
М. В. Ломоносова биологический факультет н а правах рукописи Столяров Андрей Павлович Структурно-функциональная организация эстуарных экосистем Белого моря 03. 02. 10 гидробиология Автореферат
| Вид материала | Автореферат |
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- М. В. Ломоносова биологический факультет на правах рукописи никольская кира Алексеевна, 1297.21kb.
- Структурно-функциональная организация палеоамигдалы: фундаментальные закономерности, 2391.63kb.
- Юров иван юрьевич структурно-функциональная организация хромосом при нервно-психических, 850.19kb.
- Лекция № клетка элементарная биологическая система. Структурно функциональная организация, 341.33kb.
- Автореферат диссертации на соискание ученой степени, 1241.4kb.
- На правах рукописи, 470.03kb.
- На правах рукописи, 1724.93kb.
- На правах рукописи, 378.58kb.
- На правах рукописи, 594.43kb.
Глава 2. Объект, материалы и методы исследований
2.1. Объект исследования
В качестве объекта исследования были выбраны эстуарные экосистемы Белого моря с характерными сообществами трех основных размерно экологиче-ских групп: микро-, мейо- и макроорганизмов. Наибольшее внимание было сосредоточено на макробентосных сообществах, являющихся важнейшим компонентом мелководных прибрежных эстуарных экосистем.
2.2. Исходные данные и районы исследования
Исследования бентосных сообществ (микро-, мейо- и макробентос) проводились в основном в районе Чернореченской губы, расположенной в Кандалакшском заливе Белого моря. Чернореченская губа включает в себя значительное разнообразие эстуарных систем – от микроэстуариев мелких ручьев, небольших литоральных ванн до типичных макроэстуариев с значительным градиентом солености (0 – 24 ‰) и мезоэстуариев с менее сильными и резкими изменениями солености (5-8 – 22 ‰) и имеющих большую связь с основным бассейном Белого моря. Она включает в себя все основные структурно-функ-циональные единицы беломорских эстуарных экосистем – соленый марш с пре-обладающими макрофитами, мидиевую банку с доминированием фильтраторов Mytilus edulis, участки повышенных концентраций органических веществ – няши или накопители органики, зоны с различными солевыми режимами, сублиторальные и литоральные местообитания. Чернореченская эстуарная экосистема весьма детально изучена (25 лет исследований, начиная с 1985 г.) как в пространстве (эстуарий реки Черной, губа Грязная, Лапшагина губа, литоральные ручьи, крики и т.д.), так и во времени (в течение летних сезонов, начиная с мая и заканчивая сентябрем, а также – ряда лет). Исследования также проводились в губе Ермолинская, Чупа, Ругозерская.
Основу исходного материала составили оригинальные данные, полученные в ходе многолетних исследований (1985 – 2010 гг.). Общий объем использованного материала составил более 1000 проб по макро-, мейо- и микробентосу (около 800 проб макробентоса, 250 проб мейобентоса и 100 проб микробентоса).
Данные по макробентосу были собраны и обработаны в основном самостоятельно (1985-2010 гг.), по мейо- и микробентосу совместно с И.В. Бурков-ским, А.А. Мазеем, А.А. Удаловым (1994-2000, 2004 гг.), по микрофитобентосным водорослям использовался материал, предоставленный М.А. Сабуровой (2000 г.), по бактериям – В.В. Ильинским (2000 г.).
2.3. Методы сбора и анализа данных
Сбор материала и обработку данных проводили по стандартным гидробиологическим методикам. Для видовой идентификации использовали различные виды световой микроскопии.
В качестве интегральной характеристики сообщества были вычислены средние плотности и биомассы видовых популяций микро-, мейо- или мак-робентоса, число видов, индексы видового разнообразия Шеннона (Shannon, Weaver, 1949) и выравненности распределения обилий или биомасс видовых популяций Пиелоу (Pielou, 1966).
Для классификации видов или локальных сообществ, формирующихся на разных станциях, проводили кластерный анализ методом среднего присоединения на основе матриц сходства Съеренсена (Sorensen, 1948), Пианки (Pianka, 1974), Чекановского-Съеренсена (Песенко, 1982).
Для анализа размерной структуры сообщества и степени ее нарушенности использовался ABC-метод (abundance-biomass comparison method) (Warwick et al., 1987) и рассчитывался ABC-индекс (Meire, Dereu, 1990).
