
М. В. Ломоносова биологический факультет на правах рукописи никольская кира Алексеевна системно-информационные аспекты познавательной деятельности позвоночных диссертация
| Вид материала | Диссертация |
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- М. В. Ломоносова биологический факультет н а правах рукописи Столяров Андрей Павлович, 657.12kb.
- Самовольцева Кира Алексеевна Дылевич Елена Валерьевна закон, 168.26kb.
- На правах рукописи Гуськова ирина Алексеевна, 682.76kb.
- На правах рукописи, 772.97kb.
- На правах рукописи, 315.69kb.
- На правах рукописи, 372.38kb.
- Тема: «Системно-деятельностный подход - универсальный инструмент познавательной деятельности», 64.45kb.
- Самовольцева Кира Алексеевна Агеева Елена Валерьевна закон, 118.95kb.
- На правах рукописи, 544.87kb.
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
имени М.В.ЛОМОНОСОВА
Биологический факультет
на правах рукописи
НИКОЛЬСКАЯ Кира Алексеевна
СИСТЕМНО-ИНФОРМАЦИОННЫЕ АСПЕКТЫ
ПОЗНАВАТЕЛЬНОЙ ДЕЯТЕЛЬНОСТИ ПОЗВОНОЧНЫХ
Диссертация
на соискание ученой степени доктора биологических наук,
представленная в форме научного доклада
03.03.01 – физиология
Москва - 2010

Общая характеристика работы
Актуальность проблемы. Сущность и содержание когнитивной активности, ее роль в организации поведения, обучения, памяти, а также причины развития различных психосоматических нарушений волновали ученых во все времена. Разнообразные научные дисциплины на протяжении многих тысячелетий изучали познавательную деятельность и немало специальных философских школ пытались дать ей общее определение. Свои частные определения предлагала и логика, и теория познания, и философская школа сознания, и нейронауки и психология.
В конце Х1Х в. возникла экспериментальная психология как наука, которая была призвана сформировать представления об общих закономерностях поведения и особенностях психических процессов. О необходимости ее появления говорит тот факт, что в начале ХХ в. возникло два независимых экспериментальных течения, роль которых в изучении тонких психических процессов трудно переоценить. Э. Торндайк в Америке основал объективно-психологическое направление, посвященное изучению двигательных форм поведения животных – навыков, которое в дальнейшем, благодаря Дж. Уотсону (1913), получило название «бихевиоризм». А в России И.П. Павлов основал физиологическое направление, в задачу которого входило изучение рефлекторных принципов деятельности головного мозга на примере слюноотделительного условного рефлекса. Поворотный момент в истории развития отечественной науки о высшей нервной деятельности (ВНД) произошел в 40-50г.г. ХХ столетия после того, когда было показано (П.К. Анохин, Э.А. Асратян, И.С. Бериташвили, Л.Г. Воронин, Н.П. Протопопов, П.С. Купалов и многие другие), что элементарные формы условных рефлексов не дают представление о всем многообразии и сложности интегративной деятельности мозга. Необходима была разработка более сложных поведенческих моделей.
Когнитивное направление, возникшее в 1956-70е гг. как протест против бихевиоризма (Bruner, 1964; Miller, 1964, 1979, 1990; Neisser, 1967; Piaget, 1970; Newell, 1973 и др.), провозгласило своей главной методологической доктриной изучение психических процессов, обуславливающих поведение (Хомский, 1991) и дало мощный толчок для изучения познавательной деятельности (ПД). Все эти способности изучались и вне когнитивизма, но именно последний внес в анализ новый ракурс рассмотрения: понимание того, как происходит обработка информации в мозге. Однако сохранение в современных когнитивных исследованиях бихевиористического подхода к оценке ПД не позволило реализовать эти идеи. Требовались новые концепции и методологические подходы. Одним из них стал информационный подход, который должен был исследовать механизмы переработки информации и ответить на вопрос о ее роли в формировании целенаправленного поведения. По мнению Ф. Розенблата (1965) ответ может быть дан только тогда, когда будет показано, как именно информация используется в целях управления действиями организма.
При знакомстве с современной литературой, посвященной различным аспектам ПД, она поражает противоречивостью мнений по всем вопросам, касающихся сущности, природы, процессов и даже ее структуры. В одних моделях провозглашается, что ПД - это система, манипулирующая представлениями (Newell, 1973; Pylyshyn, 1991; F. Keijzer, 2003;), поэтому не только репрезентации, но и процедуры их использования можно изучать, не обращаясь к уровню физической реализации (McShane, 1991). В других версиях, напротив, внимание сосредоточено на построении таких моделей интеллекта, которые принимают за основу нейронную организацию мозга (Smolensky, 1988; Shaw et al, 2006; Jung, Haier, 2007). Нет ясности и в отношении природы ментальных процессов: одни видят ее в физиолого-биохимических или морфологических перестройках мозга (Nauta, Karten, 1970; Plomin, DeFries,1998), в то время как другие настойчиво разрабатывают биофизическую основу интеллекта (Århem, Liljenstrom, 1997; Хазен, 2000). Разноголосица мнений о сущности познавательной деятельности сказалась и на вопросах, связанных с ее эволюцией. Несмотря на серьезную критику представлений о линейной зависимости функции от ее структуры (Barcroft, 1934), многие исследователи и сегодня полагают, что эволюция аналитико-синтетической деятельности определяется структурной организацией мозга (Воронин, 1977; Карамян, Малюкова, 1987; Jerison, 1978; Campbell, Hodos, 1991). При этом, существуют и прямо противоположные взгляды, отстаивающие концепцию, согласно которой интеллект - общее свойство, присущее всем видам животным, как и принципы мышления за исключением способности к языку у людей (Гельмгольц, 1856; Сеченов, 1879; Крушинский, 1979; Зорина, 1993; Macphail, 1985, 2002).
