
М. В. Ломоносова биологический факультет н а правах рукописи Столяров Андрей Павлович Структурно-функциональная организация эстуарных экосистем Белого моря 03. 02. 10 гидробиология Автореферат
| Вид материала | Автореферат |
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- М. В. Ломоносова биологический факультет на правах рукописи никольская кира Алексеевна, 1297.21kb.
- Структурно-функциональная организация палеоамигдалы: фундаментальные закономерности, 2391.63kb.
- Юров иван юрьевич структурно-функциональная организация хромосом при нервно-психических, 850.19kb.
- Лекция № клетка элементарная биологическая система. Структурно функциональная организация, 341.33kb.
- Автореферат диссертации на соискание ученой степени, 1241.4kb.
- На правах рукописи, 470.03kb.
- На правах рукописи, 1724.93kb.
- На правах рукописи, 378.58kb.
- На правах рукописи, 594.43kb.
Отдельные относительно самостоятельные структуры эстуария
Наиболее важными отдельными структурами эстуарных экосистем, обеспечивающими их нормальное функционирование и целостность, являются соленый марш – область первичного продуцирования органического вещества и мидиевая банка – область биофильтрации и преобразования низкокачественного детрита во вторичную продукцию детритофагов, сестонофагов и их прижизненные выделения (фекалии и псевдофекалии).
Мидиевые банки играют важную роль в трансформации органического вещества и энергии. Располагаясь в зонах активной гидродинамики (чаще всего на выходе из эстуариев) (Daurer, 1993; Meire et. al., 1991; Shillabeer, Tapp, 1990), они играют огромную роль в осаждении взвеси и очищении от нее поверхност-ной (прибрежной) воды (Dame, 1993).
Сообщество мидиевой банки имеет выраженную поясную структуру с преобладанием в нижней литорали и сублиторали крупных фильтраторов Mytilus edulis, в средней – более мелких детритофагов макомы и гидробии, а в верхней – еще более мелких олигохет и той же гидробии. Mytilus edulis состав-ляет 80-90% всей биомассы сообщества нижней и 30-50% средней литорали. Соответственно, число видов, общая плотность и общая биомасса макрозообентоса в районе мидиевой банки были максимальными в нижнем и среднем горизонте прибрежно-отливной зоны, резко падая к верхней части осушной полосы.
В скоплениях мидии основная функциональная роль в сообществе макрозообентоса принадлежит сестонофагу Mytilus edulis, составляющему 94% (в июне) и 89% (в сентябре) биомассы всего сообщества. В промежутках между щетками в биотопе чистого песка роль основного доминанта в сообществе переходит к детритоядному эвригалинному брюхоногому моллюску Hydrobia ulvae, составляющему около 50% плотности и биомассы всего сообщества. Видовое разнообразие и выравненность сообщества макрозообентоса (рассчи-танные на основе биомассы) были более низкими в скоплениях мидии нежели между ними, что обусловлено резким доминированием M. edulis по биомассе в мидиевых щетках и H.ulvae по плотности видовых популяций в биотопе чистого песка. Несмотря на то что в средней и нижней литорали преобладают пески различной зернистости с относительно бедным содержанием органического ве-щества, обитающие здесь популяции детритофагов (в основном макома и гид-робия) образуют достаточно плотные скопления между щетками мидий, что обусловлено доступностью и постоянной возобновляемостью ресурсов качественного детрита за счет жизнедеятельности мидий и образования ими фекалий и псевдофекалий, которые легко усваиваются обитающими здесь организмами (Столяров, Бурковский, 2006).
Соленые марши - прибрежные территории, постоянно заливаемые морской водой и покрытые густой растительностью. Они весьма типичны для литорали опресняемых губ и эстуариев и не только определяют облик животного населения на соответствующей территории, но и оказывают заметное влияние на многие биогеохимические процесссы в прибрежной экосистеме.
Соленый марш представляет собой довольно сложно организованный и вполне самостоятельный компонент эстуарной прибрежной экосистемы. Ведущая роль принадлежит высшей растительности, которая является здесь не только основным продуцентом (роль микрофитобентоса незначительна из-за отсутствия в плотном дерновистом грунте условий для его развития), но и наиболее важным косвенным фактором, определяющим качественный и количественный состав беспозвоночных, особенно в конце лета, в период своего максимального развития.
В пределах соленого марша, занимающего большую часть верхней литорали, четко выделяются два подгоризонта, различающихся характеристиками среды и видовым составом организмов. В верхнем подгоризонте преобладают растения, обычно образующие плотную дерновину, препятствующую развитию здесь роющих беспозвоночных. В нижнем подгоризонте среди разреженного растительного покрова довольно обильны роющие животные, характерные и для остальной части литорали. В целом распределение растений имеет более выраженный зональный характер, чем распределение беспозвоночных. В пределах подгоризонтов распределение большинства видов растений и животных носит случайно-мозаичный характер, в общем соответствующий локальному распределению факторов среды (Бурковский, Столяров, 2001).
3.1.2. Уровень мейобентоса
Видовой состав и разнообразие сообщества
Учет групп мейобентоса проводили в основном до крупных таксонов (Nematoda, Ostracoda, Harpacticoida, Turbellaria, Halacarida). Исключение составляла группа псевдомейобентоса (личинки беспозвоночных), определение которых проводилось до вида. Также в отдельные годы изучались группы нематод и гарпактикоидных копепод (1995, 2000 гг.) (Удалов, Бурковский, 2002; Чертопруд, 2005; Столяров, Бурковский, 2008).
Большая часть нематод была представлена морскими эвригалинными видами, встречающимися на всем протяжении эстуария. Пресноводные виды редки и встречаются спорадически в кутовой части эстуария. Видовое богатство и разнообразие сообщества нематод слегка снижаются в сторону реки. При этом существенного обеднения видового состава нематод в опресненном районе эстуария не происходит.
Видовое разнообразие гарпактикоидных копепод гораздо ниже, чем нематод, и убывает с уменьшением солености воды от моря к реке. Все виды можно разделить на морские эвригалинные, предпочитающие мористый район эстуария, солоноватоводные и морские эвригалинные, распространенные по всему эстуарию, но предпочитающие центральные районы, и некоторые солоноватоводные и пресноводные, встречающиеся в кутовом районе эстуария. Пресноводные и солоноватоводные виды малочисленны и обитают преимущественно в опресненном районе эстуария при низкой солености, а морские эвригалинные - характеризуются высокой плотностью популяций и предпочитают мористый район, воды которого имеют более высокую соленость.
