактор от материнской, а другой от отцовской формы, один – доминантный, а другой – рецессивный.
Все эти положения легли в основу законов Менделя.
^ Первый закон – это закон единообразия гибридов первого поколения, то есть все гибриды первого поколения единообразны как по генотипу, так и по фенотипу.
Используем следующую символику:
АА – красные цветы, аа – белые цветы (Р:- родители); F – гибриды (
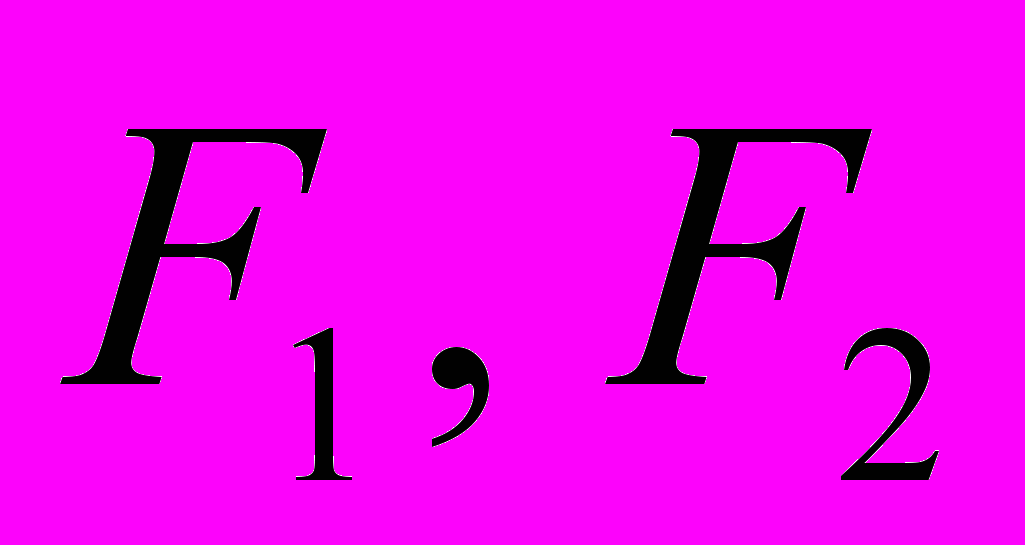 - гибриды первого и второго поколений);
- гибриды первого и второго поколений);  - возрастное скрещивание,
- возрастное скрещивание,  - анализирующее скрещивание;
- анализирующее скрещивание;  - смена первого поколения при отсутствии проявления признака.
- смена первого поколения при отсутствии проявления признака. Следовательно, скрещивание красноцветного гороха с белоцветным будет представлено следующим образом:
Р: АА х аа - 1-й год
Гаметы: А а
Аа
- в год скрещивания (признак не виден) - 1-й годАа
 - на следующий год (при цветении) - 2-й год
- на следующий год (при цветении) - 2-й год - на следующий год (при цветении) - 3-й год
- на следующий год (при цветении) - 3-й годАА: 2Аа: аа – Расщепление – по фенотипу 3:1
красные + красные: белые – по генотипу 1:2:1
В практической селекции первый закон Менделя играет важную роль при определении доминирования. Чтобы узнать, какой признак доминирует, необходимо скрестить формы с альтернативными признаками и если в
получают искомый признак, то он доминантен. Например, мы не знаем, как наследуется устойчивость к стеблевой ржавчине злаков. Для того, чтобы узнать, необходимо скрестить устойчивый сорт с восприимчивым, и если в первом поколении будут получены все восприимчивые растения, то признак устойчивости рецессивный.^ Второй закон Г. Менделя имеет следующую трактовку: - При самоопылении гибридов первого поколения получают расщепление по фенотипу 3:1 и по генотипу 1:2:1. Это закон расщепления.
^ Закон чистоты гамет. Для объяснения сущности закона достаточно рассмотреть неполное доминирование, когда скрещиваются растения львиного зева с красными и белыми цветками. При этом получают в первом поколении все розовоцветковые растения, то есть признак как бы разбавился, однако во втором поколении наблюдается расщепление по фенотипу и генотипу в соотношении 1:2:1 (1 часть красноцветковых: 2 части розовоцветковых: 1 часть белоцветковых). Таким образом можно убедиться, что гаметы не смешиваются и не растворяют друг друга, они остаются в чистоте.
Позднее стало известно, что гипотеза чистоты гамет предугадывает механизм мейоза. Было установлено, что гены одной пары признаков находятся в одинаковых локусах (участках) гомологичных хромосом. Такие гены получили название аллельных.