К сожалению, представления о принципах и закономерностях ПД животных остаются противоречивыми. в значительной мере причиной этого является отсутствие четких формализованных психофизиологических понятий, таких как «поведение», «мотивация», «обучение» «сложность» среды и задачи. Встречающиеся многочисленные модификации определений отражают либо различные подходы к пониманию этих психофизиологических дефиниций, либо их различные аспекты. Кроме того, отсутствие четких представлений о способах образования и уровнях сложности существующих форм условнорефлекторного поведения, их классификации создают серьезные трудности при интерпретации данных. К сожалению, на практике разработка методов познания осуществляется крайне вяло по сравнению с развитием самой науки. Еще в 1956г. Дж. Бернал с горечью констатировал, что ученые сначала находят что-то, а затем уже, как правило, безрезультатно размышляют о способах, которыми это было открыло. Понятно, что изучение технологии обучения само по себе не обеспечивает всеобъемлющего познания основ ВНД, но научная разработка их может помочь дисциплинировать ход исследования и определить возможную область изучения, а значит, и возможную область теоретических обобщений.
В русле двух направлений использовались два различных методических подхода для изучения аналитико-синтетической деятельности. В отечественной школе для получения сложных форм поведения обучение животных традиционно проводилось по методике наращивания звеньев цепи при участии экспериментатора (Воронин, 1952; Рокотова, 1954). Это привело к отождествлению познавательного процесса с феноменом цепных условных рефлексов. В рамках бихевиористического направления познавательные возможности животных исходно изучали с помощью методики свободного выбора (Торндайк, 1898), отличительная особенность которой состояла в том, что животное должно было самостоятельно решить проблемную ситуацию. Однако, до сих пор сравнительный анализ методических особенностей обучения и их роли в определении приспособительных возможностей сформированного поведения так и не проведен.
Причину разногласий в механизмах обучения ряд исследователей (Бериташвили, 1966; 1968; Купалов, 1978; Асратян, 1982; Воронин, 1989) видели не только в особенностях методических подходов, но и в специфике индивидуальных проявлений ВНД животных (Теплов, 1961; Крушинский, 1993). Несмотря на значительный вклад отечественной физиологической науки в изучение типологических закономерностей основных свойств ВНД, выполненных под руководством И.П. Павлова, в современных работах исследователи предпочитают обойти проблему разнообразия за счет привлечения различных статистических приемов, нивелируя, тем самым, индивидуальный аспект. Поиски маркеров поведения, с помощью которых можно было бы быстро оценить изучаемый признак, привели к использованию крайне упрощенных поведенческих моделей для оценки состояния ПД (открытое поле, крестообразный, Т- или У-образный лабиринты и др.), хотя еще в середине прошлого столетия было убедительно доказана несостоятельность таких моделей для выявления основных закономерностей ВНД животных (Воронин, 1952). Актуальность предпринятого исследования обусловлена тем, что без строго анализа, без понимания того, какие проявления ВНД изучаются при применении конкретного способа обучения и поведенческой модели, трудно разобраться в сложном лабиринте представлений о ПД, поведении и их эволюции.
Попытки различных научных школ представить картину ПД на основании частных, хотя и верных наблюдений привели к одностороннему освещению основных закономерностей интегративной деятельности. По оценкам ряда исследователей (Черниговский, 1962; Анохин, 1978; Батуев, 1986) современная нейробиология в значительной степени отошла от принципа системности в исследованиях ВНД по сравнению с представлениями, развивавшимися И.М. Сеченовым, И.П. Павловым, А.А. Ухтомским, А.Н. Северцовым, А.А. Заварзиным. К сожалению, это связано с печальным фактом пренебрежения к синтетическому мышлению и недооценкой общего над частным. Отсутствуют обоснованные информационные методы оценки аналитико-синтетических процессов при осуществлении ПД позвоночных. Поэтому чрезвычайно актуальна разработка нового концептуального подхода, основанного на принципах целостности и системности, адекватных, сопоставимых с человеческой деятельностью, познавательных моделей для понимания принципов и закономерностей ПД.
Цель исследования – экспериментально обосновать возможность использования системно-информационного подхода для изучения закономерностей познавательной деятельности и других проявлений ВНД у различных представителей позвоночных в норме, а также подвергнутых различным функциональным и повреждающим воздействиям.
Задачи исследования:
- Разработать системно-информационного метод на основе привлечения ряда положений семиотики и теории информации, позволяющий изучать принципы ПД и закономерности аналитико-синтетических процессов, обеспечивающих решение проблемной ситуации и организацию целенаправленного поведения у позвоночных.
- Сформулировать пищедобывательную задачу как аналог когнитивной деятельности человека и разработать конструкцию экспериментальной среды, позволяющей регулировать объем входной информации за счет изменения топологии пространства.
- Создать символьную систему регистрации действий животного, позволяющую осуществлять компьютерную запись поведения животного в реальном режиме времени, соотнести символы с определенным видом информации, а также разработать систему анализа информационных показателей, позволяющих сопоставить их с традиционными условнорефлекторными показателями обучения.