Интегральные характеристики сообщества
Наиболее существенный вклад в общую плотность сообщества мейобентоса вносят нематоды (в среднем 50-80%), гарпактикоидные копеподы находятся в субдоминантах и в среднем составляют до 10 % от общей плотности всего мейобентоса. В то время как наибольший процент от общей биомассы сообщества составляли остракодовые рачки и псевдомейобентос.
Общие показатели структуры сообщества мейобентоса (общая плотность, биомасса и разнообразие), в отличие от макробентосных характеристик (Столяров, Бурковский, 2004, 2005), имеют тенденцию к понижению значений в сторону мористой части эстуария (по мере повышения солености воды), где расположена обширная мидиевая банка, и повышению − от нижних горизонтов литорали к верхним. Соответственно опресненная зона эстуария характеризуется повышенной плотностью организмов мейобентоса (рис.4).
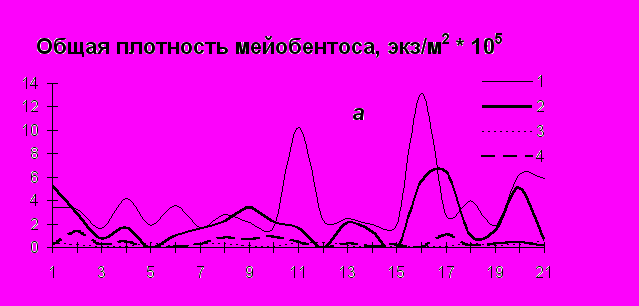
Р
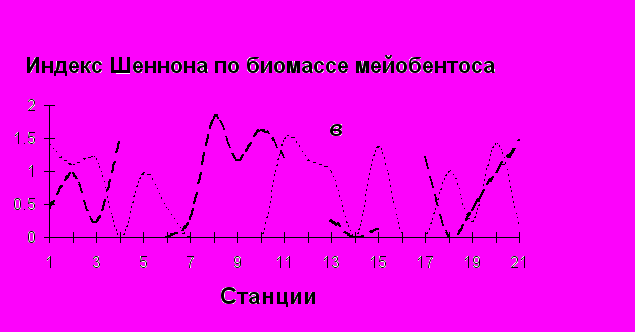

ис. 4. Изменение общей плотности (а), общей биомассы (б) и видового разнообразия (в) организмов мейобентоса в эстуарии р. Черной по мере увеличения солености воды. 1 – организмы эвмейобентоса в июне, 2 – организмы эвмейобентоса в сентябре, 3 – ораганизмы псевдомейобентоса в июне, 4 – организмы псевдомейобентоса в сентябре. Станции и разрезы расположены по мере увеличения солености воды. Станции 1 -7 – нижняя литораль, 8-14 – средняя литораль, 15-21 – верхняя литораль.
В целом мейобентосные животные преобладали на илистых грунтах эстуария с повышенным содержанием органических веществ. Причем, в опресненном районе, где процент алевропелита выше, общая плотность организмов мейобентоса (эвмейобентоса и личинок беспозвоночных) тоже была выше. В вертикальном направлении (по отношению к мареграфическому уровню) численность мейобентосных животных увеличивалась от нижней литорали в сторону верхней, что связано с повышением заиленности грунта в верхних горизонтах литорали и, как следствие, концентрацией там органических веществ. Надо отметить, что биомасса псевдомейобентоса в июне в начале сезонной сукцессии была выше в мористом районе эстуария, а в сентябре – в опресненном (значительная плотность молоди макомы).
При сравнении биомассы различных групп эвмейобентоса мористого и опресненного участка эстуария было установлено, что в июне доли остракодо-вых рачков и гарпактикоидных копепод в общей биомассе сообщества мейо-бентоса были выше в опресненном районе, а также в верхней литорали по всему эстуарию. В сентябре эта тенденция сохранялась для остракод, а группа гарпактицид имела обратную тенденцию – снижения доли в общей биомассе сообщества мейобентоса в опресненном районе. Нематоды характеризуются повышенной плотностью организмов в заиленных местообитаниях (в опреснен-ном районе и верхней литорали), однако доля этой группы в общей биомассе эвмейобентосных животных снижается в сторону опресненных районов (осо-бенно харатерно для нижней литорали). Таким образом, по мере уменьшения солености воды происходит изменение соотношения крупных таксонов эвмейо-бентосных организмов с нематодно-гарпактикоидной на остракодно-нематод-ную с границей между ними около 8-10‰ (в районе верхних порогов). Что касается личинок беспозвоночных (псевдомейобентос), то и в начале (в июне) и в конце (в сентябре) сезонной сукцессии их плотность была выше в верхнем горизонте приливно-отливной полосы (особенно заметно в конце сезона). В на-чале сезонной сукцессии в верхней литорали опресненного района личинки беспозвоночных редки и встречаются спорадически, что объясняется неблагоп-риятными условиями жизни для большинства представителей этой группы мейобентоса – существенным опреснением (условия обитания приближаются к пресноводным). Отметим также, что в конце сезонной сукцессии в сентябре, несмотря на общую низкую плотность организмов псевдомейобентоса в ниж-ней литорали по сравнению с вышерасположенными участками, наблюдались относительно высокие показатели их биомассы, что связано с повышенной плотностью молоди макомы в нижних горизонтах приливно-отливной полосы.
Сходство и различие локальных сообществ
При классификации локальных сообществ по биомассе крупных таксонов эвмейобентоса (нематоды, остракоды, гарпактикоидные копеподы, галакариды и турбелярии) и видовых популяций псевдомейобентоса можно выделить со-общества преимущественно верхней (и средней) литорали (18-21) и сообщества нижней литорали (и средней) (6-9), расположенные по всему эстуарию, сообщества опресненного района эстуария (1, 7, 16, 17) и сообщества верхней и средней литорали, расположенные в районе верхних порогов между опресненн
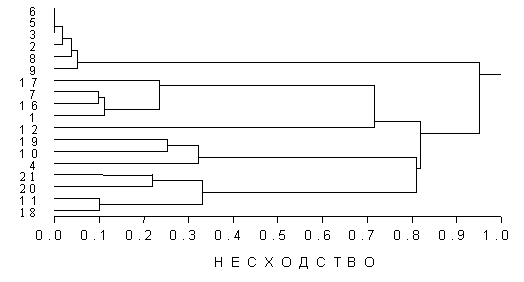
ой и мористой областью эстуария (4, 10, 19) (рис. 5).