Для характеристики структурных изменений бентосных сообществ в динамических условиях эстуария использовали специальные показатели (общая гетерогенность, градиентность, дискретность), характеризующие структуру упорядоченных вдоль градиента солености или времени матриц сходства (Pielou, 1984).
Для расчета смертности пелагических и бентосных личинок использовали два показателя: dx - процент погибших планктонных личинок в период оседания на грунт или бентосных в период развития от личинки до сеголетка (за сезон) и интенсивность смертности (в сутки):
I = (ln N1 - ln Na) / T ,
где N1 – средняя плотность планктонных (в 1 м3) или бентосных (под 1 м2) личинок за сезон, Na – средняя плотность бентосных личинок (под 1 м2) или сеголетков (под 1 м2) за сезон, соответственно. T – время развития планктонной личинки от момента появления до оседания на грунт и время развития бентосной личинки до сеголетка (Бигон и др., 1989).
Все расчеты проводились при помощи пакетов программ MS Excel, 2002, SYSTAT 7, SYSTAT 10, ECOS 1.3 (Азовский, 1995).
Траты популяций на обмен рассчитывали по формуле: R = awbN, где R – траты на обмен, W – средняя масса особи поселений, оцененная как частное от деления биомассы (B) на плотность поселений (N), a – коэффициент интенсив-ности метаболизма, b – коэффициент регрессии. Коэффициенты a и b оценивали как средние из литературных источников (Алимов, 1989; Винберг, 1950, 1966; Голиков и др., 1985, 1988; 1990; Huebner, 1973).
Калорийность растений рассчитывали, исходя из средних значений калорийности сырого (или сухого) вещества. У Chlorophyta – 0.5, у Phaeophyta – 0.4 ккал на 1 г сырой массы, в среднем 0.5 (4.6) ккал (Алимов, 1989; Голиков и др., 1988; Катанская, 1981).
Калорийность макрозообентоса рассчитывали, исходя из средних значе-ний калорийности 1 г их сырой массы (Голиков и др., 1985, 1988, 1990).
Продукцию видовых популяций рассчитывали по удельной продукции (P/B) (Алимов, 1989; Голиков и др., 1985, 1988; Заика, 1972; Заика, 1983).
Продукция массовых видов ( Macoma balthica, Mya arenaria, Mytilus edulis и Hydrobia ulvae), популяции которых были представлены репрезентативной выборкой и для которых была получена информация об их размерной струк-туре, рассчитывалась по формуле:
P=N0W0 + ∑ Nср. (Wt+1 - Wt) ккал/м2 в год, где N0W0 – биомасса молоди и сеголеток, Nср. – средняя плотность поселений особей соседних генераций,
(Wt+1 - Wt) – ожидаемый прирост массы особей каждой генерации (Алимов, 1989; Винберг, 1968; Голиков, 1976, Голиков, Скарлато, 1977).
Для макрофитов годовую удельную продукцию принимали равной 1.2 (Блинова, Возжинская, 1971; Катанская, 1981).
Результаты вычислений оценивали за год при средней температуре по-верхностной воды в местах взятия проб (около 6 C) и Q10 = 2.25 (Винберг, 1983; Голиков и др., 1985, 1988, 1990).
Ассимилированную популяциями макробентоса энергию (A) определяли как сумму энергетических трат на дыхание (R) и продукцию (P) за год
(A=P+R ккал/м2 в год).
Потребление (C) определяли из расчета, что средняя усвояемость состав-ляет 75% (C = 1.25A ккал/м2 в год).
Продукцию всего бентосного сообщества раcсчитывали по формуле (Алимов, 1989, 2000; Винберг, 1968; Заика, 1972):
Pb= Pр+Pph+ Pd + Pst+ Pcr - Aph - Ad - Ast - Acr= Pр- Rph- Rd - Rst- Rcr, где
P – продукция, A – ассимиляция, R – траты на дыхание, Pр – растения (в нашем случае макрофиты), V – фитофаги, F – фильтраторы, Cr – хищники.
Продукцию зообентоса определяли как сумму продукции всего нехищного и хищного зообентоса минус ассимиляция хищников.
Все энергетические показатели были представлены в килокалориях.
ГЛАВА 3. Структурная организация эстуарных сообществ
3.1. Видовая и пространственная организация биоты
3.1.1. Уровень макробентоса
Видовой состав, разнообразие и пространственная структура
сообщества
Изменчивый солевой режим, мозаичное распределение грунтов и органи-ческого вещества, сложные рельеф дна и динамика придонной воды – все вместе обусловливают большое разнообразие условий существования для гид-робионтов эстуария.