- Исследовать особенности организации многозвенного пищедобывательного поведения в зависимости от способа обучения: методики жесткого обусловливания в виде наращивания звеньев цепи и сведения в цепь и методики свободного выбора; оценить влияние метода на адаптивные свойства сформированного поведения.
- Выяснить необходимые и достаточные условия, обеспечивающие возможность осуществления ПД в условиях свободного выбора; изучить зависимость обучения от таких факторов, как: пространственная сложность среды, длительность пищевой депривации, условия подкрепления, длительность межопытного интервала и экспозиции информации (времени опыта).
- Использовать системно-информационный подход для выявления индивидуальных различий в проявлении когнитивных возможностей животного, психо-эмоционального паттерна, динамики обучения, функционирования памяти и особенностей возбудительно-тормозных процессов в норме и при функциональных (фармакологических и физических) и патологических (повреждениях структур мозга, нейровирусной инфекции, алкоголизации, 10-минутной клинической смерти) воздействиях.
- С помощью системно-информационного метода исследовать:
- возможные нейрохимические корреляты типологии ПД;
- роль гиппокампа и амигдалы в проявлении индивидуальных особенностей ПД животных;
- особенности ПД у различных представителей позвоночных животных, различающихся уровнем организации мозга.
Научная новизна.
Разработан новый подход на основе привлечения семиотических и информационных положений для системного изучения закономерностей ПД животных и аналитико-синтетических процессов, а также индивидуальных характеристик различных сторон проявления ВНД – когнитивных, условнорефлекторных, психо-эмоциональных и основных нервных процессов при решении проблемной пищедобывательной ситуации.
Показаны преимущества представления поведения в виде меняющейся последовательности символов, позволяющей отражать динамику аналитико-синтетических процессов. Установлена реальная возможность перехода от количественной фиксации результатов обучения к качественному описанию динамики процессов, обуславливающих этот результат. Выделены основные операции, лежащие в основе аналитико-синтетической деятельности, обеспечивающие распознавание различных видов информации, а также операционный локомоторный механизм их осуществления. Показан универсальный характер правил обработки информации при становлении целенаправленного поведения у всех изученных групп позвоночных, реализующихся по безусловному типу.
С помощью системно-информационного метода оказалось возможным в пределах одного эксперимента изучение индивидуальных особенностей ПД животных. Выделены три типа ПД и определены принципы, лежащие в основе различий протекания когнитивных процессов, психо-эмоциональных и условнорефлекторных проявлений. Установлена взаимосвязь между свойствами познавательного типа и характеристиками основных свойств нервных процессов.
Выявлена ведущая роль процедуры предъявления информации (последовательное или одновременное) в детерминации адаптивных возможностей формируемого условнорефлекторного поведения. Показана зависимость обнаружения эффектов различного рода воздействий на организм от процедуры предъявления информации.
Определены факторы, выполняющие системообразующую роль при реализации ПД животных в условиях свободного выбора, установлена оптимальные границы, в пределах которых возможно успешное решение пищедобывательной задачи, завершающейся образованием целенаправленного поведения.
Применение системно-информационного подхода позволило выявить роль гиппокампа и амигдалы в определении индивидуального рисунка ВНД животных. Обосновано представление, что основной психофизиологический эффект гиппокампэктомии проявляется в сужении поля внимания, затрудняющего процесс извлечения информации из долговременной памяти за счет резкого усиления инерционных свойств ЦНС, а амигдалэктомии – в ослаблении концентрации внимания за счет усиления возбудимости ЦНС, провоцирующей неустойчивость возбудительно-тормозных процессов.
Установлена корреляция нейрохимической активности мозга: медиаторной, белковой, ферментативной с индивидуальными особенностями ПД. Выявлена дифференцированная реакция корково-подкорковых систем мозга на индивидуальные проявления ПД: корковые нейроны (сенсомоторная кора) обнаруживают избирательную чувствительность, а подкорковые (хвостатое ядро) и лимбические образования (гиппокамп и амигдала) отвечают однонаправленными нейрохимическими перестройками.
Использование системно-информационного метода позволило установить, что основной эволюционный прогресс в области ВНД при повышении уровня организации мозга отражен в интенсификации информационного процессинга, обеспечивающего более «экономные» в когнитивном отношении затраты при решении задачи. Обосновано представление, что когнитивная функция как свойство ВНД можно рассматривать в качестве эволюционного фактора, определяющего условия среды обитания, в которых данные виды смогут преодолеть отрицательное давление отбора и успешно ее освоить.
практическое значение работы.
Предложенный информационный подход на основе символьного представления поведения животного позволяет изучать не только закономерности проявления ВНД, но и специфику аналитико-синтетических операций, обеспечивающих распознавание различных видов информации, необходимых для решения задачи и организации целенаправленного поведения.
Разработанный системно-информационный метод позволяет в пределах одного эксперимента получать исчерпывающую характеристику индивидуальных особенностей протекания когнитивных процессов, психо-эмоциональных и условнорефлекторных проявлений, основных нервных процессов, способствует формированию представлений о типологии ПД животных, закономерностях осуществления и структурно-биохимических механизмах, обуславливающих специфические черты их проявления. Высокая чувствительность и результативность системно-информационного метода может служить основой для осуществления дифференцированного подхода при оценке различных воздействий на организм.