Рис. 5. Дендрограмма сходства локальных сообществ мейобентоса в эстуарии. Станции 1 -7 - нижняя, 8-14 – средняя, 15-21 – верхняя литораль.
При этом наименьшее сходство видовой структуры мейобентоса демонстрируют станции верхней литорали (18-21), приуроченные к участкам умеренной гидродинамики (верхние и нижние пороги, мыс песчаный), а наибольшее − станции нижней (6-9), расположенные в районах низкой гидродинамики (в середине опресненного и мористого водоемов с илистыми осадками). Таким образом, хорошо различаются сообщества верхней и нижней литорали; опресненного, переходного и мористого районов эстуария.
Размерная структура сообщества мейобентоса
Для эвмейобентосных организмов характерно некоторое уменьшение средних размеров по мере заиления осадков (и увеличения кладофоры, зостеры, солончаковых растений) к серединам солоноватой и солоноватоморской зонам и увеличение размеров к узостям эстуария с песчанистыми грунтами. В основном такая тенденция изменения размеров связана с закономерностями распределения наиболее многочисленной группы мейобентоса – класса нематод, которые в заиленных местообитаниях имели более мелкие формы. Остракоды также имели меньшие размеры в заиленных местообитаниях, в то время как гарпактикоидные копеподы и особенно турбелярии с увеличением доли алевропелита становятся более крупными. Распределение эвмейобентоса меньше зависит от градиента солености и в основном связано с характеристиками грунта, размером зерна, содержанием доступного органического вещества; биомасса и обилие этой группы были выше в опресненном районе, осадки которых более заиленные, а процент алевропелита и органического вещества выше.
Индивидуальные веса организмов псевдомейобентоса были значительно выше в опресненном районе эстуария в конце сезонной сукцессии. В опресненном районе преобладали личинки и молодь солоноватоводных олигохет и хирономид, а в солоноватоморском – морских эвригалинных видов: Macoma bal-thica, Pygospio elegans, Fabricia sabella. При этом Macoma balthica в течение се-зона все более осваивает опресненный район эстуария.
3.1.3. Уровень микробентоса
Видовой состав и распределение сообщества инфузорий
Всего на изучаемых участках эстуария реки Черной обнаружено 124 вида инфузорий: в морском районе (ст.1) – 100, в солоноватоморском (ст. 2 и 3) – 94-87, в солоноватом (ст. 4 и 5) – 72- 59 видов. С уменьшением солености воды и удаленности от моря количество видов заметно снижается. Ежегодно основу сообщества инфузорий в эстуарии составляют 23 массовых и 37 второстепенных видов, что составляет около половины всего видового богатства сообщества.
Локальные сообщества, формирующиеся в разных зонах эстуария, представляют собой различные варианты комбинаций морских и солоноватоводных видов, отвечающие местным условиям, – в первую очередь режиму солености. Можно выделить 3 основные группы: 1) морские эвригалинные виды, развива-ющиеся при средней солености выше 14 – 16 ‰ (указана соленость над грунтом), но способные выдерживать непродолжительное опреснение до 3-5 ‰; 2) - морские более эвригалинные виды, развивающиеся при средней солености выше 8 – 10 ‰, но выдерживающие кратковременное понижение ее до 0 – 2 ‰; 3) – солоноватоводные и морские эвригалинные виды, предпочитающие обитать при средней солености 4 – 8 (10) ‰, но выдерживающие как продолжительное ее снижение до 0 ‰, так и повышение до 15 ‰ (некоторые до 18 – 20 ‰).
Интегральные характеристики структуры сообщества
С уменьшением солености заметно уменьшается сложность эстуарного сообщества: снижаются показатели видового разнообразия, общей плотности и биомассы инфузорий. Наиболее простые и изменчивые сообщества формируются в опресненной зоне эстуария, где наблюдаются значительные приливно-отливные колебания солености при средних значениях 3-8 ‰, находящиеся на грани приспособительных возможностей как морских, так и пресноводных организмов (Хлебович, 1974).
Сходство и различие локальных сообществ (станций)
Наибольшее сходство демонстрируют сообщества инфузорий, расположенные в мористом (ст.1 и ст. 2) и опреснном районах (ст.4 и ст. 5), а наименьшее – между ними (ст.2 – ст.4) (рис. 6). Это свидетельствует, что основные изменения в структуре эстуарного сообщества инфузорий происходят в промежуточной зоне – зоне наибольшего варьирования солености воды.

Рис. 6. Дендрограмма кластерного анализа сообщества инфузорий в эстуарии.
Размерная структура инфузорного микробентоса
Вдоль продольной оси эстуария от реки к морю происходит увеличение численности и биомассы инфузорий большинства размерных классов (РК), обусловленное увеличением солености воды и Eh осадка. Каждый размерный класс (РК) включал организмы определенных размеров. Например, РК 0.25 образован организмами, размеры которых менее 0.25х103 мкм3, РК0.5 – размерами от 0.25х103 до 0.5х103 мкм3, РК1 – от 0.5х103 до 1х103 мкм3 и т.д. Связь относительного обилия РК в сообществе инфузорий с абиотическими факторами, как правило, обусловлена особенностями распределения отдель-ных видов и является, по-видимому, суммарным результатом их индиви-дуальной реакции на изменения условий среды обитания. При этом обилия инфузорий связаны как с соленостью, так и с гранулометрическим составом грунта. Мелкие виды убиквисты обильно развиваются на всем протяжении эстуария и не проявляют сильных связей с факторами среды. Наиболее крупные инфузории, напротив, проявляют весьма сильную зависимость от гранулометрического состава грунта, соленость на них оказывает меньшее влияние.