Анализ состава макробентоса в разных частях бассейна эстуариев указывает на относительно плавную смену видов вдоль градиента солености, нарушаемую лишь при резких изменениях этого или других факторов, особенно при переходе от котловин к порогам. Выявленные здесь надвидовые структуры (комплексы видов) нередко оказываются довольно случайными собраниями видов, необязательно повторяющимися от съемки к съемке, особенно при сильном расхождении последних во времени. В целом с увеличением солености наблюдается рост общего числа видов, видового разнообразия, численности и биомассы большинства видов макробентоса (рис. 1).

Рис. 1. Изменение числа видов (S), общей плотности (N), биомассы (B) и выравненности (EN) сублиторального сообщества макробентоса в зависимости от расстояния по мере увеличения солености воды в эстуарии реки Черной. Обозначения: С – солоноватая, См – солоновато-морская и морская зоны эстуария, М.б. – мидиевая банка.
B существующем разнообразии видов выделяются пресноводные виды, выдерживающие незначительное осолонение (в среднем до 3-5 ‰) и встречаю-щиеся в куту эстуария, солоноватоводные и морские эвригалинные, преиму-щественно населяющие центральные районы эстуария (и встречающиеся в диапазоне 3-5 – 14-16 ‰), и морские менее эвригалинные виды, преимущественно обитающие в мористом районе эстуария (при солености выше 16-18 ‰). При этом большинство эстуарных видов относятся к морским эвригалинным формам, отличающимся степенью толерантности к солености. Это эвригалинные (широкоэвригалинные), выдерживающие значительные колебания солености и распространенные преимущественно от 3-5 до 14-16 ‰, морские менее эвригалинные (или мезоэвригалинные), в основном встречающиеся от 8 - 10 до
16-18 ‰, и морские еще менее эвригалинные виды (или плейоэвригалинные), в основном населяющие воды с соленостью выше 16 - 18 ‰.
Условная граница между различными вариантами сообщества эстуария проходит при солености 5-8 (10) ‰ и 14-16 (18) ‰. В результате в эстуарных экосистемах выделяются 3 района, или зоны, различающиеся видовым составом и структурой формирующихся сообществ: солоноватая (0 – 5 (8) ‰), солоновато-морская (8 (10) – 15 (16) ‰) и морская (выше 16-18 ‰) (рис.1). Особое положение занимают массовые поселения фильтраторов (в основном мидии), приуроченные к районам с повышенной подвижностью водных масс (пространственные сужения и пороги) и промежуточной соленостью (15-20 ‰), характерные для большинства исследованных эстуариев. Фильтраторы выполняют важную роль в функционировании всей морской прибрежной экосистемы, принимая участие в осаждении и очищении воды от минеральной и органической взвеси, поступающей с речным стоком. Эта схема биогеоценотической организации эстуариев типична для всего Белого моря.
Вертикальный градиент структуры сообщества
Смена видового состава происходит не только на градиенте солености, но и в зависимости от нулевой изобаты. С увеличением глубины (от верхней литорали в сублитораль) наблюдается заметное изменение состава гидробионтов, среди которых уменьшается доля пресноводных и солоноватоводных видов и возрастает доля морских. Эта тенденция в равной мере касается беспозвоночных и макрофитов. Однако видовой состав последних в пределах эстуария зависит в большей мере от положения относительно нуля глубин, чем от солености воды.
С понижением мареграфического уровня меняется не только видовой состав сообщества, но и общие показатели его структуры: число видов, разнообразие, общая плотность и биомасса. Они закономерно возрастают от верхней литорали к нижней и сублиторали (рис.2).