Несмотря на гениальность сделанных Г. Менделем открытий, это учение долгое время оставалось не понятым. И только через 35 лет, в 1900 году, трое ученых (Карл Корренс, Гуго де Фриз, Э. Чермак) одновременно и независимо друг от друга открыли менделеевские закономерности на различных культурах. Они, как и подобает ученым, начали научный поиск и все неожиданно натолкнулись на работу Г. Менделя, а вместо того, чтобы опубликовать результаты своих исследований, все трое опубликовали восторженные отзывы о трудах Менделя, продолжателями которых оказались. Этот год стал рождением генетики.
^ Третий закон Менделя – закон независимого наследования – был сформулирован им на основании изучения потомства от дигибридных скрещиваний. Дигибридным называют скрещивание особей, различающихся по двум парам альтернативных признаков. Так при скрещивании сорта гороха, имевшего желтые гладкие семена (ААВВ), с сортом, имевшим зеленые морщинистые семена (aabb), гибриды первого поколения имели желтые гладкие семена, так как гладкая форма доминирует над морщинистой. Во втором поколении среди 556 полученных семян Мендель обнаружил 315 желтых гладких, 101 желтых морщинистых, 108 зеленых гладких и 32 зеленых морщинистых, что модно выразить соотношением 9: 3: 3: 1. Во втором поколении произошла перекомбинация форм, появились семена с новыми сочетаниями признаков – зеленые гладкие и желтые морщинистые.
На основании анализа опыта Мендель пришел в заключению, что факторы А и В у гибридов АаВb при образовании половых клеток распределяются независимо друг от друга и дают различные сочетания с одинаковой частотой. Каждая половая клетка содержит только по одному представителю от пары факторов Аb и по одному представителю от пары Вb, поэтому возможны только четыре сочетания – АВ, Аb, аВ, аb.
Сочетания гамет с различными генотипами происходят случайно, поэтому образование зигот у гибридов первого поколения при дигибридных скрещиваниях можно выразить следующим образом: 9AB: 3Ab: 3aB: 1ab, которые полностью соответствуют найденному соотношению: 9 желтых гладких (AB): 3 желтых морщинистых (Ab): 3 зеленых гладких (aB): 1 зеленый морщинистый (ab).
Расщепление по генотипу во втором поколении представлено более сложно и выглядит так: AABB + Aabb + aaBB + aabb + 2AABb + 2aaBb + 2AaBB + 2Aabb + 4AaBb.
^ Наследование при полигибридном скрещивании.
Наследование признаков при скрещивании особей, различающихся по трем и парам альтернативных признаков, представлено сложнее.
AABBCC – гомозигота доминантная (тройной доминант),
aabbcc – гомозигота рецессивная (тройной рецессив),
AaBbCc – тригетерозигота,
AaBbCcDd – тетрагитерозигота.
Когда проводил свои опыты Мендель, то у него никогда не было полного соответствия между эмпирическими и теоретическими соотношениями генотипов и фенотипов при расщеплении. Это связано с объемом выборки, чем больше выборка, тем ближе это соотношение к теоретическому.
^ Виды доминирования.
Соотношения в расщеплении у гибридов второго поколения, установленные Г. Менделем, наблюдаются при полном доминировании, когда один аллель полностью подавляет действие другого. В последствии были обнаружены другие виды доминирования. Так, при скрещивании безухих овец с овцами имеющими нормальную длину ушей, все потомство первого поколения оказывается с короткими ушами. Такое явление было названо неполным доминированием. При скрещивании таких овец между собой во втором поколении получали овец с нормальным размером ушей, короткоухих и без ушей в соотношении 1: 2: 1. В дальнейшем при скрещивании овец с нормальными ушами все потомство имело нормальные уши, при скрещивании безухих овец все потомки были безухими, а при скрещивании короткоухих овец наблюдалось расщепление в соотношении 1: 2: 1.
^ Сверх доминирование – вид доминирования, при котором у гибридов первого поколения наблюдается более сильное развитие признака, чем у исходных родительских форм. Это явление используется в птицеводстве при создании различных кроссов и гибридов мясной птицы.
Позднее был установлен еще один вид доминирования – кодоминирование, при котором у гибридов первого поколения проявляются признаки того и другого предка, выраженные в равной степени и независимо друг от друга.
По типу кодоминирование наследуются группы крови животных, человека в системе АВО, различия в строении белков, например гемоглобина, ферментов и других.
^ Взаимодействие не аллельных генов.
Г. Мендель не ограничился установлением закона независимого наследования пар аллелей, он положил начало учению о взаимодействии генов, что накладывает заметный отпечаток на формы наследования многих свойств организмов. Взаимодействие между неаллельными генами проявляется в нескольких формах: комплементарности, эпистазе и полимерии.
Комплементарность. Это взаимодействие генов, при котором новый признак у особи развивается только в том случае, если в ее генотипе присутствуют два доминантных неаллельных гена в гомо- или гетерозиготном состоянии, например, AABB, AaBb, AaBB,AABb.
