
Кафедра: "ВТ"
| Вид материала | Доклад |
СодержаниеРис.1. Компоненты нейрона 2.5. Аксон и синапс. Рис.2. Синапс |
- 28-29 октября 2010 г. Девятая международная научно-практическая конференция „Норма, 51.26kb.
- Правительстве Российской Федерации» Кафедра «Макроэкономика» Кафедра «Микроэкономика», 147.21kb.
- План курсовых мероприятий тогирро на 2011 год Содержание Кафедра стратегического менеджмента, 2958.17kb.
- Программа профессиональной переподготовки специалистов для выполнения нового вида профессиональной, 34.84kb.
- Программа производственной преддипломной практики Для студентов Vкурса, обучающихся, 449.19kb.
- Кафедра автоматики и автоматизации производственных процессов, 1002.67kb.
- Кафедра стратегического менеджмента в образовании, 2593.09kb.
- Основной образовательной программы (ооп) высшего профессионального образования бакалавриат, 76.22kb.
- Бакалаврская программа № по направлению психология кафедра, 389.94kb.
- Бакалаврская программа № по направлению психология кафедра, 359.07kb.
2.2. Нейрон.
Нейрон является основным строительным блоком нервной системы. Он является клеткой, подобной всем другим клеткам тела; однако определенные существенные отличия позволяют ему выполнять все вычислительные функции и функции связи внутри мозга.
Как показано рис.1, нейрон состоит из трех частей: тела клетки, дендритов и аксона, каждая часть со своими, но взаимосвязанными функциями.
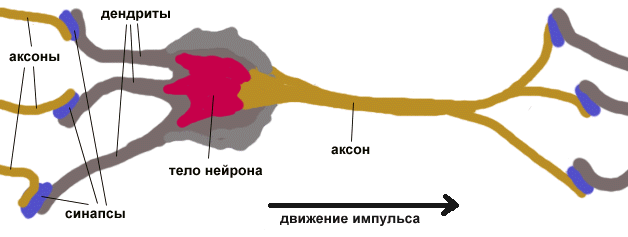
^ Рис.1. Компоненты нейрона
Функционально дендриты получают сигналы от других клеток через контакты, называемые синапсами. Отсюда сигналы проходят в тело клетки, где они суммируются с другими такими же сигналами. Если суммарный сигнал в течение короткого промежутка времени является достаточно большим, клетка возбуждается, вырабатывая в аксоне импульс, который передается на следующие клетки. Несмотря на очевидное упрощение, эта схема функционирования объясняет большинство известных процессов мозга.
Различают мякотные и безмякотные нервные клетки и волокна. Мякотные волокна входят в состав чувствительных и двигательных нервов скелетной мускулатуры и органов чувств. Они покрыты липидной миелиновой оболочкой. Безмякотные волокна составляют основную часть симпатических нервов. Они не имеют миелиновой оболочки и отделены друг от друга клетками нейроглии. Мякотные волокна более "быстродействующи": в таких волокнах диаметром 1 - 3,5 мкм возбуждение распространяется со скоростью 3 - 18 м/c. В безмякотных же волокнах такого же диаметра скорость не превышает 1 - 3 м/c. Это объясняется тем, что проведение импульсов по миелинизированному нерву происходит сальтаторно (скачкообразно). При этом потенциал действия "перескакивает" через участок нерва, покрытый миелином, и в месте перехвата Ранвье (оголенный участок нерва) переходит на оболочку осевого цилиндра нервного волокна. Миелиновая оболочка является хорошим изолятором и исключает передачу возбуждения на соседние, параллельно идущие нервные волокна. В безмякотных волокнах роль изоляторов выполняют клетки нейроглии (опорной нервной ткани). Шванновские нервные клетки - одна из разновидностей глиальных клеток.
Помимо отростков и оболочки, нервные клетки, как и многие другие животные клетки, содержат разнообразные органеллы (ядро, эндоплазматический ретикулум, шероховатый эндоплазматический ретикулум, митохондрии, лизосомы, центриоли, комплекс Гольджи и др.). Они имеют значение для поддержания жизнеобеспечения нервной клетки и ее деления, но практически не участвуют в процессе генерации и преобразования нервных импульсов. Поэтому рассматривать их не будем.