Анализ изменений размерных спектров по биомассе (основной вклад в которые вносят крупные формы) и численности (которые формируются мелкими видами) вдоль продольной оси эстуария позволяет выявить некоторые особенности распределения мелких и крупных инфузорий. Размерные спектры по биомассе образованы весьма большим числом видов крупных размеров, сменяющих друг друга на всем протяжении эстуария. Значительная пространственно-временная изменчивость факторов среды в эстуарии обуславливает появление большого количества массовых видов, развивающихся в разных точках пространства-времени, характеризующихся конкретными условиями. Высокое разнообразие крупных инфузорий в эстуарии обусловлено, по-видимому, большей чувствительностью их к изменяющимся условиям среды с узкими диапазонами толерантности по сравнению с мелкими организмами, что приводит к развитию многих видов инфузорий, адаптированных к конкретным локальным наборам факторов. Размерные спектры по данным численности, напротив, образованы небольшим числом мелких видов-убиквистов, обильно развивающихся на всем протяжении эстуария. При этом наиболее мелкие инфузории (Uronema marina (РК4)) обильны на всех станциях. Состав же более крупных размерных классов четко различается между мористой (ст. 1 и 2) и опресненной (ст. 3-5) областями эстуария.
Наблюдаемое уменьшение видового богатства сообщества инфузорий в опресненной части эстуария происходит только за счет двух размерных классов (РК16 и РК64). Инфузории этих размеров формируют основное видовое разнообразие специфической интерстициальной фауны в мористой части эстуария. Это морские плейоэвригалинные организмы, не выдерживаю-щие значительного опреснения.
Сравнительный анализ структуры сообществ микро-, мейо-, и макробентоса
Распределение разных таксономических и размерных групп эстуарных организмов определяется в основном факторами среды и носит комплемен-тарный характер.
Выделяются два комплекса факторов, определяющих структуру сооб-ществ микро-, мейо- и макробентоса (табл. 2).
Таблица 2. Вклады факторов в компоненты изменчивости среды
-
Абиотические факторы:
К1
К2
Фракции грунта < 0.1 мм (%)
-0.369
0.859
Содержание органического вещества в грунте (%)
-0.235
0.947
Характер грунта (1- песок, 2 – илистый песок, 3- песчанистый ил, 4 - ил)
-0.308
0.743
pH в поверхностном слое грунта
-0.132
0.040
Eh в поверхностном слое грунта (мв)
0.755
-0.621
Прозрачность воды (см)
0.918
-0.302
Соленость (‰), малая вода
0.937
-0.311
Соленость (‰), полная вода
0.976
-0.209
Процент объясненной дисперсии
40
37
Примечание. К1 и K2 – главные компоненты изменчивости среды.
Первая группа факторов включает соленость и прозрачность воды, окис-лительно-восстановительный потенциал (их показатели понижаются от моря к реке), вторая группа - различные характеристики осадка, содержание в нем алевропелита и органического вещества (процент которых выше в осадках оп-ресненного района эстуария).
В соответствии с ними биомасса инфузорий, жгутиковых и макрозо-обентоса уменьшается от моря к реке, а мейобентоса, микрофитобентоса и бак-терий увеличивается (рис. 7).

Рис. 7. Распределения биомасс размерно-таксономических групп бентосных сообществ вдоль продольной оси эстуария на градиенте солености воды. 1 – 5: станции. Ст. 1: 14 - 21 ‰, ст. 2: 12-19 ‰, ст. 3: 7 – 12 ‰, ст. 4: 5 – 10 ‰, ст. 5:
0 – 7 ‰.
Макробентос наиболее сильно реагирует на изменения солености воды и коррелирует с ней (r=0.82). С уменьшением солености происходит уменьшение видового разнообразия, общей биомассы сообщества и среднего размера орга-низмов при увеличении их общей плотности; исчезают крупные формы макро-бентоса (мидии, мии, литторины), а доминирующее положение занимают мелкие организмы с индивидуальной массой около 1 мг (в основном олигохеты и хирономиды).
Мейобентосные организмы (включая и личинок беспозвоночных) дости-гают значительной плотности и биомассы в опресненном районе эстуария; прямой зависимости их структуры от солености не наблюдается. Гораздо боль-шее влияние на организмы этой группы оказывают тип грунта и содержание в нем органического вещества (r=0.8). Изменения численности и биомассы инфу-зорий (основной группы микробентосных животных) значительно коррелирует с соленостью (r=0.91). С уменьшением солености видовое разнообразие, плот-ность и биомасса сообщества инфузорий снижаются.
Полученные данные позволяют говорить о комплементарном (взаимодополняющем) характере распределения организмов микро-, мейо- и макробен-тоса по эстуарию.
Одной из причин комплементарности распределения организмов микро- и мейобентоса является заиление грунта, которое приводит к уменьшению крупных форм микробентоса, обитающих в интерстициальных пространствах, и преобладнию мелких организмов. Свободная размерная ниша на подобном субстрате оказывается занятой представителями мейобентоса. Большое количе-ство органического вещества в осадках эстуария, низкая и изменчивая соле-ность воды способствуют развитию мелких видов детритофагов-собирателей, относящихся к мейо- и макробентосу (с индивидуальной массой около 1 мг сы-рой массы). И если в мористом районе доля крупных беспозвоночных живот-ных (мидии, макомы, мии) составляет 80-90 %, то в опресненном устьевом районе она снижается до 0.7 %. Как следствие, видим комплементарный (до-полняющий) характер распределения организмов разных размерных групп.
3.2. Пространственно-временные изменения структуры сообщества
эстуария
Ведущим фактором сезонных и межгодовых изменений структуры эстуарного макробентоса является солевой и температурный режимы водоема, контролирующие развитие и распределение видов по акватории эстуария.
Основное направление сезонных и межгодовых изменений в структуре сообществ обусловлено преимущественно режимом солености. С ее увеличением морские эвригалинные виды все глубже проникают в кут эстуария, а солоноватоводные и пресноводные виды, наоборот, сокращают ареал своего распространения, смещаясь в сторону реки. В связи с этим растут доли и области распространения морских видов, общий уровень их интеграции, стирание границ между основными зонами эстуария.
Наивысшая интеграция видов в сообществе (минимум групп при макси-мально возможной связанности видов в них) приходится на периоды повы-шения средней солености воды в эстуарии. Наивысшая дифференциация (много групп и низкая связанность видов в них) свойственна периодам распреснения эстуария, когда создаются условия для развития комплексов солоноватоводных и пресноводных видов.
Составляющими такого процесса являются миграции видов в постепенно осолоняемые (к концу сезона и в ряду лет) внутренние воды эстуария, а также рост личинок и молоди предыдущих лет до учитываемых размеров.
3.3. Жизненные циклы и стратегии, формирование структуры эстуарного сообщества
Ключевым процессом в поддержании видовой структуры сообщества эстуария является пополнение его за счет новообразующейся молоди.