Результаты исследования расширяют представления о необходимых и достаточных условиях, обеспечивающих успешность обучения в ситуации свободного выбора, результатом которого является образование целенаправленного поведения. Знание оптимальной области действия установленных факторов обучения обеспечивает условия для корректного проведения эксперимента.
Разработанная поведенческая модель для изучения различных проявлений ВНД в многоальтернативном лабиринте, а также основные принципы системно-информационного анализа состояния животных имеют применение в прикладных исследованиях медико-биологического профиля и использованы при моделировании нейровирусных инфекций (Институт полиомиелита, РАН, Москва), клинической смерти (Институт реаниматологии, РАН, Москва), структурной патологии мозга (Институт ВНД и нейрофизиологии, РАН, Москва; СПб университет, Санкт-Петербург), корсакоподобных синдромов (кафедра ВНД Биологического ф-та МГУ, Москва), воспалительных процессов (Институт Морфологии человека, РАН, Москва), приобретенного влечения к алкоголю (Институт Мозга РАН, Москва), развития претромбозного состояния при действии различных факторов (кафедра физиологии животных и человека Биологического ф-та МГУ, Москва), пищедобывательного поведения у муравьев (кафедра ВНД Биологического ф-та МГУ, Москва), инфаркта-миокарда при физической и холодовой нагрузках (Кемеровский государственный медицинский институт, Кемерово), влияния электростатических полей на организм (Центр медико-биологических и экологических исследований проблем МЭТПиПС, Юрмала, Латвия).
Высокие прогностические возможности системно-информационного подхода для выявления дефектов в сфере проявления различных сторон ВНД при фармакологических, физических и повреждающих воздействиях, а также результаты моделирования различных состояний позволяют рекомендовать его для использования в доклинических тестовых испытаниях.
Основные положения системно-информационного подхода для изучения ПД животных включены в учебное пособие, результаты исследования используются в лекционном курсе по основам ВНД животных и человека и в качестве задачи в рамках Большого практикума для освоения студентами методики свободного выбора и принципов системно-информационного анализа при изучении различных проявлений ВНД, проводимых на Биологическом факультете МГУ им . М.В. Ломоносова.
Имеется авторское свидетельство за № 1382256 на изобретение "Способ моделирования Корсаковского симптомокомплекса", зарегистрированного в Государственном реестре изобретений СССР от 15 ноября 1987 г.
Положения, выносимые на защиту:
- У всех исследованных позвоночных, независимо от уровня организации мозга и индивидуальных когнитивных возможностей, в качестве механизма познавательной деятельности выступает процесс, реализуемый в поведении в виде замкнутого маршрута «кольцо». С помощью этого кванта поведения, осуществляемого по безусловному типу, в процессе обучения выполняются операции: разбиение, зеркальный поворот, инверсия и комбинаторные перестановки, позволяющие формировать представления о пространстве на основе ассоциирования маршрутных отрезков, распознавание структуры задачи на основе дифференцирования и формирование правила поведения на основе интегрирования.
- Существующие способы получения сложных форм поведения: методика жесткого обусловливания (наращивание звеньев цепи и сведение в цепь) и методика свободного выбора определяют пластичность условнорефлекторного ответа. Методика жесткого обусловливания формирует поведение, адаптивное только в тех условиях, в которых оно было выработано. Методика свободного выбора позволяет перестраивать поведение при изменении условий среды и задачи.
- Возможность обучения животного в ситуации свободного выбора при наличии подкрепления определяется системообразующими факторами, обеспечивающими сопряженность осуществления ассоциативного и интегративного процессов. Уровень мотивационного состояния животного детерминирует интенсивность информационного процессинга, длительность экспозиции информации и межопытного интервала – условия перехода к аналитико-синтетическому процессу, величина многоальтернативности среды – возможность распознавания структуры задачи.
- Системно-информационный подход позволяет в пределах социума выделить три группы животных, различающихся уровнем когнитивных возможностей (интеллекта) – способности прогнозировать поведенческий ответ: прогнозировать результат на основе процесса ассоциирования, прогнозировать действие на основе процесса анализа и прогнозировать результат действия на основе процесса обобщения. Уровню интеллекта соответствует определенный тип ПД, обусловливающий принцип организации поведения: 1) сигнальный, организующий поведение по жесткому типу на основе образования элементарных SR связей; 2) процедурный, организующий поведение по типу цепочек из элементарных связей на основе селекции; 3) концептуальный, организующий поведение на основе компиляции цепочек в схему (план) за счет способности устанавливать каузальные связи – Si Sk.
- Познавательная деятельность имеет свое отражение на различных уровнях нейрохимической активности мозга: медиаторном, белковом, активности ферментов углеводного и белкового обменов в нейронах. Характер и направленность этих проявлений определяются структурной топографией нейрональной активности, индивидуальные особенности которых обусловлены физиологической, нейрохимической и морфо-функциональной гетерогенностью мозга.
- Гиппокамп и амигдала участвуют в проявлении индивидуальных различий возбудительно-тормозных отношений в соответствии с генетически заданными возбуждающими гиппокампальными и тормозными амигдалярными свойствами. В психофизиологическом аспекте функциональная роль этих структур заключается в организации «энергетического» обеспечения функции внимания: гиппокамп участвует в обеспечении функциональной лабильности доминантного очага – способности переключаться с одного локального центра на другой, а амигдала – в обеспечении его устойчивости за счет сдерживания возбуждения от чрезмерного и неуправляемого распространения из фокуса доминанты.