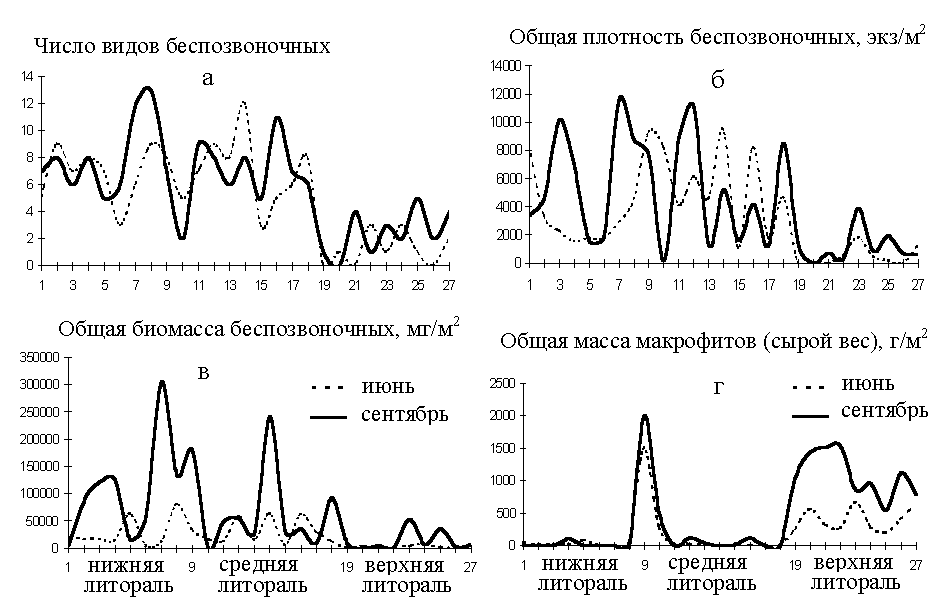
Рис. 2. Изменение числа видов (а), общей плотности (б) и биомассы беспозвоночных (в) и макрофитов (г) в эстуарии по мере увеличения солености воды. Расположение разрезов: 1 – 4 – в солоноватой части эстуария, 5 – 6 – в переходной в районе верхних порогов, 7- 9 – в солоновато-морской.
Таким образом, в эстуарных экосистемах наблюдаются два основных гра-диента факторов среды: продольный (вдоль эстуария - от опресненной части эс-туария в сторону моря) и вертикальный (от верхней литорали к нижней и сублиторали), формирующие соответствующие направленные изменения видовой структуры сообщества. Продольный градиент структуры сообщества обусловлен в первую очередь соленостью воды, скоростью осадконакопления, а поперечный (или вертикальный) определяется мареграфическим уровнем и связанными с ним факторами (временем осушения, перепадами температуры, опреснением, зарастанием литорали макрофитами, характером грунта и т.д.) (Азовский и др., 1998; Столяров и др., 2002; Столяров, Бурковский, 2005; 2008)
Сходство и различие локальных сообществ
Сообщества эстуариев характеризуются низким межвидовым и межстан-ционным сходством (среднее около 0.2), отсутствием сильных и стабильных во времени связей между большинством видов беспозвоночных, что свидетель-ствует о значительной неоднородности их видовой структуры, а также относи-тельно независимом распределении видов по эстуарию, более отвечающее континуальной модели. В целом выделенные группы видов и станций соотвествуют выделенным ранее комплексам видов (пресноводные, солоноватоводные и морские – разной степени эвригалинности) и районам эстуария (солоноватый, солоновато-морской и морской), значительно перекрываясь в своем распределении (рис. 3).
-
1
16
26
30
60
1──────┬┐ .
2──┬───┘│ .
3┬─┘ ├────────────┐ .
4┘ │ ├────────────┐
5───────┘ │ . │
6─────────┬──────────┘ . │
7─────────┘ . │
8────────┬┐ . │
9────────┘├────────┐ . ├───┐
10─────────┘ ├─┐ . │ │
11──────────────────┘ ├───────────┐│ │
12────────────────────┤ .││ │
13────────────────────┘ .││ │
14────────────────────────────────┼┘ ├──
15────────────────────────────────┤ │
16────────────────────────────────┘ │
17─────────────────────────────────────┤
18─────────────────────────────────────┤
19────────────────────────────┬┐ . │
20────────────────────────────┘│ . │
21─────────────────────────────┤ . │
22──────────────────────┬──────┼────┐ │
23───────────────┬──────┘ │ . │ │
24───────────────┘ │ . ├─┐│
25─────────────────────────────┘ . │ ││
26──────────────────────────────────┘ ││
27────────────────────────┐ . ││
28────────────────────────┼───────────┤│
29────────────────────────┘ . ├┘
30────────────────────┬───┐ . │
31────────────────────│ │ . │
32────────────────────┘ ├───┐ . │
33──────────────────┬─────│ │ . │
34──────────────────┘ │ ├────┐ │
35────────────────────────┘ │ . │ │
36──────────────────────────┬─┘ . │ │
37──────────────────────────┘ . │ │
38─────────────────────────────────┤ │
39────────────────────┐ . │ │
40─────────────────┬┐ │ . │ │
41─────────────────┘│ │ . │ │
42───────────────┬──┤ │ . │ │
43───────────┬───┘ │ │ . ├──┘
44───────────│ ├─┤ . │
45───────────┘ │ ├─┐ . │
46──────────────────│ │ │ . │
47─────────────┬────┘ │ │ . │
48─────────────┘ │ │ . │
49─────────────────┐ │ │ . │
50─────────────────┼──┘ │ . │
51───────────┬───┐ │ │ . │
52───────────┘ ├┐│ ├──────────┘
53───────────────┘├┘ │ .