В планктоне и бентосе были обнаружены личинки 33 видов беспозвоноч-ных животных, из которых 9 – типичные литоральные животные, 16 – общие для литорали и сублиторали, 8 – преимущественно сублиторальные.
Определяющими этапами процесса формирования сообщества являются нерест, рост, развитие, оседание и ассимиляция личинок, сопровождающиеся соответствующей смертностью.
В эстуарии преобладают короткоживущие солоноватоводные и морские эвригалинные виды макробентоса (1-2 года) с растянутым нерестом и развитием личинок в кладках, больше отвечающие требованиям r-жизненной стратегии. На выходе из эстуария в зоне активной гидродинамики преобладают крупные морские менее эвригалинные виды беспозоночных животных (в основном популяции фильтраторов Mytilus edulis) с большей продолжительностью жизни (10 лет и более) со сжатым нерестом и пелагической личинкой, в ряду r - K стратегий ближе к K-жизненной стратегии по сравнению с предыдущей группой. Оппортунистические мелкие виды оказываются лучше адаптированы к условиям непредсказуемо меняющейся среды эстуария (пониженная и меняющаяся соленость воды, повышенное содержание органических веществ в осадках и т.д.) за счет быстрого роста и созревания, большей защищенности личинок, высокого репродуктивного потенциала (табл. 3).
Т
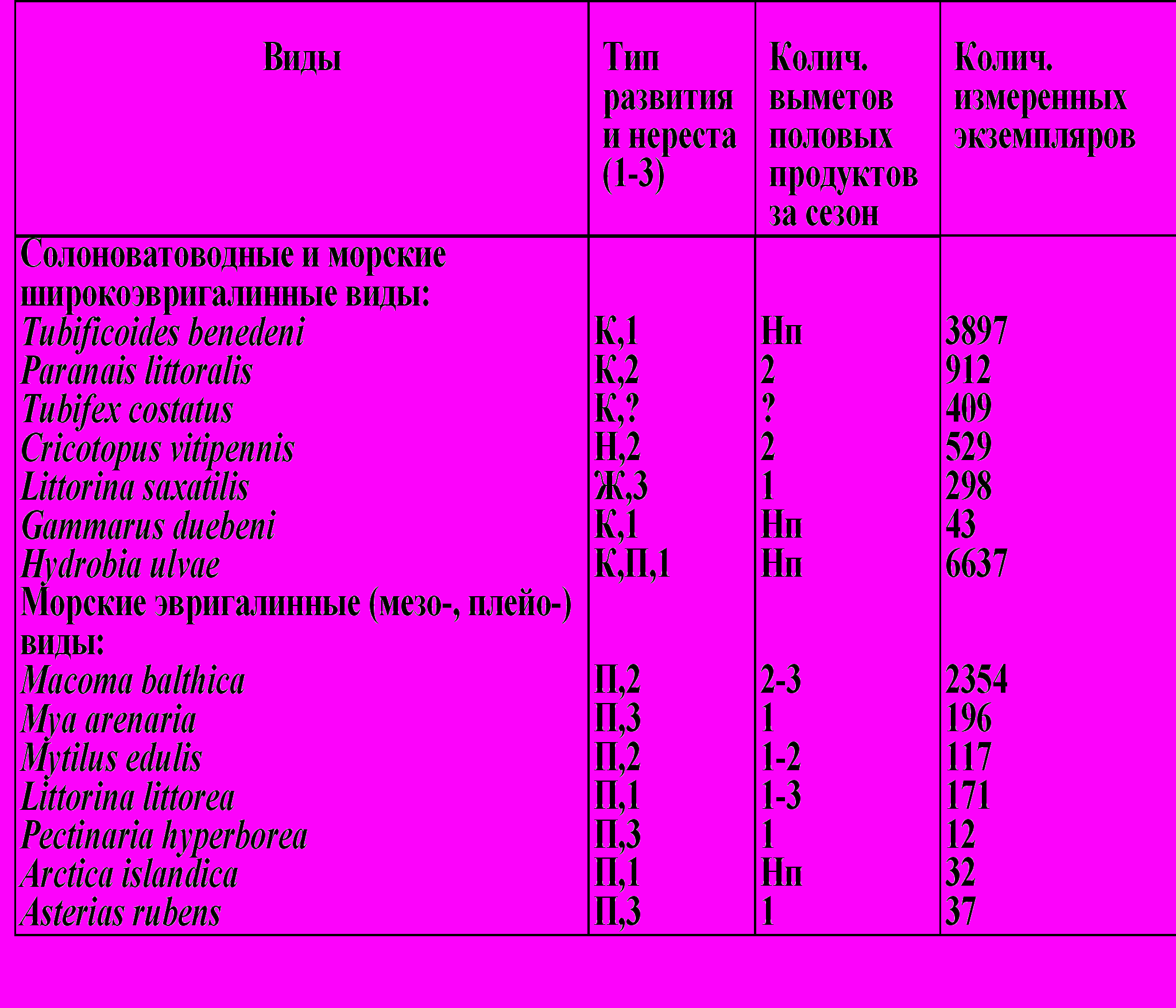
аблица 3. Типы развития и интенсивность нереста
Примечание. К – кладка во внешней среде, Д – кладка в домике, Ж – живорождение, Н – развитие во внешней среде, П – развитие с пелагической личинкой; 1 – продолжительный нерест без выраженных ритмов, 2 – продолжительный ритмичный нерест, 3 – сжатый нерест. Нп – неправильный нерест (или множество выметов).
Ранние стадии онтогенеза – наиболее уязвимое звено в развитии эстуарных беспозвоночных; максимальная смертность пелагических личинок наблюдается в толще воды и в процессе их оседания на грунт, у бентосных – в начале освоения ими биотопа. Смертность личинок и сеголеток у видов с пелагической личинкой существенно выше, чем у видов с живорождением или прямым развитием. Жесткие условия эстуария (особенно на литорали) требуют от взрослых организмов большей заботы о своем потомстве, что достигается развитием личинок в защищенных кладках, в коконах, в слинявшем панцире, внутри трубок взрослых особей, на плеоподах или внутри материнского организма.