- В эволюционном отношении принципы и закономерности познавательной деятельности едины для всех наземных позвоночных животных, несмотря на различия в организации мозга, возможностей и способа реализации поведения. Основные различия касаются морфофункционального и биохимического обеспечения (воспринимающих, интегративных и исполнительных механизмов) информационного процессинга, которое прогрессирует в ряду наземных млекопитающих. Интеллектуальные возможности вторичноводных позвоночных, не выдержавших давление отбора в наземных условиях и вернувшихся в более простую в информационном отношении водную среду обитания, ограничены жестким типом обучения.
Апробация работы.
Материалы диссертационной работы доложены: на Совещании по физиологии и патофизиологии лимбико-ретикулярного комплекса (Москва, 1968), 22-м Всесоюзном совещании по проблемам высшей нервной деятельности (Рязань, 1969), конференции «Память и следовые процессы» (Пущино-на-Оке, 1970), 6-м научном совещание и симпозиуме по эволюционной физиологии (Ленинград, 1972), 23-м Всесоюзном совещании по проблемам высшей нервной деятельности (Горький, 1972), 24-м Всесоюзном совещании по проблемам высшей нервной деятельности (Москва, 1974), 8-м Всесоюзном симпозиуме по проблемам кибернетики (Тбилиси, 1976), 25-м Всесоюзном совещании по проблемам высшей нервной деятельности (Ленинград, 1977), 13-м съезде Всесоюзного физиологического общества им. И.П. Павлова (Алма-Ата, 1979), Совещании «Гагрские беседы» (Тбилиси, 1979), 26-м Всесоюзном совещании по проблемам высшей нервной деятельности (Ленинград, 1981), Всесоюзной конференции "Проблемы нейрокибернетики" (Ростов-на-Дону, 1983), на 27-м Всесоюзном совещании по проблемам высшей нервной деятельности (Ленинград, 1984); 9-м Всесоюзном совещании по изучению, охране и рациональном использовании морских млекопитающих (Архангельск, 1986), на XV съезде Всесоюзного физиологического общества им. И.П. Павлова, (Ленинград, 1987); на Всесоюзной конференции "Сравнительная физиология высшей нервной деятельности человека и животных" (Москва, 1988), 28-м Всесоюзном совещании по проблемам высшей нервной деятельности. (Ленинград, 1989), 3rd Intern. Sympos. on experimental and clinical neurobiology (Stara Lesna, 1991), Intern. Symposium «Sensory systems and behavior of aquatic mammals» (Mосква, 1991), 11-й Международной конференции по проблемам нейрокибернетики (Ростов-на-Дону, 1995), 3rd Intern. Fibrinogen Symposium. Germany, 1996), Second World Congress for Electricity and Magnetism in Biology and Medicine (Bologna, 1997), Intern. Symposium «Cognition, emotion and autonomic responses» (Amsterdam, 1999), Всероссийской научной конференции с международным участием, посвященной 150-лет. со дня рождения акад. И.П.Павлова, (Санкт-Петербург, 1999); Международной конференции, посвященной150-летию. И.П. Павлова: "Новое в концепциях о механизмах ассоциативного обучения и памяти", (Москва, 1999), на 30-м Всероссийском совещании по проблемам высшей нервной деятельности, посвященном 150-летию со дня рождения И.П. Павлова, (Санкт-Петербург, 2000), 3rd International conference on Methods and Techniques in Behavioral Research, (Nijmegen, 2000), Meeting of EBPS and IGBP “Schizophrenia” (Amsterdam, 2000), Meeting of the Society for the Quantitative Analyses of Behavior (SQAB) (Washington, 2000), 18-й съезд физиологического общества им.И.П. Павлова. (Казань, 2001), 16th Intern. Congress on Thrombosis and Harmostasis. (Paris, 2001), 12-м Международном совещании по эволюционной физиологии (Санкт-Петербург, 2001), 11 World Congress of Psychophysiology (Ontario, 2002), Юбилейной международной конференции по проблемам нейрокибернетики (Ростов-на-Дону, 2002), Euroconference “Neural Mechanisms of Learning and Memory”. (Paris, 2002), Международной конференции «Вероятностные идеи в науке и философии», (Новосибирск, 2003), Первой Российской конференции по когнитивным наукам Казань, 2004), 2-й Международной конференции по когнитивным наукам (Санкт-Петербург, 2006), Международном совещании «Гиппокамп и память» (Пущино-на-Оке, 2006, 2009), 4-ой Международной конференции «Биологические основы индивидуальной чувствительности к психотропным препаратам (Москва, 2006).
Публикации. По теме диссертации опубликовано 108 работ, из них 56 в ведущих рецензируемых научных журналах, рекомендованных ВАК РФ и 1 коллективная монография.
Основные материалы и методы исследования.