Различают три типа комплементарности. К первому типу относят такое взаимодействие, при котором новый признак обусловлен сочетанием доминантных генов их разных гомологичных пар хромосом, тогда как их генов в отдельности не имеет самостоятельного фенотипического проявления. Классический пример первого типа – это душистый горошек (Lathyrus odoratus). У этого растения пурпурная окраска цветков контролируется доминантными неаллельными генами А и В в гомо- или гетерозиготе. Отсутствие в зиготе одного из этих генов определяет белую окраску цветов. В данном случае объясним механизм дигибридного скрещивания.
Ко второму типу комплементарности относится взаимодействие, при котором каждый из доминантных генов в отличие от предыдущего может проявлять самостоятельность, что и определяет фенотип растения или животного. Например, у мышей черная окраска шерсти контролируется доминантным геном А, добавление гена В детерминирует окраску «агути» (верх и низ шерстинки черные, а середина ее желтая). Из этого следует, что черную масть будут иметь мыши с генотипом А – bb, агути – с генотипом А – B, белую – с генотипами aaB и aabb. Следовательно, сочетание двух доминантных генов A и В дает окраску «агути», наличие в зиготе только гена А в гомо- или гетерозиготе дает черную окраску шерсти, отсутствие доминантного гена А, даже в присутствии доминантного гена В или при его отсутствии, будет определять белую окраску шерсти мышей. Таким образом, во втором поколении наблюдается расщепление в соотношении 9: 3: 4.
К третьему типу комплементарности относится взаимодействие доминантных генов из разных пар хромосом, при котором каждый из них имеет самостоятельное фенотипическое проявление. Примером может служить наследование формы гребня у кур, окраска шерсти у норок, оперение у попугаев, цвета плодов у перца и т. д.
Рассмотри этот тип комплементарности на примере формы гребня у кур. У кур породы леггорн гребень имеет листовидную форму, у виандоттов – розовидную, у европейских пород – гороховидный и у кур малайского происхождения ореховидный.
Данное расщепление, в отличие от расщепления при дигибридном скрещивании, происходит по одному признаку, то есть каждая комбинация гибридов во втором поколении – это новый признак.
Ореховидная форма гребня может развиваться в процессе гибридизации особей с розовидными и гороховидными гребнями. Привлечение листовидной формы и ореховидной не дает должного результата.
^ Эпистатическое взаимодействие генов – это один из типов неаллельного взаимодействия, при котором действие доминантного гена одной пары подавляется геном другой. Эпистаз- явление, противоположное комплементарности. Различают доминантный эпистаз, который выражается неравенством А > В и рецессивный – aa > B, aa > bb. Ген, который подавляет, называется эпистатическим, а тот, что подавляется, - гипостатическим. Гены, подавляющие действие других генов, называются ингибиторами, или супрессорами. Характерным примером доминантного эпистаза может служить наследование оперения (окраски) у кур. При этом ген С контролирует наличие красящего пигмента, с – отсутствие его. Доминантный ген I (ингибитор) из другой пары хромосом подавляет действие гена С, а рецессивный аллель его не оказывает такого действия. Следовательно, в генотипах CCII, CCIi, CcII,CcIi присутствует ингибитор в доминантном состоянии, а поэтому куры будут белыми. Белыми они будут и в случаях сочетания генов ccIi и ccii. Таким образом во втором поколении можно наблюдать расщепление 13: 3.
Примером рецессивного эпистаза может служить наследование окраски шерсти у мышей, когда белая окраска развивается у особей с генотипом ааВВ и ааbb. Такой тип эпистаза называется криптомерией. По эпистазу наследуется масть лошадей. В этом случае серая масть определяется сочетанием доминантных генов А и В в разном состоянии, вороная – отсутствием гена А, а рыжая – отсутствием обоих доминантных генов. Характер расщепления при наследовании масти лошадей 12: 3: 1.
Наиболее распространенным примером эпистаза является окраска зерен овса посевного.
Изучая потомство гибридов, полученных от скрещивания белой и пурпурной фасоли, Мендель обнаружил, что во втором поколении получается целый ряд оттенков от пурпурного до бледно-лилового и белого; и при этом количество белых форм не равно ¼, как при моногибридном расщеплении, а значительно меньше. Мендель пришел к выводу, что развитие пурпурной окраски зависит от ряда генов, каждый из которых в отдельности дает промежуточные окраски. Такой тип взаимодействия генов, при котором на один признак действуют несколько пар неаллельных генов называют полимерией.
