
Конспект лекцій дисципліни «основи біотехнології рослин»
| Вид материала | Конспект |
- Програма вступних фахових випробувань за спеціальністю «фізіологія рослин» освітньо-кваліфікаційний, 164.35kb.
- Конспект лекцій з навчальної дисципліни «основи охорони праці» для студентів 3 курсу, 15.25kb.
- Конспект лекцій з дисципліни „Радіоекологія для студентів спеціальності 040106 „Екологія,, 1393.76kb.
- В. О. Кодін конспект лекцій з дисципліни «Основи реконструкції історичних міст» для, 703.58kb.
- Конспект лекцій Удвох частинах Частина 1 Суми, 2323.63kb.
- Конспект лекцій Удвох частинах Частина 2 Суми, 1998.47kb.
- Конспект лекцій Суми Видавництво Сумду 2010, 2423.29kb.
- Конспект лекцій з дисципліни „ Управління інноваційним розвитком" для студентів факультету, 2082.69kb.
- Конспект лекцій з дисципліни «Оподаткування підприємств», 2062.58kb.
- І. М. Золотарьова конспект, 427.49kb.
КОНСПЕКТ ЛЕКЦІЙ
ДИСЦИПЛІНИ «ОСНОВИ БІОТЕХНОЛОГІЇ РОСЛИН»
ДЛЯ СТУДЕНТІВ АГРОНОМІЧНОГО ФАКУЛЬТЕТУ ВНАУ
Лекція 1.
Тема: Предмет та методи сільськогосподарської біотехнології.
- Біотехнологія як наука, її мета та завдання;
- Використання біотехнології в рослинництві та інших галузях народного господарства;
- Історія розвитку біотехнології;
- Культура ін вітро, основні терміни та поняття.
1.Швидкий ріст населення Земної кулі спонукає до збільшення виробництва продуктів харчування. Відомо, що 1,9 млрд. тон (99%) уживаної сухої речовини людство одержує в якості продуктів землеробства, 1% - з морів, океанів і прісноводних басейнів. Рослинні продукти складають 93% їжі людини, інші 7% - тваринного походження. Істотне збільшення врожаю с.г. культур за останні десятеліття досягнуто за рахунок хімізації, механізації і меліорації сільського господарства, що призвело до виникнення ряду економічних та екологічних проблем: забруднення навколишнього середовища, виснаження енергетичних, грунтових, водних та біологічних ресурсів. Постало питання пошуку нових підходів, які б дозволили не тільки підвищити врожаї і поліпшити якість культур, але були б економічно вигідними і не завдавали шкоди навколишньому середовищу.Одним з таких підходів є біотехнологія.
Біотехнологія - це сукупність технічних прийомів, що передбачають використання живих клітин або біологічних мікромолекул з метою вирішення конкретних або глобальних проблем біосфери та людства.
Термін «біотехнологія» походить від грецьких слів “bios” -життя , “techne” – майстерність,“logos”- вчення. Сама назва вказує про поєднання можливостей біології і техніки. Клітинна і генетична інженерія рослин, тварин і людини, виробництво антибіотиків, біогазу, штучного білку, інсуліну, гормонів росту, алколоїдів, біовимивання благородних металів, пестецидів за допомогою бактерій – це далеко не повний перелік питань, якими займається світова біотехнологія.
^ Біотехнологія рослин – це сукупність технічних прийомів для модифікацій, поліпшення, створення та розмноження рослинних організмів, одержання з них корисних речовин. Фактично - це вирощування і маніпуляції з клітинами, тканинами і органами рослин поза оганізмом на штучних поживних середовищах у контрольованих умовах, що дозволяє:
- проводити швидке розмноження у дуже великих кількостях;
- отримувати безвірусний матеріал;
- поліпшувати існуючі та створювати нові сорти рослин;
- вивчати складні процеси росту та розвитку рослин;
- створювати принципово нові біологічні технології для сільського господарства, медецини, харчової та хімічної промисловості.
^ Основна мета досліджень біотехнології – це поліпшення існуючих та одержання нових сортів, видів, гібридів високопродуктивних форм рослин з поліпшеними показниками якості продукції і стійких до хвороб та шкідників (корисними ознаками для людини). Для цього у сучасній біотехнології виділено три основних напрями:
- технології, що грунтуються на використанні культури клітин, тканин та органів рослин;
- ДНК-технології (молекулярно-генетичні методи аналізу рослин);
- отримання трансгенних рослин.
Варто підкреслити, що застосування біотехнологічних методів не можливе без традиційних методів генетики, селекції, фізіології, біохімії, тощо.
2.Рослинні клітини, при порівнянні з клітинами тварин і людини, мають велику перевагу – здатність у визначених умовах і на відповідних живильних середовищах регенерувати (відновлювати) повноцінні рослини. Дослідження багатьох вчених (Хаберленда, 1902р., Уайта, 1934р, Готре, 1934р) показали, що ізольовані клітини рослин діляться і дають початок недиференційованій масі, названій калусом. А зусилля багатьох вчених (Скуг, Міллер, 1957р), показали, що при повторному поміщенні цієї тканини на живильне середовище з фітогормонами дає успішне культивування і регенерацію рослин з бажаними корисними ознаками.