Экспериментальные животные. Работа выполнена на животных различных таксономических групп: рыбах (золотых рыбках -Carassius auratus, n = 20 и карпах -Cyprinus carpio, n = 17) при участии И. Сабитовой и А. Сабитова, рептилиях (черепахи - Clemmys caspica, n = 13) совместно с Ш.К. Сагимбаевой, А. Л. Шереметьевым; на насекомоядных (ежи - Erinaceus europaeus, n = 14) при участии А. Л. Шереметьева в лаборатории эволюционной физиологии (руков. чл.-корр. АН СССР, А. И. Карамян) Института эволюционной биохимии и физиологии, г. Ленинград, на грызунах (кролики породы шиншилла, n = 80; крысы: беспородные, n = 100 и линии Вистар, n = 200, мыши линии ВАLB/c, n = 350; совместно с Ш.К. Сагимбаевой, С.В. Ожерелковым, А.В. Савоненко, Т.Н. Мельниковой О.В. Ещенко, В.Н. Шпиньковой, В.Н. Костенковой, А.И. Осиповым, А.И. Расинским, Н.М. Хоничевой, М.В. Кондашевской; на хищниках (хори Mustela putorius, n = 36) совместно с Ш.К. Сагимбаевой; обезьянах (Macaca mulatta, n = 11) совместно с Ш.К. Сагимбаевой в приматологической лаборатории (руков. лаб. - дбн Л.А. Фирсов) Ин-та физиологии им. И.П. Павлова, в г. Колтуши; дельфинах (Tursiops truncatus) совместно с О.П. Петровым, В.Н. Петрушиным на Севастопольской базе.
Работа выполнена на половозрелых самцах. Длительность опыта составляла 10 мин у мышей, 13-15 мин у крыс и ежей, 20 мин у хорей, обезьян и кроликов; пищевая депривация - 22-24 ч. Каждое животное после опыта докармливалось в течение 45 мин. Закономерности ПД были изучены как у интактных позвоночных, так и у крыс Вистар, подвергнутых различным воздействиям:
(1) фармакологическим - пирацетам фирмы “Sigma” (Германия) в дозе 40 и 250 мг/кг (совместно с Кондашевской), опилонг – синтетический аналог дерморфина (Всесоюзный кардиологический научный центр РАМН) в дозе 50 и 150 мкг/кг (совместно с Ещенко, Савоненко, Мельниковой, Осиповым, Расинским) и высокомолекулярный гепарин фирмы «Serva» (Германия, 10 kDa, активность 180 МЕ/мг) в дозе 16 МЕ/250 (совместно с Кондашевской), которые вводили ежедневно внутримышечно или внутрибрюшинно в течение 5 дней до обучения;
(2) физическим (совместно с Ещенко, Шпиньковой, Костенковой) - обучение крыс Вистар на фоне слабой неоднородности магнитного поля, которую создавали с помощью 3 магнитов, расположенных под лабиринтом на расстоянии 5 см. Значения магнитной индукции, измеренные магнитометром ИЗМИРАН (Ин-т МИФИ РАН), колебались от 35 до 300 мкТл;
(3) повреждающим локальным: электролитическому билатеральному разрушению у беспородных и крыс Вистар дорсального гиппокампа током 2.3 мA в течение 35с, амигдалы – 3.7 мA, 35с, мамиллярных тел - 2.5 мА, 40с, которое производили стереотаксически под нембуталовым наркозом (40мг/кг) через нихромовые электроды (0.4 мм); координаты определяли по атласу Fifkova, Marshala (1962); контролем служили интактные и ложнооперированные крысы; которых брали в эксперимент после 2-х недельного восстановительного периода;
(4) повреждающим системным: алкоголизации (совместно с Сагимбаевой, Кешелавой) крыс, потреблявших в течение 8-ми месяцев исключительно 15% раствор этанола; состояние структур мозга оценивали с помощью световой (совместно с Э.Н. Поповой, Г.Н. Кривицкой) и электронной (совместно с С.Н. Кешелавой) микроскопии, выполненной в Институте Мозга РАМН и на кафедре гистологии МГУ, нейровирусной инфекции (совместно с Ожерелковым), вызываемой вакцинным штаммом вируса желтой лихорадки (17Д), вирусами клещевого энцефалита (КЭ) и Лангат и клинической смерти (КС) – 10-минутной полной остановки кровообращения (совместно с Костенковой, Ю.В. Заржецким) у крыс Вистар; моделирование КС и последующую реанимацию выполняли сотрудники НИИ общей реаниматологии РАМН. В эксперимент крыс брали после полного восстановления двигательной активности, через 10-12 дней.
Характеристика экспериментальных сред.
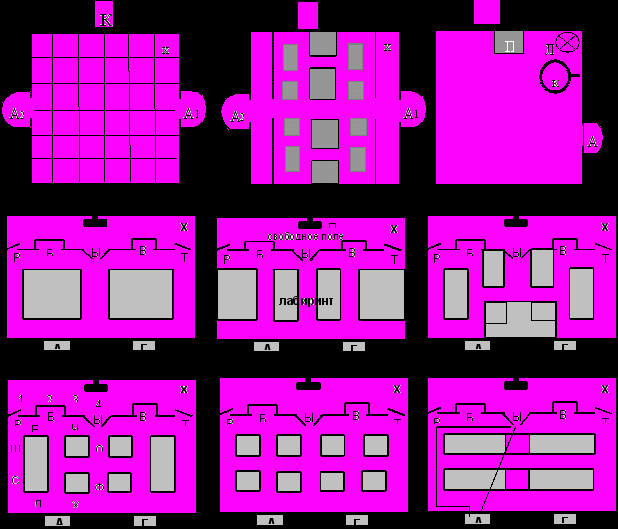
Рис. 1. Примеры экспериментальных сред.
