
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Морфогенетические закономерности формирования продуктивности озимой ржи (Secale cereale L.)
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАПовышение урожайности у сортов типа Таловской 15 представляется более реальным потому, что как увеличение количества продуктивных побегов, так и массы зерна с колоса зависят от наиболее значимых положительных вкладов одних и тех же признаков. Кроме того, в увеличение количества продуктивных побегов наиболее существенный вклад вносит масса зерна с колоса, а в увеличение массы зерна с колоса далеко не последнюю роль играет увеличение количества продуктивных побегов. В связи с этим в условиях ЦЧР морфотип растений с доминантной короткостебельностью представляется нам более перспективным в селекции на повышение потенциала продуктивности.
Для получения данных о взаимосвязях элементов продуктивности и зависимости от них урожайности был проведен полный корреляционный анализ многолетних данных с использованием способа корреляционных плеяд (Терентьев П.В., 1956; 1975). Полученные результаты показали, что изученные нами признаки распадаются на три обособленные группы (плеяды). Содержание их у изученных сортов существенно не отличалось. Эти три группы были выделены нами и с использованием факторного анализа по способу главных компонент. Они учитывают 72% суммарной дисперсии урожайности.
По данным факторного анализа для первой компоненты наиболее значимыми являлись: продуктивная кустистость, количество зерен на растении и масса зерна с растения; для второй - масса зерна с колоса, количество зёрен в колосе, К хоз., масса зерновки и количество зёрен в колосе; для третьей - количество продуктивных побегов, масса надземной части растений, общее количество побегов и количество продуктивных растений.
Признаками-индикаторами (по Смирнову Е.С., 1924) являются: в первой компоненте - количество зёрен на растении, во второй - масса зерна с колоса и в третьей - количество продуктивных побегов на единице площади (табл. 3).
Таблица 3 - Средние значения коэффициентов корреляции между аосновными элементами продуктивности в компонентах
Номер компоненты |
Элемент продуктивности |
Среднее значение коэффициента корреляции с членами компоненты |
|
1 |
Количество зерен на растении |
0,84 |
Масса зерна с растения |
0,46 |
|
Продуктивная кустистость |
0,40 |
|
|
2 |
Масса зерна с колоса |
0,72 |
Количество зерен в колосе |
0,65 |
|
К хоз. |
0,57 |
|
Количество цветков в колосе |
0,52 |
|
Масса зерновки |
0,50 |
|
|
3 |
Количество продуктивных побегов |
0,72 |
Количество продуктивных растений |
0,64 |
|
Масса надземной части растений |
0,55 |
|
Общее количество побегов |
0,47 |
Учитывая перекрёстный характер опыления озимой ржи, в процессе работы с селектируемыми популяциями, когда основные браковки необходимо проводить до цветения, целесообразно использовать в качестве критериев продуктивности долю сохранившихся растений, продуктивную кустистость и количество цветков в колосе, которые коррелируют с указанными признаками-индикаторами. Последние необходимо использовать по данным структурного анализа.
Количество сохранившихся продуктивных растений и продуктивная кустистость относятся к разным плеядам и всегда тесно связаны между собою. В результате формируется один интегральный признак - количество продуктивных побегов на единице площади, который совместно с массой зерна с колоса формирует тот или иной уровень урожайности. С использованием множественного регрессионного анализа было установлено, что колебания урожайности на 73,3-85,5% зависят от величины этих признаков. Как видно из рисунка 4, они в сильной степени отрицательно коррелируют между собою, что весьма затрудняет одновременное увеличение значений этих признаков.
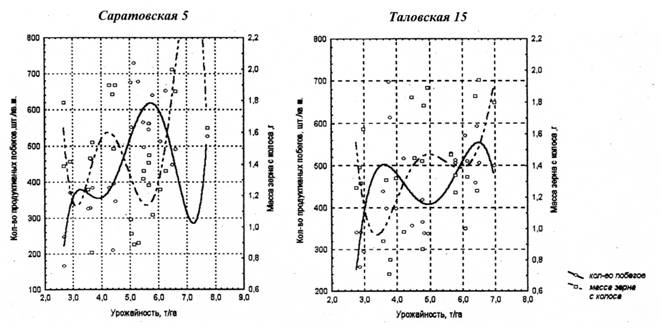
Рис. 4. Зависимость урожайности от количества побегов и массы зерна с колоса аау сортов озимой ржи
Определив коэффициенты корреляции между урожайностью, количеством продуктивных побегов и массой зерна с колоса, мы обнаружили существенные различия между изучавшимися сортами: у Саратовской 5 коэффициент корреляции между урожайностьюа и количеством продуктивных побегов равнялся +0,65, а между урожайностью и массой зерна с колоса - +0,12, у Таловской 15 - соответственно +0,42 и +0,47.