Людство очікує створення таких клітинних культур, за допомогою яких можна буде виготовляти цінні лікарські препарати, боротися з рядом спадкових, ракових серцево-судинних і інших захворювань, сприяти очищенню і поліпшенню екологічного стану навколишнього середовища. Молекули ДНК, виділені з будь-яких організмів, можуть бути розірвані у визначених точках окремих сегментівза допомогою спеціальних ферментів, а також зшиваються за допомогою інших ферментів, у результаті чого утворюються нові рекомбінантні ДНК. Зміна геному на клітинному рівні відкриває нові можливості для лікування хвороб людини, у діагностиці вірусних хвороб рослин, мікробіологічного виробництва ферментів, амінокислот, вітамінів та ін. хім. речовин (інсулін, інтерферон).
Генетична інженерія відкриває перед селекцією рослин нові перспективи, пов’язані з можливістю перенесення в них генів від бактерій, грибів, екзотичних рослин та навіть людини і тварини, що долає бар’єри, які існують в традиційній селекції. Вже зараз використовуються методи штучного запліднення в пробірці, клонування, трансплантації ембріонів, одержання трансгенних тварин і рослин.
Слід зазначити, що великі корпорації зарубіжних країн, особливо США, інвестують великі кошти в програми з використання геннної інженерії, особливо створення нових форм трансгенних рослин.
Передбачається, що прибуток від використання трансгенних рослин уже до 2020 року може скласти від 50 до 100 млрд. доларів.
3. Перші спроби культивування ізольованих тканин рослин були зроблені наприкінці XIX - на початку XX стол. німецькими вченими Г. Рехінгером (1893р), який одержав і вивчав калусні формування на коренеплодах цукрового буряка, Г.Хаберландтом, який першим чітко обгрунтував ідею про можливість вирощування окремих рослинних тканин на штучних живильних середовищах. Детальна розробка цього методу була здійснена у 30-х роках американцем Пилипом Уайтом і французом Роже Готре. Це був перший історичний етап в розвитку біотехнології як науки.
У період між 1940-1960 рр. учні і послідовники Уайта закріплюють успіх. Збільшується число видів рослин, тканини яких вирощуються ін вітро (in vitro) – це є другий період історичного розвитку біотехнології.
1949р. – Ф. Калінін почав роботи з культивування ізольованих тканин і органів рослин в Україні.
1957р. – Ф.Скуг і К.Міллер (США) довели можливість одержання коренів і пагонів з калусної тканини при дії цитокінінів та ауксинів. Ж.Морель (Франція) встановив дію гібереліну на проліферацію меристеми, її диференціацію і розвиток цілої рослини.
1957р. – Р.Бутенко розпочала фізіолого-біохімічні дослідження з культивування ізольованих клітин і тканин в умовах регенерації.
1958-1959 рр. – Ф.Стюарт, Дж. Рейнгардт експерементально довели можливість соматичних клітин утворювати зародкоподібні структури (соматичні зародки (нестатеві)).
Початок 70-х років характеризує розвиток нових напрямків біотехнології рослин – соматичної гібридизації і генної інженерії. Серед основних робіт:
1970-і роки – початок робіт з культивування ізольованих тканин і органів в інституті ботаніки АН УРСР. В Нікітському Ботанічному саду розпочаті роботи з культивування ізольованих зародків плодових культур (А.Здруйковська-Ріхтер).
1971-1975рр. – К.Као (Канада), Е.Кокінг (Великобританія), Е Такебе (Японія) розпочали роботи з ізолювання та злитя протопластів.
^ 1980-і роки – Ю. Глєба, В.Сидоров, М.Півень, І Комарницький, Т.Пастернак (Україна) розпочали ряд робіт з ізольованими протопластами. Отримано соматичні гібриди та цибриди, розпочаті перші роботи з генетичної трансформації.
^ 1990-і роки – Е. Пірузян, В.Андріанов, В.Метт, І. Стехін (СРСР) – початок робіт з клонування генів і одержання перших трансгенних рослин.
1994 р. – В США зареєстровано першу трансгенну рослину харчового використання – томат «Флейвр-Сейвр», що характеризується подовженим строком післязбирального зберігання.
1999-2000 рр. – кількість трансгенних сортів рослин у світі досягає 120 і вони займають 40 млн. га посівних площ.
2002-2005 рр. – посівні площі під трансгенними культурами в світі становлять більше 57 млн. га.