К1-К3 – варианты открытого поля: Ы –вход, А – кормушка, П – полка, Л – лампа, К – кольцо. Л1-Л5 – примеры лабиринтных сред: П – полка, А и Г – подкрепляющие кормушки, Б и В – ложные, Р и Т – выходы; Нв – неопределенность выбора решения, рассчитанная по формуле Шеннона (1963); крестиком отмечено стартовое положение животного; в Л6 стрелкой показан вектор направления на цель, прямоугольники – непроходимые участки; заштрихованные зоны открывали после проведения процедуры первичного обучения.
Поведение животных исследовали в трех типах сред: «жилая комната», открытое поле и лабиринты различных модификаций. жилая комната представляла часть лабораторного интерьера размером 3 м х 1.5 м, состояла из столов, кресла, книжных полок и полуоткрытых ящиков, где животные могли свободно перемещаться в течение 3-4 ч. Среду использовали для классификации крыс по их двигательной активности в условиях социума, которую регистрировали в течение 15 мин до начала основного эксперимента (3 опыта в течении двух недель); открытое поле представляло камеру с одной либо с двумя подкрепляющими кормушками, расположенных друг напротив друга (рис. 1, К1). Среду усложняли либо за счет пространственной сложности (рис. 1, К2) – создавали лабиринтное поле, которое помещали в зону между кормушками, либо за счет введения дополнительных тонических (полка, кольцо) и фазических (свет, звук) раздражителей (рис. 1, К3). Лабиринты представляли камеру, разделенную перегородкой на две части: свободное поле и лабиринтную часть (рис.1, Л1-Л6), которые соединялись тремя дверями, открывающимися в одном направлении (один вход в лабиринт и два выхода в свободное поле). Пространство лабиринта было симметричным и в зависимости от числа разветвлений могло иметь несколько осей симметрии.
Поведенческие модели. Задачу на чередование места подкрепления – повторное подкрепление в данной кормушке можно получить только в том случае, если животное перед этим посетило другую (вторую), расположенную на противоположной стороне – исследовали в двух вариантах экспериментальных сред: К1 и К2 (рис. 1). Алгоритм навыка представлял собой многократное чередование посещения мест кормления: А1А2 в пределах фиксированного времени опыта.
Пищедобывательную задачу изучали в двух вариантах: в открытом поле К3 и лабиринтах (рис. 1). В первом случае животные должны были сформировать 5-звенный ситуационный условный рефлекс в циклической форме: при помещении в камеру животное самопроизвольно должно было подойти к полке (Sобстановка*Sполка Д1 = 1-е звено), нажать на нее (SполкаД2 = 2-е звено), в результате включался свет – 100Лк, на который животное должно было подойти к рычагу (Sсвет*Sкольцо Д3 = 3-е звено), дернуть его (SкольцоД4 = 4-е звено), подойти к кормушке и получить пищу (SкормушкаД5 = 5-е звено). После этого животное могло повторить всю последовательность действий.
В лабиринтных средах животным предлагали решить проблемную пищедобывательную задачу – чтобы многократно получать подкрепление в кормушках, необходимо после взятия пищи в пищевой среде (лабиринт) самопроизвольно выйти в непищевую среду (свободное поле) для того, чтобы при повторном заходе в лабиринт в тех же кормушках иметь возможность получить пищу. Алгоритм поведения мог включать от 4-х до 5-ти оперантных действий: при помещении в стартовую точку свободного поля (рис. 1) животное должно было самопроизвольно подойти к полке и нажать на нее для того, чтобы открылся вход в лабиринт (в случае 5-звенного навыка) или сразу подойти ко входной двери (в случае 4-звенного навыка). В лабиринте найти одну подкрепляющую кормушку или две из 4-х имеющихся, выйти самопроизвольно в свободное поле через один из двух выходов, после чего можно повторить весь цикл действий.
Процедура обучения. В работе использовали три методики обучения: наращивание звеньев цепи, сведение в цепь и свободный выбор. В первом случае (рис. 1; К3, Л3) экспериментатор управлял обучением, используя различные подсказки (шорох, подталкивание рукой), помогающие животному решить проблемную ситуацию. Во втором случае (Л3) обучение проводили на каждой половине лабиринта с последующим объединением их в единое целое. В третьем случае (Л3-Л6) животному предоставлялась возможность без предварительного знакомства со средой самому решить задачу без участия экспериментатора. Во всех случаях животное имело возможность корректировать ошибки. После завершения обучения проводили следующие тесты: (1) переделка навыка, суть которой заключалась в изменении значимости ложных и подкрепляющих кормушек при сохранности структуры задачи; (2) на «сытость» – выполнение навыка на сытом фоне (1 опыт); (3) на пластичность поведения – выполнение навыка при изменении конфигурации лабиринта (установка перегородки) в пределах пробы, мешающая животному выполнить предпочитаемый маршрут.