Помимо внутренних нейронов, воспринимающих и преобразующих импульсы, поступающие от других нейронов (через синапсы, рассматриваемые в следующем пункте), еще существуют нейроны, воспринимающие воздействия непосредственно из окружающей их среды (рецепторы), а также нейроны, непосредственно воздействующие на исполнительные органы - эффекторы, например, на мышцы или железы. Если нейрон воздействует на мышцу, его называют моторным нейроном, или мотонейроном. Среди нейрорецепторов различают 5 типов клеток, в зависимости от вида возбудителя:
- фоторецепторы (возбуждаются под действием света и обеспечивают работу органов зрения);
- механорецепторы (реагируют на механические воздействия - располагаются в органах слуха, равновесия; осязательные клетки также являются механорецепторами; некоторые механорецепторы располагаются в мышцах и измеряют степень их растяжения);
- химорецепторы (избирательно реагируют на присутствие или изменение концентрации различных химических веществ, на них основана работа органов обоняния и вкуса);
- терморецепторы (реагируют на изменение температуры либо на ее уровень - холодовые и тепловые рецепторы);
- электрорецепторы (реагируют на токовые импульсы и имеются у некоторых рыб, амфибий и млекопитающих, напр., у утконоса).
2.3. Тело ячейки.
Нейроны в мозгу взрослого человека не восстанавливаются; они отмирают. Это означает, что все компоненты должны непрерывно заменяться, а материалы обновляться по мере необходимости. Большинство этих процессов происходит в теле клетки, где изменение химических факторов приводит к большим изменениям сложных молекул. Кроме этого, тело клетки управляет расходом энергии нейрона и регулирует множество других процессов в клетке. Внешняя мембрана тела клетки нейрона имеет уникальную способность генерировать нервные импульсы (потенциалы действия), являющиеся жизненными функциями нервной системы и центром ее вычислительных способностей.
2.4. Дендриты.
Большинство входных сигналов от других нейронов попадают в клетку через дендриты, представляющие собой густо ветвящуюся структуру, исходящую от тела клетки. На дендритах располагаются синаптические соединения, которые получают сигналы от других аксонов. Кроме этого, существует огромное количество синаптических связей от аксона к аксону, от аксона к телу клетки и от дендрита к дендриту; их функции не очень ясны, но они слишком широко распространены, чтобы не считаться с ними.
В отличие от электрических цепей, синаптические контакты обычно не являются физическими или электрическими соединениями. Вместо этого имеется узкое пространство, называемое синоптической щелью, отделяющее дендрит от передающего аксона. Специальные химические вещества, выбрасываемые аксоном в синаптическую щель, диффундируют к дендриту. Эти химические вещества, называемые нейротрансмиттерами, улавливаются специальными рецепторами на дендрите и внедряются в тело клетки.
Определено более 30 видов нейротрансмиттеров. Некоторые из них являются возбуждающими и стремятся вызывать возбуждение клетки и выработать выходной импульс. Другие являются тормозящими и стремятся подавить такой импульс. Тело клетки суммирует сигналы, полученные от дендритов, и если их результирующий сигнал выше порогового значения, вырабатывается импульс, проходящий по аксону к другим нейронам.
^ 2.5. Аксон и синапс.
Аксон может быть как коротким (0,1 мм), так и превышать длину 1 м, распространяясь в другую часть тела человека. На конце аксон имеет множество ветвей, каждая из которых завершается синапсом, откуда сигнал передается в другие нейроны через дендриты, а в некоторых случаях прямо в тело клетки. Таким образом, всего один нейрон может генерировать импульс, который возбуждает или затормаживает сотни или тысячи других нейронов, каждый из которых, в свою очередь, через свои дендриты может воздействовать на сотни или тысячи других нейронов. Таким образом, эта высокая степень связанности, а не функциональная сложность самого нейрона, обеспечивает нейрону его вычислительную мощность.
Синаптическая связь, завершающая ветвь аксона, представляет собой маленькие утолщения, содержащие сферические структуры диаметром 30..60 нм, называемые синоптическими пузырьками, каждый из которых содержит большое число нейротрансмиттерных молекул.
Вообще, в состав синапса входят пресинаптическая, постсинаптическая мембраны и синаптическая щель.
Различают медиаторные (химические) и безмедиаторные (электрические) синапсы. Во всех этих случаях импульсы передаются посредством деполяризующего действия электрического тока, который генерируется в пресинаптическом элементе. В случае электрических синапсов возможна передача импульсов как в одном, так и в двух направлениях. Есть также синапсы, в которых сочетаются электрическая и химическая передача импульсов (например, в огромных чашевидных синапсах цилиарного ганглия цыпленка).