4. Біотехнологію рослин можна уявити як проведення різних фізіологічних або генетичних маніпуляцій з клітинами, тканинами, органами рослин, протопластами в штучних контрольованих умовах освітлення, живлення, температури, тобто, в умовах «ін вітро» (від лат. “vitrum” означає скло). Цей термін вказує, що робота проводиться в стерильних і контрольованих умовах у лабораторному посуді (пробірках, колбах, чашках Петрі та ін.). На противагу термін «ін віво» означає, що перебіг процесів у живому організмі відбувається в природніх умовах. А поміщення в умови ін вітро протопластів, клітин або експлантів вживається термін «введення в культуру». Експланти –будь-які частинки тканин, органів рослин, які вводяться в культуру (апекси пагонів або коренів, шматочки листків, бруньки, бутони, зародки, пиляки та ін.)
Структурна організація рослин як біологічних систем складається з різних рівнів: клітина – тканина – орган – організм – популяція. Всі рівні мають свою спецефічність та ступінь складності, але головний взаємозв’язок полягає у тому, що кожна елементарна клітина зберігає у собі всю генетичну інформацію про свій ріст та розвиток. Якщо клітину вилучити з рослини і перенести у штучні контрольовані умови, то відбувається відновлення цілого з частини. Таким чином, кожна рослинна клітинамає потенційні властивості стати повноцінною рослиною, або кажут - вона тотипотентна.
Тотипотентність (від лат. totus – весь, цілий, potencia – сила) – властивість клітин реалізувати власну генетичну інформацію, яка забезпечує їх диференціацію і розвиток до цілого організму. Тотипотентність клітин складає основу наукової ідеології біотехнології рослин. Такі методи біотехнології рослин як мікроклональне розмноження, соматичний ембріоїдогенез, культура зародків і гаплоїдів, соматична гібридизація і генна інженерія пов’язані з реалізацією тотипотентності клітин рослин трьома шляхами:
- формування зачатків коренів або стебла бепосередньо з ексланта чи клітини;
- формування зародкоподібних структур бепосередньо з ексланта чи клітини;
- формування зачатків коренів,стебла або зародкоподібних структур через калусну тканину.
Калусна тканина – це маса неспеціалізованих клітин, які утворюються в результаті пошкодження або при вирощуванні експланта на штучному поживному середовищі.
Лекція 2.
Тема: Регулятори росту і розвитку рослин (РРР).
1. Фітогормони в онтогенезі рослин;
2. Використання регуляторів росту рослин в практиці рослинництва;
3. Класифікація фітогормонів та методи їх отримання;
4. Основні типи РРР, їх фізіологічна дія та використання.
1.Регулятори росту рослин (РРР) – це природні або синтетичні органічні сполуки, які активно впливають на обмін речовинвищих рослин, регулюють фізіологічні та морфогенетичні програми росту і розвитку рослинного організму. Регулятори росту, які продукуються самою рослиною, називаються фітогормонами. За типом дії РРР поділяють на стимулятори та інгібітори. Проте такий поділ умовний, оскільки більшість із них в низьких концентраціях стимулює, а у високих - пригнічує ті чи інші процеси (2,4Д у високих концентраціях є сильний гербіцид, а в низьких -синтетичний регулятор росту рослин).
Фітогормони переміщуються по рослині і впливають на ріст і диференціацію тих тканин і органів, куди потрапляють. Таким чином, фітогормони – солуки, за допомогою яких здійснюється взаємодія клітин, тканин і органів рослин. Вони синтезуються і функціонують в мікрокількостях і на відміну від інших метаболітів, у тому числі вітамінів, здатні викликати в рослині формоутворюючі процеси (ріст коренів, пагонів, утворення квіток, плодів, тощо).
2. З експертних оцінок наукових закладів відомо, що продуктивність нових сортів і гібридів с.г. культур нижча їх генетичної можливості на 40 і більше відсотків. У зв’язку з цим господарства, як правило, висівають насіння польових культур з високими нормами висіву, що призводить до перевитрат високоякісного зерна. Цьому можна на сьогодні запобігти , якщо використовувати синтетичні регулятори росту, які допомагають рослинам протистояти несприятливим факторам і проявити сві генетичний потенціал.
У практиці рослинництва широко використовують синтетичні РРР для підвищення енергії проростання і польової схожості насіння, запобігання виляганню хлібів, регуляції плодоношення, підвищення врожайності, приживання саджанців і живців, а також для полегшення механізованого догляду за рослинами та ін.
^ Препарати: ТУР, емістим, метиур, дипромол, фарбізол, ресин, івин, капонин та ін.
Способи застосування синтетичних РРР – намочування насіння, живців у їх розчинах, обприскування вегетуючих органів рослин. Найефективнішим вважається обприскування розчинами фіторегуляторів у поєднанні із засобами хімічного захисту рослин від хвороб і шкідників (так зване застосування бакових сумішей).