Представив фенотипическую структуру признака лурожайность в виде иерархии модулей по П.П. Литуну и В.А. Драгавцеву (1988), мы сравнили эти два сорта. Для этого были вычислены простые и частные коэффициенты корреляции между составляющими каждого модуля, а также коэффициенты множественной корреляции между результирующим и компонентными признаками для каждого модуля. При этом были обнаружены значительные различия между коэффициентами парной корреляции массы зерновки с количеством зёрен в колосе: -0,79 у Саратовской 5 и -0,39 у Таловской 15. У Саратовской 5 достоверно сильнее связь между сохранностью растений к уборке и количеством всходов: +0,78 против +0,17 у Таловской 15. Они же существенно влияли на количество продуктивных растений, а те, в свою очередь, на количество продуктивных побегов. Наблюдались также значительные различия и по величине множественной корреляции между продуктивной кустистостью растения и обуславливающими её общим количеством побегов на растении и долей продуктивных побегов: +0,25 у Саратовской 5 и +0,59 у Таловской 15.
Вероятно, эти различия обуславливают бoльшую зависимость урожайности сорта Саратовская 5 от количества продуктивных побегов на единице площади, чем это можно видеть у сорта Таловская 15, и, практически, отсутствие у этого сорта корреляции между урожайностью и массой зерна с колоса.
Относительно слабая отрицательная корреляция между количеством зёрен в колосе и массой зерновки и сильная положительная (0,83) между массой зерна с колоса и массой зерновки при постоянном значении количества зёрен в колосе представляют значительный интерес в связи с реальной возможностью увеличения продуктивности у сортов типа Таловския 15 за счёт увеличения массы зерна с колоса. Относительно большая стабильность этого признака, как и большинства элементов продуктивности колоса, определенная нами разными способами, облегчает селекционную работу в этом направлении, что было уже доказано селекционерами, работавшими с озимой рожью (Скорик В.В., 1975, 1993; Dill P., 1983; Ласкин В.П., Бамбышев У.С., Валеев А.З., 1987).
Густота продуктивного стеблестоя, от которой больше всего зависит урожайность сорта Саратовская 5, и связанные с ней признаки (сохранность растений, продуктивная кустистость) относятся к сильноварьирующим и поэтому являются трудноселектируемыми. Это делает морфотип, аналогичный этому сорту, более уязвимым и поэтому менее перспективным. К тому же, увеличение густоты у этого морфотипа обострит проблему устойчивости к полеганию. Поэтому дальнейшее повышение потенциала продуктивности этого морфотипа требует изменения архитектоники растения в направлении уменьшения высоты и/или увеличения прочности стебля, что связано с новыми проблемами.
СТРУКТУРНЫЙ АНАЛИЗ И ЕГО ЗНАЧЕНИЕ ДЛЯ СЕЛЕКЦИОННОЙ ПРАКТИКИ
Метод структурного анализа, предлагаемый З.А.Морозовой (1983) не заменяет обычный анализ, а, опираясь на него, позволяет получить дополнительную ценную информацию, скрывающуюся обычно за средними значениями признаков, получаемыми при обычном анализе.
В своей работе мы попытались дать оценку этому методу с точки зрения селекционера, апробировав его на обширном материале, внеся при этом некоторые дополнения в предлагаемую автором методику, суть которых сводится к следующему.
- Дополнили предлагаемый метод характеристикой качества сформировавшихся зерновок.
- Для характеристики степени сбалансированности органогенеза ввели критерии несогласованности. Критерий I (КI) представляет собой сумму максимальных отклонений по интенсивности, асимметрии и эксцессу, критерий II (КII) - сумму максимальных отклонений по асимметрии и эксцессу.
Вычисленные таким образом коэффициенты несогласованности мы сопоставили с урожайностью изучавшихся в КСИ, ПСИ и ЭСИ сортов (табл. 4). Полученные данные свидетельствуют о том, что низкоурожайные сорта и наборы характеризуются меньшей согласованностью процессов органогенеза. При этом второй критерий (KII) больше связан с уровнем урожайности. В 80% случаев у диплоидов и в 100% случаев у тетраплоидов наблюдалось совпадение уровня урожая со степенью сбалансированности процессов органогенеза, которую отражает этот показатель. Величины первого критерия (КI) не всегда, особенно у диплоидов, совпадали с уровнем урожайности изучавшихся сортов.