54────────────────│ │ .
55───────────────┬┘ │ .
56───────────┬───┘ │ .
57───────────┘ │ .
58─────────────────┬─┐ │ .
59─────────────────┘ ├──┘ .
60───────────────────┘ .
├───────┼───────┼───────┼───────┼───────┤
1.0 0.8 0.6 0.4 0.2 0.0
Ст. 1-16 – Солоноватая зона
Ст. 17-25 – Солоновато-морская
Ст. 26-29 – Мидиевая банка
Ст. 30-60 – Морская зона
Рис. 3. Кластер-анализ по станциям (сублитораль). Расположение станций:
1-16 – солоноватая, 18-25 – солоновато-морская, 26-29 – мидиевая банка,
30- 60 – морская зоны эстуария. Точками показан уровень значимого сходства.
Из кластерограммы видно, что локальные сообщества мористого района эстуария более сходны по видовой структуре, чем сообщества опресненного. Причина этого – жесткие условия среды в солоноватоводном районе эстуария.
Сравнительный анализ структуры литорального и сублиторального сообществ
В ходе анализа выявлена тенденция уменьшения сходства видовой структуры сообщества от сублиторали и нижних горизонтов литорали к средним и верхним, что обусловлено низким видовым разнообразием и значительной вариабельностью структуры сообщества верхних горизонтов прибрежной зоны. Соответственно станции нижних горизонтов литорали и верхней сублиторали более сходны по видовой структуре, чем станции верхних горизонтов. Среднее сходство локальных сообществ верхней литорали было 0.27, средней - 0.34, нижней - 0.44, сублиторали - 0.50.
Наблюдается также снижение вдоль эстуарного градиента видовой структуры сообщества от сублиторали и нижней литорали к средней и верхней. При этом градиент хорошо выражен и достоверен в нижних и средних горизонтах прибрежной зоны (сублитораль, нижняя и средняя литораль) и отсутствует в верхних.
Размерная структура сообщества макрозообентоса.
Размерная структура макробентосных животных в разных районах эстуария существенно различается. Наблюдается увеличение размеров и индивидуального веса организмов макрозообентоса (и в июне, и в сентябре) от устья реки в сторону моря и от верхнего горизонта литорали к среднему и нижнему (табл. 1).
Таблица 1. Средний индивидуальный вес организмов макрозообентоса в разных районах и зонах эстуария р. Черной
| Горизонты прибрежной зоны | Усредненный вес организмов, мг | ||||
| Солонова-тая зона | Верхние пороги | Солоновато- морская зона | Мидиевая банка | Морская зона | |
| Сублитораль | 14±10 | 2.2±2 | 28.2±22.2 | 504±504 | 60±60 |
| Нижняя лит. | 10.9±3.9 | 32±12 | 28±15 | 88.4±170 | 30±15 |
| Средняя лит. | 5.2±2.3 | 33±15 | 11.4±4.6 | 19±46 | 12±5 |
| Верхняя лит. | 6.4±4 | 10.2±8 | 10±5 | 4.7±4.5 | 10±5 |
Повышение среднего индивидуального веса животных в продольном направлении от устья реки в сторону моря обусловлено повышением общей солености воды, уменьшением варьирования ее показателей в этом направлении и большей заиленностью опресненного участка эстуария, что приводит к смене мелких пресноводных и солоноватоводных беспозвоночных с низким индивидуальным весом (в основном олигохеты и хирономиды) более крупными морскими эвригалинными животными (главным образом моллюски: макома, мия, литторины и мидия). Снижение средней индивидуальной массы беспозвоночных в вертикальном направлении - от сублиторали и нижней литорали в сторону средней и верхней объясняется ухудшением условий обитания (увеличение времени осушения, большим опреснением верхней литорали и заилением) большинства крупных организмов (макома, литторина, мия, мидия), их уменьшением в числе и сменой на более мелких олигохет, хирономид и гидробию.