Наприклад, виробнича перевірка в господарствах Черкаської, Вінницької, рівненської областей показала, що передпосівна обробка насіння зернових і зернобобових емістимом (1-2 мл/т) разом з мікроелементами (100 мл композиції в 10 л води) або ж обробка вегетуючих рослин: озимих культур весноюпри денній температурі більше 10ºС, ярих зернових і зернобобових у фазі 3-5 листків дали приріст урожаю від 5 до 8,3 ц/га.
Вчені розробляють нетоксичні синтетичні аналоги гормонів шкідників (ОК-37, ОК-84, ОК-85), які одночасно є ефективними засобами захисту рослин. Так, препарат ОК-37 у дозі 4 г/га викликає загибель 90-100% личинок колорадського жука. Ефективні вони також в боротьбі зі шкідниками цукрових буряків (бурякова попелиця, лучний метелик), капусти, проса, картоплі та ін.
З меттою стимуляції бульбоутворення і підвищення вррожайності середньостиглих сортів картоплі рекомендований препарат фарбізол (малотоксичний, використовується шляхом передпосадкового обприскуваннябульб 0,01% водним розчином). Слід зазначити, що цей препарат зменшує вміст нітратів у бульбах, підвищує кількість аскорбінової кислоти.
Для цукрових буряків рекомендований ефективний регулятор росту ресин. Він поліпшує схожість насіння, стійкістьвегетуючих рослиндо коренеїда, підвищує вміст цукру.
3. У рослин розрізняють такі основні типи фітогормонів: ауксини, цитокініни, гібереліни, абсцизова кислота, етилен, брасиностероїди.
Спеціалісти ведуть пошук нових ендогенних фізіологічно активних речовин, які з часом можуть набути статус фітогормонів. Тільки за останні роки до фітогормонів були віднесені жасмонова кислота, олігосахарини, фузікокцин (Муромцев Г.С., 1996). Кожна хімічна категорія фітогормонів має характерний вплив на ріст і спеціалізацію клітин рослин, що використовується ін вітро. Проте треба чітко запам’ятати, що генотип рослин не змінюється під дією будь-якогорегулятора росту. РРР лише допомагають рослинам розкрити їх генетичний потенціалбез появи нових спадкових властивостей. Крім того, РРР ні в якому випадку не замінюють елементів живлення.
Ауксини
Ауксини (від грецького аuxein - збільшуватись, рости) - група фітогормонів, які регулюють процеси поділу та розтягування клітин, сприяють формуванню коренів, провідних пучків, оплодня.
Історія вивчення ауксинів починається з прізвищ Ч.Дарвіна (1880), П.Бойсен-Йєнсена.(1913), Ф.Вента (1926-1928). У роботах цих дослідників доведено, що вигин колеоптиля злаків до однобічного світлового подразника (фототропізм) пов'язаний із синтезом активного хімічного стимулу в їх верхівках і пересуванням його вниз по рослині. Виділення ауксину у чистому вигляді і його ідентифікація були зроблені у 1934 році.
3 хімічної точки зору ця речовина визначається як індоліл-3-оцтова кислота, хоча частіше її називають гетероауксином. Тільки у 1972 році за допомогою мас-пектроскопії було остаточно доведено, що саме індолілоцтова кислота (ЮК) пересувається з верхівок колеоптилів.
ІОК є основним ауксином рослин, але не єдиним. Відомо багато речовин індольної природи з ауксиновою дією, які синтезовані лабораторним шляхом: індолілпоропіонова, індолілмасляна, індолілпіровиноградна, нафтилоцтова кислота. Існують ауксини неіндольної природи: 2,4-дихлорфеноксиоцтова кислота (2,4-Д); 2,4,5-трихлорфеноксиоцтова кислота (2,4,5-Т); 2,3,6-трихлор-бензойна кислота та ін. Вони у високих концентраціях використовуються як гербіциди.
До ауксиноподібних речовин відносять також деякі природні фенольні сполуки: ферулову кислоту, коніфериловий спирт, ванільні, кофейну кислоту. Їх активність нижча, ніж ЮК. Саме те, що знайдено багато сполук із ауксиновою активністю, говорить про ауксин у множині, а не про ауксин як одну хімічну сполуку.
Ауксини утворюються в молодих частинах рослин, які активно ростуть: точках росту стебла, верхівках коренів, молодих листках, бруньках, квітках і плодах. Транспортування ЮК по рослинних тканинах відбувається полярно - від верхівки пагона до кореня, від листка до верхівки пагона або ж вгору по кореню. В тканинах рослин ЮК знаходиться у двох формах: вільний і зв'язаній. Біологічна активність притаманна тільки вільним формам ЮК, Нерівномірним розподілом ауксинів в осьових органах пояснюють ростові рухи, а також тропізми рослин.