В значительной мере судьба личинок зависит от конкретного биоценотического окружения. Скопления того или другого вида указывают, как правило, на соответствующие благоприятные условия, поддерживающие его популяцию. Поэтому личинки, находящиеся среди взрослых особей, имеют больше шансов выжить и вырасти. Бентосные личинки с момента появления (выхода из кладок или рождения) оказываются максимально приближенными к своим родителям и, таким образом, изначально осваивают пространство наиболее благоприятных факторов. В отличие от бентоса, в структуре планктонного пула (адаптированной к условиям пелагиали) не предопределена ближайшая судьба осевших личинок; она целиком зависит от процессов, протекающих на дне водоема. Пул пелагических личинок в данном случае выступает как "слепой" донор. Большинство пелагических личинок по нашим наблюдениям (личинки пескожила, гидробии, мии, макомы, спио) в основном оседают за границами массовых поселений взрослых организмов. И только позднее, перемещаясь с приливно-отливными и ветровыми течениями, некоторые из них достигают благоприятных мест обитания. Таким образом, идентичность структуры ювенильного и взрослого макробентоса достигается раньше в группе видов, имеющих бентосную личинку, позже – у видов с пелагической личинкой. Исходная пространственная близость бентосных личинок к родителям определяет более быстрое продвижение их в направлении структуры взрослых организмов.
3.4. Функциональная организация эстуарных экосистем
Трофические группы и их распределение по эстуарию
В эстуарных экосистемах из-за их относительной мелководности, малой прозрачности вод и преобладания илистых осадков с повышенным содержанием органических веществ формируются комплексы организмов с детритным типом питания, которые потребляют органическое вещество поверхностного и подповерхностного слоя грунта; однако если в опресненой зоне доминируют мелкие виды детритофагов, то в мористом районе преобладают более крупные организмы этой группы, способные собирать детрит с поверхности осадка и частично в самом придонном слое воды на границе осадок/вода.
В зонах повышенной гидродинамики преобладали подвижные и неподвижные сестонофаги, фильтрующие придонный и наддонный слой воды и извлекающие из него органические частицы. Роль неподвижных сестонофагов (Mytilus edulis) многократно возрастает на выходе из эстуария, которые образуют здесь обширные банки. Они являются мощными биофильтрами эстуарных экосистем.
Продукция и потоки энергии в экосистеме эстуария
Энергетический поток в эстуариях слагается из двух каналов: основного детритного и второстепенного – пастбищного. В роли продуцентов выступают морские травы (галофиты соленых маршей, Zostera marina), макроводоросли (нитчатая зеленая водоросль Cladophora fracta, бурые водоросли – Fucus vesiculosus, Ascophyllum nodosum), микрофитобентосные и планктонные водоросли. Основную долю органического вещества в эстуарии р. Черной производят макрофиты: до 95-99 % в верхнем и нижнем горизонте литорали, а также верхней сублиторали и от 50-60 % в среднем горизонте.
Роль прибрежного фитопланктона, представленного морскими видами, мала (он беден) из-за повышенной мутности воды, а также сильного опреснения его поверхностного слоя (до 2 ‰). В живом виде макрофиты в основном потребляются крупными гастроподами (в основном литторинами и частично гидробиями), большинство же видов макробентоса – детритофаги.
Продукция бентосного сообщества опресненного района эстуария (168 ккал/м2 в год) значительно превосходила таковую в солоноватоморском (приближается к 0 ккал/м2 в год), что связано с высокой первичной продукцией макрофитов в опресненном районе (230-371 ккал/м2 в год в опресненном против 247-193 ккал/м2 в год в солоноватоморском) (рис. 8). Все продукционные показатели в д
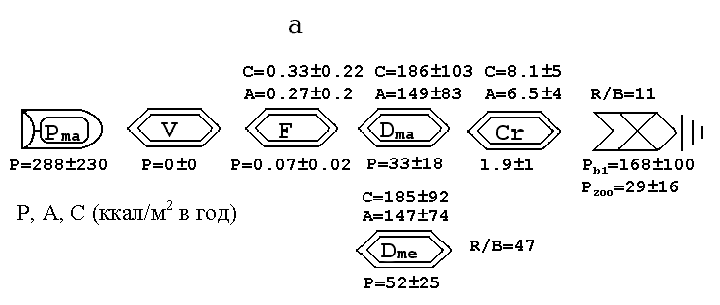
анном случае рассчитывались для всей прибрежной зоны (верхней, средней, н


ижней литорали и сублиторали).
Рис. 8. Потоки биоэнергии в разных районах эстуария. а – солоноватая зона, б – солоновато-морская зона, в – мидиевая банка. Pma – макрофиты, V – фитофаги,
F – фильтраторы, Dma – макродетритофаги , Cr – хищники, P – продукция, A – ассимиляция, C – потребление, Pzoo – общая продукция макрозообентоса, определяемая как сумма продукции всего зообентоса минус ассимиляция хищников, Pb – продукция бентосного сообщества равная разности между первичной продукцией растений (в нашем случае макрофитов) и общим дыханием гетеротрофных организмов (P-R).
Именно здесь в куту эстуария располагаются обширные соленые марши с высокими биомассами высших растений, которые после отмирания и разложения ассимилируются многочисленными детритоядными организмами. Продукция же макробеспозвоночных была значительно выше в солоноватоморском районе (75 ккал/м2 в год) и ниже в опресненном (29 ккал/м2 в год), а продукция мейобентосных животных, наоборот, - ниже в мористом (17 ккал/м2 в год) и выше в опресненном кутовом районе (52 ккал/м2 в год), что дополнительно свидетельствует о возрастании роли мелких организмов в опресненном районе эстуария.
Установлено, что в районе мидиевой банки, расположенной в зоне активной гидродинамики на выходе из эстуарного русла реки Черной, траты на дыхание гетеротрофных организмов значительно превосходили первичную продукцию растений (-1491 ккал/м2 в год), т.е. внутри системы неподвижных сестнофагов Mytilus edulis существует огромный дефицит энергии, который может быть компенсирован только импортом ее из опресненного района эстуария. Собственно в этом естественном биофильтре огромная потенциальная энергия, накопленная в опресненном эстуарном районе и сосредоточенная (рассредоточенная) в виде различных органических частиц (растворенных и нерастворенных) переходит во вторичную продукцию биофильтраторов Mytilus edulis и их прижизненных выделений (фекалиев и псевдофекалиев).