Таблица 4 - Критерии несогласованности процессов органогенез у сортов, различавшихся по урожайности
Год |
Питомник |
учшие сорта |
Худшие сорта |
||||||
учшая поло-вина набора |
два наилуч-ших сорта |
худшая поло-вина набора |
два наихуд-ших сорта |
||||||
КI |
KII |
КI |
KII |
КI |
KII |
КI |
KII |
||
Д и п л о и д н ы еаа с о р т а |
|||||||||
1992 |
аКСИ |
0,94 |
0,48 |
0,40 |
0,25 |
1,47 |
0,76 |
1,45 |
0,50 |
1993 |
аКСИ |
1,74 |
0,76 |
2,10 |
0,90 |
1,62 |
1,12 |
1,20 |
1,10 |
1994 |
аКСИ |
0,73 |
0,30 |
0,57 |
0,17 |
0,60 |
0,33 |
1,53 |
1,08 |
1995 |
аКСИ |
1,30 |
0,70 |
1,55 |
1,45 |
2,05 |
1,12 |
2,10 |
1,25 |
аПСИ |
2,08 |
1,44 |
1,70 |
1,05 |
2,62 |
1,66 |
1,60 |
0,85 |
|
аЭСИ I |
1,07 |
0,59 |
0,80 |
0,60 |
1,91 |
1,25 |
1,75 |
1,20 |
|
аЭСИ II |
2,33 |
1,38 |
2,10 |
0,75 |
2,44 |
1,39 |
2,02 |
0,92 |
|
аЭСИ III |
2,97 |
1,75 |
2,45 |
1,35 |
1,47 |
0,90 |
4,10 |
2,65 |
|
1996 |
аКСИ |
0,76 |
0,50 |
1,30 |
0,95 |
1,22 |
0,82 |
1,95 |
1,40 |
аПСИ |
2,50 |
1,62 |
1,70 |
0,85 |
1,56 |
1,00 |
0,95 |
0,55 |
|
1997 |
аКСИ |
1,25 |
0,75 |
0,81 |
0,32 |
1,55 |
0,65 |
2,08 |
0,98 |
аПСИ |
2,25 |
1,65 |
1,30 |
0,72 |
1,70 |
0,88 |
1,70 |
0,98 |
|
1998 |
аКСИ |
1,15 |
0,65 |
1,56 |
0,72 |
2,30 |
1,70 |
1,88 |
1,00 |
аПСИ |
1,60 |
1,00 |
1,19 |
0,49 |
1,15 |
0,95 |
1,00 |
0,54 |
|
Средний |
1,62 0,189 |
0,70 0,133 |
1,40 0,162 |
076 0,102 |
1,69 0,145 |
1,11 0,116* |
1,81 0,203 |
1,07 0,141 |
|
Т е т р а п л о и д н ы еа с о р т а |
|||||||||
1993 |
КСИ |
0,82 |
0,62 |
0,45 |
0,25 |
1,15 |
0,70 |
0,40 |
0,30 |
1994 |
КСИ |
1,70 |
1,28 |
1,00 |
0,70 |
2,35 |
1,45 |
2,50 |
1,50 |
1995 |
КСИ |
1,21 |
0,64 |
0,40 |
0,30 |
1,71 |
0,93 |
1,75 |
0,90 |
ПСИ |
1,65 |
0,93 |
1,80 |
0,95 |
2,55 |
1,50 |
2,55 |
1,50 |
|
1996 |
КСИ |
0,80 |
0,55 |
0,98 |
0,58 |
2,15 |
1,65 |
2,14 |
1,59 |
1997 |
КСИ |
0,80 |
0,30 |
0,87 |
0,23 |
0,80 |
0,60 |
0,56 |
0,53 |
1998 |
КСИ |
0,95 |
0,60 |
0,95 |
0,60 |
1,40 |
0,95 |
1,40 |
0,95 |
Средний |
1,13 0,150 |
0,70 0,119 |
0,92 0,175 |
0,510,101 |
1,73 0,246 |
1,11 0,158 |
1,610,303 |
1,04 0,193* |
|
Так как эти два критерия отличаются лишь тем, что КI учитывает интенсивность органогенеза в разные периоды, а KII нет, мы изучили этот показатель более тщательно. Оказалось, что не только разные морфотипы, но и разные сорта имеют свои особенности по интенсивности морфогенеза в разные периоды.
Общей закономерностью для всего изучавшегося материала является постепенное затухание интенсивности формообразовательного процесса (табл. 5). Наиболее интенсивным он является в первый период и наименее интенсивным - в третий период онтогенеза. Исключение составляют тетраплоидные формы, у которых интенсивность формообразования наименьшая во второй период, а в третьем наблюдается её увеличение. Это согласуется с известной плохой озернённостью колоса и большой массой зерновки у тетраплоидов (Тороп А.А., 1969).
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
Авторефераты по всем темам >>
Авторефераты по биологии