Сучасне уявлення про фізіологічну дію ауксинів
Класична дія ауксинів - посилення росту за рахунок стимуляції розтягнення клітин. ЮК активує роботу мембранної помпи іонів Н+, знижує рН позаклітинного середовища і посилює, тим самим, пластичність клітинних стінок. Підвищення концентрації протонів послаблює водневі зв'язки між мікрофібрилами целюлози, геміцелюлози і ксилоглюканів, що дозволяє їм ковзатись відносно одна одної при підвищенні тургорного тиску. Цікавим спостереженням є "кислий ріст" клітин за рахунок розтягнення клітинних стінок без ауксинів при рН близьких до 3,0. Можливо, що у розпушуванні клітинної станки беруть участь ферменти із низькими значеннями рН.
Однак видовження клітини - це не просто розтягнення клітинної стінки. Про це свідчить той факт, що клітинні стінки при розтягненні не стають тоншими, тобто одночасно відбувається синтез de novo компонентів стінки.
Це, в свою чергу, дає підстави враховувати участь ауксинів в синтезі і транспорті нових клітинних полісахаридів. Вченими вже встановлена причетність ІОК до стимуляції синтезу різних типів РНК (м-РНК, т-РНК, р-РНК) і білка в клітинах, які розтягуються.
ІОК зумовлює явище апікального домінування, коли апікальна меристема гальмує ріст бокових меристем. Домінування верхівки — класичний приклад того, як одна частина рослини контролює іншу за допомогою фітогормонів. Природу апікального домінування пояснюють декілька гіпотез:
1) Верхівочна меристема (брунька) найбільш насичена ауксином, і є атрагуючим центром притягнення води та різних поживних речовин, яких не вистачає для бокових бруньок;
2) Під дією ауксину синтезується інгібітор росту, який проникає у бокові меристеми і гальмує їх розвиток.
Синтетичні ауксини широко використовуються на практиці для укорінення черенків, покращення зростання тканин при щепленні, для запобігання опадання зав'язі, а також як сильні гербіциди. Так, суміш 2,4-Д і 2,4,5-Т застосовувалась США під час війни у В'єтнамі (дефоліант "Ейджент Оранж"), Слід зазначити, що ЮК у супероптимальних концентраціях також здатна гальмувати ріст, виступаючи вже інгібітором.
Цитокініни
Цитокініни були відкриті в результаті інтенсивної роботи з культурою тканин, яку проводив американський дослідник Ф.Скуг у 1945-1955 роках. 3 хімічної точки зору вони є похідними 6-амінопурину (аденіну). На сьогодні ідентифіковано близько і5 природних цитокінінів (зеатин, зеатинрибозид, зеатинрибозидфосфат та ін.), В умовах ін вітро широко використовують високоактивний синтетичний препарат кінетин ( одержують при ав-токлавуванні ДНК) і бензиламінопурин /БАП/.
Більше всього цитокінів там, де проходить швидкий поділ клітин. Особливо багато міститься їх у проростаючому насінні, достигаючих плодах, верхівках коренів, пухлинних клітинах; менше - у стеблах, листках. Синтез цитокініів у дорослих рослин локалізується в апікальній меристемі коренів, звідки вони транспортуються у надземні органи з ксилемним соком.
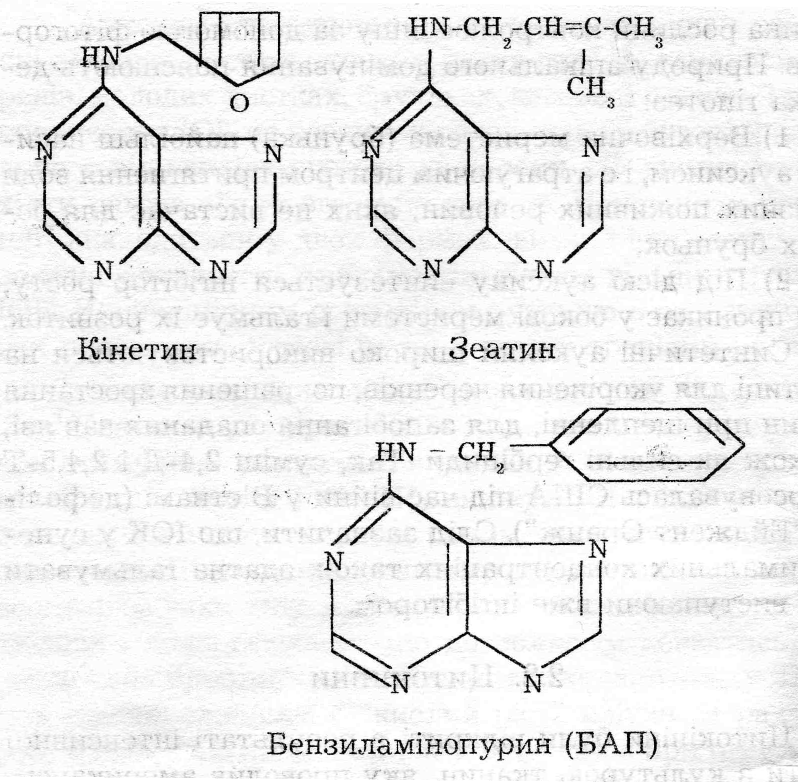
^ Фізіологічна дія цитокінінів
Цитокініни поліфункціональні у своїй дії на різних етапах росту і розвитку рослин. Сучасна наробка фактичних даних і їх обговорення дозволяють акцентувати увагу на двох основних ефектах фізіологічної дії цитокінінів:
1 - стимуляція процесів клітинного поділу і диференціації;
2 - затримка процесів старіння відокремлених органів.