Основные и зависимые (донорские и реципиентные) подсистемы
Анализ вещественно-энергетического баланса показал, что в эстуарных экосистемах существуют два главных направления потоков биоэнергии. Первый направлен от высокопродуктивных горизонтов верхней литорали и соленого марша (первичная продукция растений превышает дыхание гетеротрофных организмов) к низкопродуктивным нижним горизонтам (песчаный пляж, сублитораль) (траты на дыхание консументов превышают первичную продукцию растений).
Второй поток энергии направлен от высокопродуктивных опресненных районов эстуария к энергетически бедной мидиевой банке, расположенной на выходе из эстуария в зоне активной гидродинамики.
Возникающие потоки энергии между разными районами эстуарной экосистемы за счет поступления избыточной энергии от энергетически богатых районов с высокой первичной продукцией к энергетически бедным обеспечивают ее целостность и стабильность во времени.
Типы питания и пищевые сети
В эстуарии преобладает детритная сеть питания, где основной группой животных являются собирающие детритофаги, а значительную долю пищевого комка большинства беспозвоночных животных составляет бесструктурное вещество – результат пищеварения различных детритных комплексов (табл. 4).
Таблица 4. Среднее процентное соотношение компонентов в пищевых
комках массовых видов (экспертная оценка - по содержимому кишечников)
Виды | Мин. части-цы | Оформлен-ные расти-тельные частицы | Zostera marina | Бесструк-турное вещество | Прочие |
| 1. Littorina littorea | 8.7 | 26.6 | 21.7 | 43 | 0 |
| 2. L. obtusata | 4.4 | 4.0 | 4.1 | 80.3 | 7.2 |
| 3. L. saxatilis | 12.8 | 17.6 | 7.6 | 54.8 | 7.2 |
| 4. Hydrobia ulvae | 33 | 0 | 0.5 | 63.3 | 3.2 |
| 5. Mya arenaria | 2.55 | 0 | 0 | 97.1 | 0.35 |
| 6. Macoma balthica | 63.1 | 0 | 0 | 38.7 | 0 |
| 7. Mytilus edulis | 3.75 | 0 | 0.18 | 93.5 | 2.27 |
| 9. Scoloplos armiger | 55 | 0 | 0 | 40 | 5.0 |
| 11. Tubificoides benedeni | 37 | 0 | 0 | 63 | 0 |
| 12. Chironomus salinarius | 9.7 | 0 | 0 | 90.3 | 0 |
Примечание. "Оформленные растительные частицы" – в основном фрагменты бурых (Chorda, Fucus), зеленых (Cladofora fracta), а также диатомовых водорослей. "Бесструктурное вещество" – результат пищеварения различных детритных комплексов.
Даже в зонах активной гидродинамики, где преобладают трофические группировки потребителей суспензий (сестонофаги), животные используют в качестве существенного источника питания взвесь детритных частиц (с ассоциированными микроорганизмами), которые образуются или непосредственно в эстуарии (в результате разложения многочисленных макрофитов и микрофитобентоса) или поступают с речными водами.
Таким образом, в эстуарной экосистеме одним из основных источников энергии является детрит и его производные, образованные в результате разложения многочисленных макрофитов и водорослей, а основной поток энергии к животным – детритофагам идет по кратчайшему пути: детрит макрофитов с ассоциированными микроорганизмами (бактерии, грибы, простейшие) – макрозообентос.
Эксперименты по искусственному увеличению плотности Macoma balthica L. (пищевые ресурсы эстуарной экосистемы)
Способность данной экосистемы поддерживать наличную биомассу организмов изучалась в серии экспериментов по искусственному увеличению плотности Macoma balthica (в 2-11 раз) – одного из доминирующих видов эстуария. В результате проведенного эксперимента было показано, что наличные ресурсы вполне достаточны для поддержания значительно более высокой плотности этих организмов (в 5-10 раз выше существующей), что соответствует двукратному увеличению суммарной биомассы остального макробентоса. При этом вы-сокая плотность не оказывает сколько-нибудь существенного влияния на основ-ные структурно-функциональные показатели сообщества в целом (табл. 5).
Таблица 5. Влияние 11 - кратного увеличения плотности Macoma balthica на структуру макробентоса за время эксперимента (май - август).
| Общие показатели | Начало эксперимента, май | Конец эксперимента, конец августа | |||||
| Контроль 1 (6 повтор,) | Опыт (10 повтор) | Контроль 2 (6 повтор) | |||||
| Средняя плотность | Средняя биомасса | Средняя плотность | Средняя биомасса | Средняя плотность | Средняя биомасса | ||
| Число видов | 9 | 17 | 14 | ||||
| Суммарные значения Все виды Все виды без мии | 276±129 275±129 | 4424±3208 1525±819 | 267±111 266±111 | 4877±2000 1789±1300 | 335±242 333±244 | 4826±2421 1840±1252 | |
| Средневзвешенная индивидуальная масса: Все виды Все виды без мии | | 16 6 | | 18 7 | | 14 6 | |
| Видовое разнообразие (индекс Шеннона) | 0.99 | 1.17 | 1.73 | 1.59 | 1.60 | 1.46 | |
| Выравненность (по Шеннону) | 0.34 | | 0.50 | | 0.46 | | |
Примечание. Доверительные интервалы установлены при уровне значимости p=0.05 для малых выборок на основе t-распределения по Стьюденту. Средняя плотность и биомасса даны в экз. на 1/16 м2 и в мг на 1/16 м2, соответственно.
Из таблицы видно, что число видов, общая плотность, биомасса, видовое разнообразие и выравненность сообщества макробентоса в опыте и контроле в конце эксперимента были очень близкими, что подтверждает и оценка структурного сходства (по плотности или биомассе популяций или по качественному составу) между двумя сообществами. Коэффициенты сходства были предельно высокими (99 %).
Таким образом, существующие ограничения в численности и биомассе гидробионтов обусловлены не дефицитом пространственных или пищевых ресурсов (большинство видов – детритофаги), но комплексом факторов среды (прежде всего солености), негативно влияющих на самые ранние стадии развития беспозвоночных.
Структура бентосного сообщества при разных условиях осадко-накопления
Некоторыми авторами было показано (Warwick, 1986, ABC- метод классификации), что эстуарные сообщества можно классифицировать как "сильно нарушенные", даже при отсутствии влияния других факторов, кроме естественных.
