
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Научно-практическое обоснование интенсификации пчеловодства за счет использования биологических особенностей медоносных пчел и применения апипродуктов
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАаа 3.2.2.а Использование экстерьерных признаков для контроля чистопородности пчел. Установлено, что наиболее контрастными показателями оказались: длина хоботка, кубитальный индекс, форма задней границы воскового зеркальца 5-го стернита и дискоидальное смещение. Нами разработан экспресс-тест наа чистопородность (рис. 2). По четырем сторонам диаграммы указаны значения основных признаков, ограничивающие рамки породы. Нужно определить значения этих четырех признаков в исследуемой пробе пчел и отложить их на диаграмме. Точка, их объединяющая, укажет породность пчел.
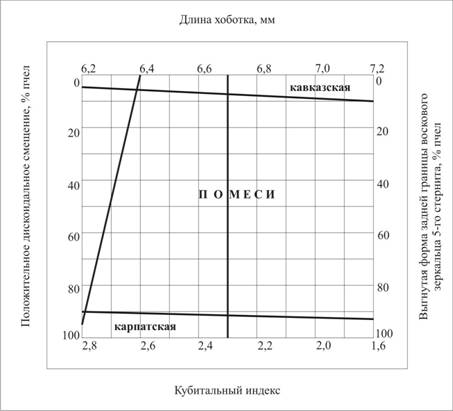
Рис. 2.а Диаграмма для экспресс-теста пчел на чистопородность
3.2.3. Оценка породности пчел в совхозах Северного Кавказа. Экспресс-тест показал, что для поддержания чистопородности при формировании племенного ядра в совхозе Кисловодский может использоваться не более 20% пчелосемей, отвечающих стандарту карпатской породы по всем четырем признакам (табл. 7), в пчелосовхозе Беканский - не более 19%, в совхозе Кабардино-Балкарский - не более 6%. (Данный подраздел выполнялся совм. с Петровым А.И.)
Таблица 7
Доля пчелосемей, отвечающих стандарту карпатской породы
по основным экстерьерным признакам в совхозе Кисловодский, %
По длине хоботка |
По кубитальному индексу |
Поа дискоидальному смещению |
По форме задней гран воск зерк 5-го стернита |
79,6 |
79,6 |
38,6 |
20,5 |
3.2.4. Разработка концепции метода стабилизации генетической основы медоносных пчел.
а) Обследование и использование аизолированного случного пункта. По лоцманским картам была выявленаа возможность использования в качестве изолированного случного пункта Каменниковский полуостров, вдающийся в акваторию Рыбинского водохранилища на 30 км. Во время цветения малиныа обследовали его территорию на отсутствие медоносных пчел. В результате спаривания пчелиных маток с трутнями установлено, что изоляция была надежная и спаривания прошли в чистоте. Таким образом, этот полуостров был использован для реализации запланированной схемы инбридинга (табл. 8).
Таблица 8
Схема инбридинга
Поколение |
Физиологически |
Генетически |
F1 F2 F3 F4 F5 |
сестра х брат сестра х брат племянница х дядька сестра х брат племянница х дядька |
дочь х мать дочь х мать внучка х бабка дочь х мать внучка х бабка |
б) Характеристика инбредной группы пчелиных семей, находящейся в условиях пространственной изоляции.а Плодная карпатская матка-родоннанчальница № 192 (вместе с ее семьей привезена из Иршавского района Закарпатской области) светло-бурой окраски. Ее плодовитость - 1952 яиц в сутки, показательа недопроизводства расплода - 6%. В опытной группе 30 семей. Матки каждого последующего поколения являлись дочками маткам предыдущего.
Таблица 9
Изменчивость признаков маток по поколениям (n = 30)
lim |
M |
m |
Cv, % |
|
Масса неплодных маток, мг |
||||
F1 |
171 - 231 |
201,4 |
2,88 |
7,85 |
F2 |
163 - 212 |
183,6 |
2,51 |
7,52 |
F3 |
162 - 224 |
182,4 |
2,44 |
7,34 |
F4 |
168 - 218 |
186,9 |
2,18 |
6,43 |
F5 |
169 - 216 |
192,7 |
2,36 |
6,75 |
Яйценоскость маток, яиц/сут. |
||||
F |
- |
1952,0 |
- |
- |
F1 |
968 - 2864 |
1775,2 |
141,32 |
24,12 |
F2 |
1131 - 2270 |
1710,4 |
85,30 |
18,46 |
F3 |
1092 - 2614 |
1688,7 |
73,08 |
20,62 |
F4 |
1100 - 2547 |
1907,6 |
125,17 |
22,98 |
F5 |
1240 - 2800 |
1880,5 |
130,44 |
25,47 |
Маток получено пять поколений, трутней - три. Отбор отсутствовал. Трутни выводились в каждой семье в одинаковом количестве (по 10 тыс. шт.). Спустя три летних сезона мы получили результаты. У маток изучались плодовитость и масса. Из таблицы 9 видно, что матки во всех поколениях обладали высокой яйценоскостью - от 1688,7 яиц/сут. в F3 до 1907,6 яиц/сут. - в F4. В F1 плодовитость составила 1775,2 яиц/сут., в F2 - 1710,4 яиц/сут., в F5 - 1880,5 яиц/сут. Разность по яйценоскости недостоверна.