Структурна подібність цитокінінів з аденіном - компонентом ДНК і РНК - не випадкова і пов'язана з їх функціональними особливостями. Достовірно встановлена специфічність дії цитокінінів як стимуляторів мітозу.
При вирощуванні ізольованих клітин ін вітро особливо чітко виявляється залежність поділу клітин від введення до поживного середовища екзогенного цитокініну.
При взаємодії з ауксинами і гіберелінами цитокініни беруть участь у новоутворенні і диференціації органів ін вітро та ін віво, що пов'язують з їх впливом на біосинтез ДНК, РНК, білків, а також на перерозподіл продуктів обміну у рослині.
Цитокініни стимулюють розвиток латеральних точок росту (бокових бруньок), тобто беруть участь у подоланні апікального домінування. У підрозділі 2.1. ми розглядали, що ауксини утворюються у верхівках, приміщуються вниз по стеблу і пригнічують розвиток бокових бруньок. Чим ближче бокові бруньки розміщені до верхівки, тим сильніше пригнічені, і навпаки. Таке гальмування росту можливо зняти шляхом видалення верхівки або нанесенням цитокініну на бокові бруньки. Саме цим способом домагаються утворення пагонів з усіх пазухових меристем бруньки ін вітро на підвищених концентраціях цитокінінів в поживних середовищах.
Здатність цитокінінів затримувати старіння проявляється на листках. Відокремлений листок у воді швидко старіє, що проявляється у пожовтінні, втраті ним білка, ДНК, РНК. Якщо нанести на такий листок краплю кінетину, то оброблена ділянка залишиться зеленою. Затримка старіння листка також відбувається при утворенні коренів на черешку листка. З цього можна зробити висновок, що цитокініни, які утворюються в коренях, переміщуються в листки і підтримують структурно-функціональну життєдіяльність листків. Цитокініни продовжують тривалість життя ( об'єкти довго зберігають гарний вигляд) свіжої капусти, салату та інших зелених овочів, а також зрізаних квітів, що використовується на практиці.
Гібереліни
Гібереліни були відкриті у 1926 році японським вченим Е.Куросавою при вивченні ураження рису грибом Gibberella. Хворі рослини сильно витягувались у довжину, вилягали і гинули. Екстракт гриба, а також виділена з нього у кристалічному вигляді речовина викликали аналогічну симптоматику захворювання. Ці речовини були названі гіберелінами. (від назви гриба). 3 хімічної точки зору гібереліни відносяться до сполук дитерпеноїдів. Вихідними сполуками для їх синтезу у клітинах є ацетат і мевалонова кислота. Для комерційних цілей гібереліни отримують з культури грибів шляхом мікробіологічного синтезу.
На сьогодні відомо близько 60 природних гіберелінів, які позначаються скорочено ГК1; ГК2 і т.д. Найбільш типовим представником комплексу гіберелінів є гіберелова кислота (ГК3 ), яка знайдена в усіх вивчених рослинах:
У вищих рослин найбільш багаті на гібереліни швидкоростучі органи - молоді апікальні листки, бруньки, незріле насіння і плоди.
Звідси вони мігрують по флоемі і ксилемі уверх-вниз, тобто гібереліни не виявляють полярності транспорту, яка характерна для ауксинів. Велике значення для утворення гіберелінів у рослині має світло (воно сприяє їх біосинтезу в хлоропластах листків).
^ Фізіологічна дія гіберелінів
Різнопланові експерименти з вивчення фізіологічної дії гіберелінів дозволяють зробити висновок, що гібереліни - компоненти систем, які регулюють ріст і розвиток рослин .
Найбільш виражена дія гіберелінів - їх здатність стимулювати ріст, видовження стебла за рахунок розтягування клітин ( а не їх поділу), що вказує на подібність із ефектом дії ауксинів. За допомогою гіберелінів відновлюють нормальний ріст карликових сортів гороху, кукурудзи або перетворюють карликову форму квасолі у витку ліану. На практиці гіберелінами обробляють коноплю та льон для підвищення виходу волокна.
Другий класичний ефект дії гіберелінів пов'язаний з виходом насіння злакових із стану спокою. Після того, як насіння поглине крізь мікропіле воду, зародок в зоні щитка починає синтезувати гібереліни ГА3, ГА4. Гібереліни переміщуються в алейроновий прошарок, що оточує ендосперм, і стимулюють утворення гідролітичних ферментів (альфа-амілаз та ін.). Ферменти починають розщеплювати запасний крохмаль ендосперму до простих цукрів, які використовуються для росту зародка. Таким чином, гібереліновий "запуск" утворення амілаз є необхідною умовою для проростання насіння. Механізм цього процесу остаточно не з'ясований. Вважають, що гібереліни стимулюють синтез м-РНК, які специфічні для амілаз.