В качестве индикации нарушений в структуре эстуарных бентосных сообществ и оценке отклика этой структуры на органическое обогащение осадков использовался АВС-индекс (Beukema, 1988; Craeymeersch, 1991; Meire, Dereu, 1990; Warwick, 1986). Этот индекс основан на том, что биомасса как более консервативный показатель медленнее реагирует на изменения среды по сравнению с численностью, являющейся более динамичным показателем реакции сообщества. Поэтому в стабильных сообществах преобладают крупные виды с медленной динамикой, а в нарушенных – мелкие и более динамичные.
В эстуарии р. Черной вдоль продольной оси по мере увеличения солености воды наблюдается увеличение ABC-индексов от кутовых частей эстуария с низкими показателями к мидиевой банке с более высокими (в морской зоне значения ABC-индексов немного снижаются по сравнению с мидиевой банкой в связи с преобладанием в сообществе макробентоса более мелких полихет). Соответственно можно выделить две зоны с низкими и высокими значениями ABC-индексов. Пространственно границы этих зон почти совпадают с границами опресненной и мористой областей (мидиевая банка вместе с морской зоной) эстуария (табл. 6).
Таблица 6. Показатели АВС- индексов в разных районах эстуария р. Черной
| Горизонты прибрежной зоны | Средние значения ABC-индексов | ||||
| Солоноватая зона | Верхние пороги | Солоновато- морская зона | Мидиевая банка | Морская зона | |
| Сублитораль | -7.1 | -11.3 | +1.1 | +4 | +1.8 |
| Нижняя лит. | +1.64 | +3.4 | +4.45 | +10.4/+5 | +4.1 |
| Средняя лит. | +1.52 | +2.5 | +4.3 | +6.7/-4.2 | +3.1 |
| Верхняя лит. | -2.73 | -1.5 | -7.8 | -4.5/-4.8 | -2.7 |
Примечание. Середина мидиевой банки/Вся мидиевая банка.
Низкие значения ABC-индексов в опресненной области эстуария указывают, что опресненный район находится в зоне повышенного влияния не только солености воды, но и углеродной нагрузки. Повышенное содержание органического вещества в осадках опресненного района способствует формированию здесь сообщества мелких детритофагов - преимущественно r-стратегов. На выходе из эстуария в зоне активной гидродинамики формируется обширная мидевая банка с доминированием крупных сестонофагов Mytilus edulis. Также сле-дует отметить, что верхние горизонты литорали, представленные в основном мягкими грунтами, характеризуются более низкими и вариабельными показа-телями ABC – индексов по сравнению с нижними горизонтами.
ГЛАВА 4. Сравнительный анализ эстуарных экосистем, их разнообразие и значение
Классификация и типы эстуарных экосистем
Эстуарии имеют разный размер, протяженность, площадь и глубину акватории, разную степень изолированности от моря, а также различный водообмен с пресным стоком, разный градиент солености воды и, наконец, разное время существования. В зависимости от занимаемой площади эстуарии можно классифицировать – от микроэстуариев (приливные ручьи, стоки, лужи, ванны и т.д.) до мезо- (небольшие бухты, заливчики, зарегулированные небольшие водоемы, эстуарии небольших речек, соленые марши с расположенными на их территории водоемами), макро- (настоящие эстуарии средних рек, небольшие заливы, фиорды и т.д.) и, наконец, мегаэстуариев (большие заливы, дельты больших рек, сильно опресненные моря).
Разнообразие и пространственная структура эстуарных экосистем
Основные тенденции в структурных изменениях в продольном направлении – от устья реки в сторону моря и в вертикальном по мере повышения мареграфического уровня рассмотренных эстуариев совпадают: увеличиваются по мере увеличения солености воды и уменьшаются от сублиторали и нижних горизонтов литорали к верхним горизонтам. Однако степень выраженности обнаруженных изменений, количество структурных вариантов и переходов между ними значительно отличаются. Чем больше зон с различным режимом солености, чем значительнее и протяженнее градиент этого фактора, тем больше зон с различной видовой структурой и, соответственно, выше общее видовое разнообразие сообщества. Чем удаленнее от моря расположен эстуарий, тем большую роль в нем начинают играть пресноводные и солоноватоводные виды, чем ближе к основному бассейну, тем больше морских форм.
Заключение
В заключение следует подчеркнуть, что эстуарий – это уникальная экосистема со специфическими элементами, свойствами и взаимодействиями. Предложенная нами прототипическая модель эстуария характеризуется определенным набором экологических (сущностных) признаков. Во-первых, входящие в эстуарную экосистему сообщества организмов находятся в режиме перманентного изменения, контролируемого факторами среды. Во-вторых, пространственно-временная структура эстуарных сообществ в основном определяется солевым режимом водоема и в меньшей степени другими факторами (характером грунта, содержанием в них органического вещества и т.д.). В-третьих, сообщества эстуариев характеризуются низким межвидовым и межстанционным сходством, отсутствием сильных и стабильных во времени связей между большинством видов беспозвоночных (образуются так называемые квазиструктуры). При этом наивысшая интеграция видов в таких образованиях приходится на периоды повышения средней солености воды в эстуарии, а наивысшая дифференциация – на периоды распреснения. В-четвертых, распределение разных таксономических и размерных групп организмов по эстуарию имеет комплементарный характер, которое определяется их разным отношением к факторам среды (солености, содержанию органических веществ в осадках эстуария и т.д.). В-пятых, формирование структуры сообщества макробентоса во многом определяется пополнением за счет новообразующейся молоди. В эстуарии преимущественное развитие получают мелкие оппортунистические виды с растянутым нерестом, быстрым ростом и созреванием, большей защищенностью личинок, а ограничения в их численности и биомассе обусловлены комплексом факторов среды, негативно влияющих на самые ранние стадии развития беспозвоночных. В-шестых, эстуарии в основном относятся к гетеротрофным системам. Преобладает детритная сеть питания, роль пастбищного пути второстепенна (преимущественно в микробентосе). И, наконец, в-седьмых, основной поток биоэнергии в эстуарных экосистемах направлен от опресненных районов эстуария с высокой первичной продукцией макрофитов к энергетически зависимым плотным поселениям сестонофагов (мидиевой банке), расположенным на выходе из эстуарного русла.
Все эти перечисленные параметры являются обязательными при определении экосистемы как эстуария. Именно эти параметры являются опорными в построении прототипической модели эстуарной экосистемы. При наложении предложенной модели на другие эстуарные системы можно будет говорить о вариативных моделях эстуариев, что открывает перспективы дальнейших исследований этих уникальных и интересных объектов.