Второе и последующие поколения были инбредными. Во втором и в третьем поколениях яйценоскость снизилась, надо полагать, в связиа с депрессией, вызванной повышением гомозиготности. В последующих поколениях - увеличилась.аа
Средняя масса неплодных маток в первом (аутбредном) поколении составила 201,4 мг, во втором (инбредном) поколении она снизилась до 183,6 мг (Р>0,999), в третьем поколении - до 182,4 мг (Р>0,999). В четвертом поколении она составила 186,9 мг (P>0,999), а в пятом - произошло повышение до 192,7 мг (Р>0,95). Матки были крупные, активные, схожие с родоначальницей по окраске. Лишь 3,3% из них были уродливыми. Разнообразие маток по яйценоскости в начале инбридинга снижается. В первом (аутбредном) поколении оно составило 24,12%, во втором (инбредном) снизилось до 18,46%, в F3а и F4 атакже было относительно невысоким - 20,62% и 22,98%, в F5 - поднялось до 25,47%. По весу матки разнились гораздо меньше, чем по плодовитости. Коэффициент вариации в F1 составлял 7,89%, в F2 - 7,52%, в F3 - 7,34%, в F4 - 6,43%, в F5 - 6,75%. Установлено также,а что на первых этапах инбридинга (F2; F3) средние величины линейных экстерьерных показателей и их изменчивость снижаются, а в последующих поколениях (F4; F5) стабилизируются или начинают повышаться. Влияние инбридинга на указанные признаки хотя и прослеживается, но очень незначительно и только на начальных этапах.а
в)а Распределение половых аллелей и недопроизводство расплода. На всех пасеках мира встречается, так называемый, пестрый расплод генетической природы (недопроизводство расплода). Он представляет собой картину, когда на фоне запечатанного расплода имеются открытые ячейки с личинками разных возрастов. Таких незапечатанных ячеек может быть до 50%, но не более. Это проявление влияния различных сочетаний в серии множественных аллелей гена пола. Данное явление - рудимент прежнего способа определения пола потомства, бывшего у предков перепончатокрылых и дошедшего до современных пчел. Чем больше эта пестрота, тем медленнее развивается семья, что влечет за собой недопроизводство продукции.
Матка с половыми аллелями АВ, оплодотворенная трутнем с аллелем С (АВ х С), производит исключительно гетерозиготные яйца, с 100% выживаемостью. Матка с аллелями АВ, спаренная с трутнями, несущимиа аллель А, произведет 2 вида оплодотворенных яиц: АА и АВ. аИз гетерозиготных яиц АВ разовьются рабочие пчелы, а из гомозиготных яиц АА - диплоидные трутни, которых рабочие пчелы распознают и уничтожат на стадии молодых личинок. Следовательно, только 50% яиц разовьются в рабочих пчел. Семья останется слабой и не проявит высокую продуктивность.
Перед началом эксперимента была построена математическая модель инбредной изолированной группы пчелиных семей, которая отражает распределение половых аллелей в 17 поколениях у 30 маток. Считается, что матку в естественных условиях осеменяют от 8 до 12 трутней. Мы приняли условие, что матку осеменили 10 трутней. Далее условия виртуальной модели соответствовали условиям практической схемы. Отличие заключалось в том, что в практической схеме в каждом поколении было постоянное количество маток (30 шт.), а в виртуальной модели предполагалось в каждом поколении иметь столько маток, сколько могло возникнуть комбинаций половых аллелей в их генотипах.
В виртуальной модели мы приняли условие, что среди десяти трутней, осеменивших матку, один несет в себе половой аллель, идентичный одному из половых аллелей матки. Это сделано для того, чтобы приблизить исходные условия математической модели к исходным условиям практической схемы. А исходным условием практической схемы являлась сама матка-родоначальница № 192. Известно, что у нее недопроизводство расплода составляло всего лишь 6%.
Итак, виртуальная модель имела следующий вид (рис. 3).
матка-
родоначальницааа Ха трутни, ее осеменившие
ав ааа с д е f g h i j kа
аа
возможные комбинации
от спаривания
а а ва
аа ас вс
аа adаа bd
аа ae beаа трутни от матки-аа
а родоначальницы
F1а af bf Ха а, в
аа ag bg
аа ah bh
аа aiа bi
аа ajа bj
аа akаа аbk
Рис. 3.а Схема математической модели инбридинга
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
Авторефераты по всем темам >>
Авторефераты по биологии