Важливе значення гіберелінів у процесі яровизації і цвітіння. Яровизація - реакція рослин на вплив низьких позитивних температур (+2 - +10° С) у певний період онтогенезу. Центром сприйняття яровизаційного впливу в рослині може бути точка росту або будь-яка зона, в якій відбувається поділ клітин. Яровизація проявляється у прискоренні початку періоду плодоношення (цвітіння). Встановлено, що у ході яровизації підвищується рівень гіберелінів, що дозволяє холодову обробку замінити обробкою неяровизованих рослин гіберелінами. Так, екзогенно введений гіберелін у багатьох дворічних рослин виключає потребу в яровизації і викликає їх цвітіння.
Гібереліни викликають партенокарпію - процес, при якому плоди розвиваються без запліднення. Для цього квітки рослин обприскують розчином гібереліну, що практично застосовується у виноградарстві (отримують без насіннєві ягоди великого розміру).
Серйозної уваги заслуговує здатність гіберелінів зміцнювати у деяких рослин вираженість статі. Встановлено, що вміст гіберелоподібних речовин в рослинах огірка корелює із кількістю чоловічих квіток. Багаточисельними роботами показана маскулінізація (зсування статі у чоловічий бік) рослин родини гарбузових гід впливом гіберелінів.
^ Абсцизова кислота і етилен
Для оптимальної регуляції росту необхідні не тільки стимулятори (типу ауксинів, цитокінінів, гіберелінів), але й інгібітори, до яких відносять абсцизову кислоту і етилен.
Абсцизова кислота (АБК);
У 60-хроках в результаті незалежних досліджень А.Едікота та П.Уорінга були відкриті специфічні речовини-інгібітори деяких процесів у рослин. А. Едікот виділив з молодих плодів бавовнику сполуку, яка викликала опадання листків цієї рослини. Автор назвав и абсцизином (від лат. abscisio - опадати, спадати). П.Уорінг показав, що екстракт з листків берези індукує стан спокою березових бруньок. Речовину, яку було виділено у кристалічному вигляді, назвали дорміном (фр. dormir - спати). Виявилось, що дормін теж викликав опадання
коробочок бавовнику, а за хімічною будовою дормін та абсцизин подібні. У 1967 році було вирішено назвати цю речовину абсцизовою кислотою (АБК).
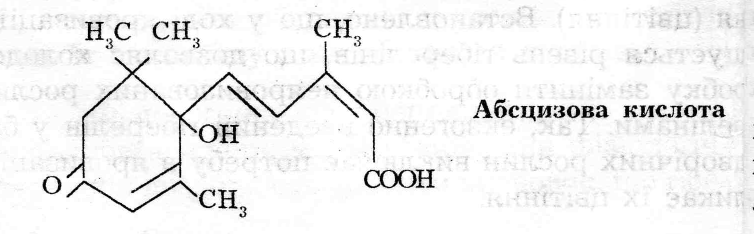
3 хімічної точки зору АБК відноситься до терпеноїдів із складною будовою. Більшість органів вищих рослин здатні синтезувати АБК, транспорт речовини відбувається по провідній системі (флоемі, ксилемі). Вміст АБК визначають за допомогою біотестів та фізичними методами.
Механізм дії на сьогодні невідомий. Експериментально доведена функція АБК як природного інгібітора в явищі геотропізму кореня. Замикання продихів в умовах посухи супроводжується збільшенням концентрації АБК. Дія АБК пов'язана із спокоєм бруньок і насіння, опаданням квіток, плодів, старінням і дозріванням.
У світовій практиці АБК використовують для обприскування плодових, щоб викликати одночасне опадання плодів для скорочення часу при збиранні врожаю.
Проводяться дослідження дії АБК в умовах ін вітро. Додавання до поживного середовища 7,6 мкМ АБК впливало на ріст і формування ембріогенних клітинних агрегатів у суспензійних культур клітин ярої пшениці Тriticum durum L (Шаяхметов І.Ф., Шакирова Ф.М., 1996). Автори вважають, що АБК бере участь у зміненні балансу фітогормонів і сприяє "дозріванню" соматичних ембріоїдів. АБК попереджує утворення кореневих структур, що
виводить процес регенерації з тупикового напрямку - ризогенезу, після якого, як правило, не вдається відновити ембріогенні потенції культури і одержати рослини-регенеранти.
Етилен - це газ, який добре розчиняється у воді, має характерний запах; хімічна формула Н2С=СН2.
Відкриття етилену як гормону рослин відноситься до кінця XIX - початку XX ст., коли був виявлений вплив этиленового компоненту кам'яновугільного газу на передчасне скидання деревами листків, швидке в'янення квітів, та пригнічення росту пагонів. На початку 30-х років стало відомо, що етилен прискорює дозрівання плодів цитрусових.