Принцип функционирования медиаторного синапса состоит в следующем. Импульс возбуждения, достигший пресинаптического элемента (дендрита или аксона), вызывает нейросекрецию биологически активного вещества - медиатора (посредника) в синаптическую щель. В качестве медиатора могут выступать такие вещества, как ацетилхолин, норадреналин, адреналин; реже - G-аминомасляная кислота, серотонин, нейропептиды. Токи действия, вызываемые пресинаптическими импульсами, не оказывают заметного влияния на постсинаптическую мембрану, но приводят к разрушению оболочки пузырьков с медиатором. При этом мельчайшие порции (кванты) медиатора поступают через отверстия в пресинаптической мембране в синаптическую щель, соединяются с белковым (холинорецептивным) веществом, где поляризуют постсинаптическую мембрану и одновременно превращают особые рецептивные участки постсинаптической мембраны в ячеистую структуру, что приводит к ее кратковременной избирательной проницаемости для разных ионов. Так, при возбуждении поры становятся больше, и увеличивается их проницаемость для всех гидратированных ионов независимо от их размера (в том числе и натрия Na+). При этом генерируется возбуждающий постсинаптический потенциал (ВПСП), а в качестве медиатора выступает ацетилхолин. При торможении поры уменьшаются и пропускают лишь ионы небольшого размера, например, калия (K+) и кальция (Ca2+). При этом генерируется тормозной постсинаптический потенциал (ТПСП), а в качестве нейромедиатора обычно выступает G-аминомасляная кислота или глицин. Как же происходит транспорт ионов при возбуждении и торможении и каким образом он влияет на появление возбуждающего или тормозного потенциала?
Рассмотрим одну из гипотез. В состоянии покоя между постсинаптической мембраной и окружающей нейрон средой существует определенная разность потенциалов (около 0,08 В). Это объясняется тем, что в состоянии покоя внутри постсинаптической клетки (ПСК) повышена концентрация ионов калия K+; концентрация же ионов калия с внешней стороны постсинаптической клетки невысока, но здесь высока концентрация ионов натрия (Na+) и хлора (Cl - ). В состоянии покоя у ПСК каналы пропускают ионы лишь небольшого размера, поэтому ионы калия начинают выходить наружу, унося с собой и положительный заряд. Это приводит к появлению разности потенциалов покоя (или просто мембранного потенциала покоя), причем эндоплазма в этом случае заряжается отрицательно, а вблизи наружной стороны мембраны ПСК скапливается положительный заряд. В случае, если в состоянии покоя удается снизить мембранный потенциал на некоторую величину, это приводит к тому, что каналы начинают пропускать довольно крупные ионы натрия, т.е. каналы переходят в состояние возбуждения (объяснение этого см. выше). В случае возбуждения, ионы натрия из внешней среды через открытые каналы начинают проникать внутрь ПСК. Возникает скачок постсинаптического ВПСП-потенциала. Процесс возбуждения длится очень короткое время и сопровождается диффузией ацетилхолина. Для того, чтобы клетка возбудилась, ее потенциал должен быть снижен до определенного значения, называемого порогом возбуждения.
В случае торможения, наоборот, мембранный ПСК-потенциал увеличивается по абсолютному значению (эндоплазма приобретает больший отрицательный заряд), так как в этом случае поры постсинаптической и пресинаптической мембраны уменьшаются, ионы калия начинают выходить наружу, в результате чего снижается мембранный потенциал.
В перерывах между отдельными импульсами возбуждения ацетилхолин разлагается ферментом холинэстеразной на холин и уксусную кислоту.
Весь выше рассмотренный процесс моно увидеть на рис.2.
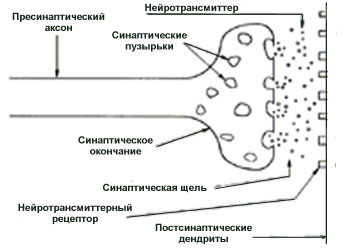
^ Рис.2. Синапс
Кроме распространения бинарного сигнала, обеспечиваемого возбуждением первого импульса, в нейронах при слабой стимуляции могут также распространяться электрохимические сигналы с последовательной реакцией. Локальные по своей природе, эти сигналы быстро затухают с удалением от места возбуждения, если не будут усилены. Природа использует это свойство первых клеток путем создания вокруг аксонов изолирующей оболочки из шванковских клеток. Эта оболочка, называемая миелиновой, прерывается приблизительно через каждый миллиметр вдоль аксона узкими разрывами, называемыми узлами, или перехватами Ранвье. Нервные импульсы, приходящие в аксон, передаются скачкообразно от узла к узлу. Таким образом, аксону нет нужды расходовать энергию для поддержания своего химического градиента по всей своей длине. Только оставшиеся неизолированными перехваты Ранвье являются объектом генерации первого импульса; для передачи сигнала от узла к узлу более эффективными являются градуальные реакции. Кроме этого свойства оболочки, обеспечивающего сохранение энергии, известны ее другие свойства. Например, миелинизированные нервные окончания передают сигналы значительно быстрее немиелинизированных. Обнаружено, что некоторые болезни приводят к ухудшению этой изоляции, что, по-видимому, является причиной других болезней.