Неаллельные гены, действующие однозначно на образование одного признака, называются полимерными или множественными. При полимерном взаимодействии ферменты, образующиеся под контролем неаллельных генов, действуют на развитие одного и того же признака и добавление отдельного гена усиливает его проявление. Это кумулятивная полимерия. Когда степень выраженности признака не зависит от количества доминантных генов, и он развивается при наличии даже одного гена, такое взаимодействие называется некумулятивной полимерией. Примером кумулятивной полимерии может служить окраска зерна пшеницы. В этом случае темно-красное зерно определяется двойным доминантом, белое – двойным рецессивном.
У пшеницы растениеводы различают две окраски зерна – красную и белую. Если скрещивать сорт с красным зерном, например, разновидности Лютесценс с сортом разновидности Альбидиум (белое зерно), то во втором поколении отмечается расщепление 3: 1, то есть по обычной моногибридной схеме. Однако имеются отдельные сорта с темно-красным зерном. Например, разновидности Пиротрикс 28, при скрещивании которого с белозерным сортом, например, Грекум 114, расщепление идет в соотношении 15: 1.
По типу полимерии наследуются такие важные признаки, как высота растения, продолжительность вегетации, количество белка в зерне, яровость и озимость, содержание витаминов и др.
Яровость определяется доминантными полимерными генами, тогда как озимость – рецессивными. Скрещивание яровой и озимой пшеницы, в результате которого получим Курганский сорт Шадринская, дает гибриды с более продолжительным вегетационным периодом, но трансгрессивно увеличивается продуктивность.
Трансгрессивный характер проявления количественного признака заключается в том, что в результате скрещивания появляются формы с более сильным выражением признака, или этот признак ослаблен в сравнении с родителями. При скрещивании двух сортов яровой пшеницы со светло-красным зерном во втором поколении непременно появятся гибриды с темно-красным зерном. Появляются формы и с белым зерном. Следовательно, трансгрессии могут быть положительными и отрицательными. Если какой-то признак усиливается, то это положительная трансгрессия. При скрещивании озимых форм с яровыми увеличивается не только продуктивность, содержание белка, устойчивость к полеганию, но и адаптивность в местных условиях. В тоже время, иммунологические параметры Курганских сортов несколько снижены по сравнению с родителями, что говорит об отрицательной трансгрессии.
В случае с курганской селекцией видна недооценка такого важного признака, как продолжительность вегетации. Это произошло по тому, что в период селекции выбраковывали менее продуктивные колосья, что сочетается со скороспелостью.
4. Выводы.
К началу XX века в растениеводстве и животноводстве был накоплен экспериментальный материал о наследовании потомками признаков родительских форм. Особенно ценные данные были получены во второй половине XVIII века И. Кёльрейтером, который изучал полученные им гибриды у 54 видов растений и установил ряд закономерностей в наследовании признаков: равное влияние на признак отцовской и материнской форм, возврат признака у гибрида к одной из исходных родительских форм. Он впервые обратил внимание на дискретный характер наследования признаков, установил наличие пола у растений. Важное значение имели работы О. Сажре и Ш. Нодена, Т. Найта, А. Т. Болотова, К. Ф. Рулье, а так же многих других ученых и практиков, которые наблюдали и описывали характер наследования признаков у растений и животных.
Ч. Дарвин в своей работе «Происхождение видов» и в последующих трудах обобщил опыт и наблюдения практиков и естествоиспытателей по изучению явлений наследственности и изменчивости, которые на ряду с отбором являются движущими факторами эволюции органической природы.
Основоположником генетики принято считать Г. Менделя, который впервые разработал метод научного подхода к изучению наследственности и в своих опытах с растительными гибридами установил важнейшие законы наследования признаков, но его работы не были должным образом оценены современниками и не оказали существенного влияния на дальнейшее развитие генетики.
В 1900 году Г. де Фриз в Голландии, К. Корренс в Германии и Э. Чермак в Австрии независимо друг от друга установили, что полученные ими результаты по наследованию признаков у растительных гибридов полностью согласуются с данными Г. Менделя, который за 35 лет до них сформулировал правила наследственности, которые Г. де Фриз предложил называть законами наследования признаков.
В научные исследования тех лет активно включилась огромная армия микробиологов, биохимиков, генетиков и цитологов, которые осуществили научную революцию в биологии. Генная инженерия дала людям дрожжевых грибков. Линии культур клеток бактерий, продуцирующих биологически – активные вещества, белки для человека и животных.
Революционными можно назвать достижения американских ученых по созданию защищенного картофеля и сортов других трансгенных растений, устойчивых к вредителям, в частности к колорадскому жуку, фантастические открытия по клонированию животных.
Уже сейчас с помощью тонких генетических методов отбирают клоны клеток или вирусов, содержащих гибридные ДНК. Люди овладели методами расшифровки последовательностей нуклеотидов в молекуле ДНК, а в борьбе с болезнью века, СПИДом, уже созданы диагностические препараты, а ранняя диагностика это залог успешного лечения и раковых опухолей.
Наука развивается быстрыми темпами и за ней наше будущее.