Пізніше було встановлено, що стиглі плоди (банани, яблука) виділяють етилен, особливо при пошкодженні.
Дуже маленькі кількості ("сліди") етилену виділяють практично всі органи покритонасіннєвих рослин. У найбільших кількостях [>1 нл/(г.год)] він утворюється у дозріваючих, старіючих тканинах або у відповідь на поранення та інші стресові фактори. Добра розчинність етилену у воді дозволяє йому транспортуватись у водному розчині по рослині.
Етилен, як газ, відрізняється від інших фітогормонів своєю летючістю. 3 цієї причини етилен однієї рослини може впливати на перебіг процесів в іншій рослині, яка знаходиться поряд. Класичний ефект дії етилену спостерігається у плодоовочевих сховищах або при тривалих морських транспортуваннях - перезрілі плоди посилюють дозрівання сусідніх, менш стиглих плодів.
Важливим ефектом фізіологічної дії етилену вважається також стимуляція опадання листків - етилен впливає на розростання прошарку відокремлення, який знаходиться біля основи черешка листка.
Фізіологічна особливість етилену враховується на практиці. За допомогою низьких концентрацій кисню і високих концентрацій вуглекислого газу у середовищі проводять затримку дозрівання плодів при їх тривалому зберіганні. Кисень потрібен для утворення і дії етилену, а вуглекислий газ, навпаки, є його антагоністом. Спеціалісти вважають, що приблизно 100 тис. молекул СО2 здатні протидіяти активності. однієї молекули етилену. В зв'язку з цим суміш газів, яку використовують у сучасних сховищах плодів, обов'язково містить такі кількості СО2, які повністю блокують реалізацію активності ендогенного етилену.
Брасиностероїди
Крім п'яти класичних фітогормонів (ауксини, цитокініни, гібереліни, АБК і етилен) у 1979 році було знайдено і виділено у чистому вигляді з пилку ріпака Brassica napus L новий тип фіторегуляторів, які описані в літературі під назвою брасиностероїди. На сьогодні відомо 12 представників брасиностероїдів, які мають стероїдну природу і відрізняються своїми радикалами (Хрипач В.А. та ш., 1995).
Основна властивість брасиностероїдів - дуже висока фізіологічна активність. Якщо класичні фітогормони діють у концентраціях 10-6 – 10-5 моля, то брасиностероїди - в дозах 10-8-10-11 моля, Після обробки брасиносте-роїдами виявлена сильнодіюча стимуляція росту зернобобових, овочевих. плодових культур. Японські вчені У.Такематцу, А.Такеучі (1989) при обробці пшениці у фазі цвітіння водним розчином одного з брасиностероїдів одержали збільшеення маси колоса і кількості зерна у колосі на 20-30%.
Встановлено, що брасиностероїди сприяють збільшенню розмірів бульб картоплі. Вивчається їх дія на тварин і людину (з хімічної точки зору вони споріднені з гормонами линьки комах - екдистероїдів). Сучасні
експериментальні дослідження свідчать про перспективність використання брасиностероїдів у сільському господарстві (у них невелика токсичність та дуже низькі норми витрат).
С
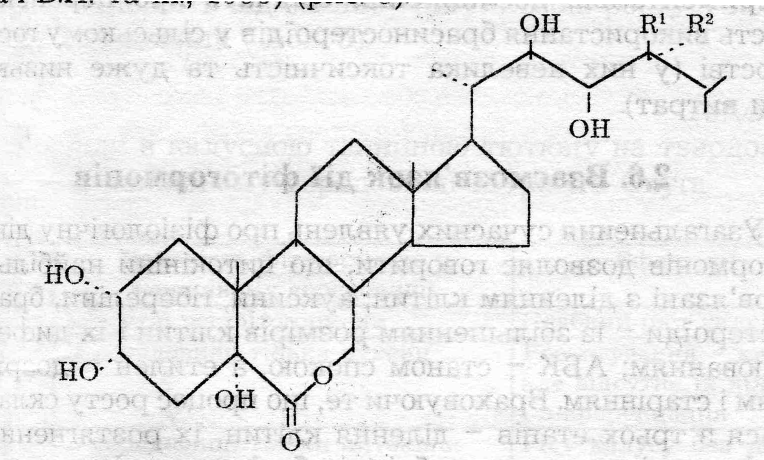 труктуреі формули деяких брасиностероїдів:
труктуреі формули деяких брасиностероїдів:Брасинолід R1 = СН3, R2 = Н
Норбрасинолід R1 = R 2 = Н
Епібрасинолід R1= Н, R2 = СН3
Гомобрасинолід R1 = С2Н5, R 2 = Н
; Доліхолід R1 " R 2 = СН3
Гомодоліхолід R1 - R 2 = СНСН3
Лекція 3.