Анализ данных. Для получения количественных (сколько и/или как долго) и качественных данных (что и как делает) все экспериментальное пространство разбивалось на зоны и каждой присваивали свой знак. Запись поведения животного в опыте представляла собой текст в виде последовательности символов: например, 7s6os|5oh4ЫsОohs ИКА+ЛЕРswКИ ohhМЯГ+ГsЗУsoЩТ. Свободное поле промаркировано цифрами, лабиринтное пространство обозначено заглавными буквами русского алфавита, семь из которых обозначали оперантные объекты (входная дверь, 4 кормушки и два выхода из лабиринта в свободное поле). Прописными латинскими буквами обозначены безусловные реакции, которые по степени выраженности двигательной активности и психо-эмоциональному напряжению объединены в 9 классов: (1) ориентировочные (принюхивание, стойки), (2) ориентационные (остановки, колебания в развилках), (3) груминг (умывание, чесание); (4) двигательная заторможенность (сидение, лежание), (5) пассивное избегание (замирание, скуление); (6) смещенные реакции (лизание, грызение предметов); (7) стрессовые реакции (чихание, зевание, отряхивание и т.п.); (8) невротические проявления (тики, прыжки, навязчивое чесание), (9) активное избегание (карабканье вверх, выпрыгивание из камеры). Первые два класса мы отнесли к контекстному типу, а остальные 7 классов к неконтекстному, среди последних выделяли пассивные – 3-5-й классы и активные – 6-9-й классы проявления.
Поведение регистрировали с помощью оригинальной, созданной совместно с А.И. Осиповым, компьютерной программы, позволявшей анализировать различные условнорефлекторные показатели обучения: число проб за опыт (NПр) и опытов (NОП), двигательную активность – число пройденных отсеков (ДA), время осуществления одной пробы как Топыта/NПР, скорость перемещения - Vпр = ДA/ Топыта в сек; среднюю длину маршрута пробы - число отсеков, пройденных за одно выполнение (Lпр); долю конкретной безусловнорефлекторной реакций (PБРi) из всех сделанных; длину непрерывного пробега - отрезок маршрута, свободный от безусловных реакций (Lrun) как показатель прогностических возможностей, согласно Келеру (1932); число подкреплений (Nп) и его вероятность; ошибочные действия (Noш) двух типов: подходы к ложным кормушкам, а также повторные подходы к подкрепляемым кормушкам до выхода из лабиринта. Поскольку в процессе обучения животное имело возможность корректировать ошибочные действия, каждая проба могла состоять из разных по качеству действий-попыток. При оценке когнитивных затрат, затраченных животным на распознавание условий задачи и формирование плана поведения учитывались не только правильные действия, но ошибочные посещения значимых (семантических) зон до выхода животного из лабиринта. Когнитивные затраты подсчитывались как относительно проб – Кз = [Nпр x (Nп + ∑Noш)]/104), так и относительно времени, Кз = [Nоп х (Nп + ∑Noш)]/103), которые животным были затрачены на распознавание задачи и формирование поведения.
На основе исходных показателей вычисляли интегральные характеристики поведения, такие как мотивационная активность (МА), поведенческая эффективность (ПЭ) и организованность навыка (Он), оцениваемые относительно идеального автомата – максимально организованной в данной среде живой системы, которой присваивались предельные эмпирические значения животного. Подсчет осуществлялся по формулам: МА = (Nпр,i/Nпр,a + Рп,i/Рп,a + Рвхкк,i /Рвхкк,а)/3; ПЭ = (ДA,i/ДA,a + Tпр,i/Tпр,a + Puca,i/Puca,a)/3; Он = (Lrun,i/Lrun,a + Nош,a/Nош,i + Lпр,a/ Lпр i/)/3, – где Р – вероятность, П – подкрепления, ВхКК- выполнение 2-звенной последовательности, значения показателей для идеального автомата: Nпр,a = 45; Nп,a = 88; РВхкк,а = 0.93; ДA,а = 810; Tпр,а = 17.5; Puca,а = 0.10; Lrun,a = 8; Nош,a = 4.
Для оценки аналитико-синтетических процессов проводили анализ частоты встречаемости символьных последовательностей в опыте разной длины (n-грамм = 2, 3, ….n) отрезков. Комбинации, повторенные в первом опыте два и более раз считали зафиксированными в долговременной памяти (на основании эмпирического опыта) и рассматривали в качестве информационных единиц (ИЕ), образующих исходный алфавит. Каждой ИЕ присваивали свой знак и в соответствии с этими правилами отслеживали момент завершения набора новых ИЕ в процессе обучения, интенсивность набора и его объем. С помощью условной вероятности (вероятность перехода) выясняли силу сцепления каждой ИЕ с другими единицами, а также процесс укрупнения отрезков в маршруты, звенья и решения. Проверяли ИЕ на зеркальность (выполнение симметричной последовательности действий) и инверсию (воспроизведение ИЕ в обратном порядке) и подсчитывали коэффициент таких операций в опыте как отношение числа данной операции ко всем выполненным ИЕ в опыте. Каждое звено и их последовательность, входящие в структуру решения, анализировали в соответствии с тремя критериями: момент первого неслучайного появления n-граммы (критерий 1), момент перехода к неслучайной реализации в каждом опыте (критерий 2) и с первой пробы в опыте (критерий 3). Критерий 2 и 3 использовали для оценки оперативной и долговременной памяти.
Тактику поведения в отношении заданной структуры задачи, процесс становления целенаправленности навыка и его оптимизацию осуществляли с помощью алгоритмического анализа. Для этого каждый маршрут разделяли на фрагменты (блоки), устанавливали принадлежность к типу решения в соответствии с классификацией подсчитывали их долю.
Оценку психо-эмоционального состояния суммарно за весь период обучения и в соответствии с этапом распознавания структуры задачи осуществляли с помощью частотного анализа появления различных реакций в поведении с учетом их весовых коэффициентов, которые предварительно были вычислены на основе большого массива данных (у 100 крыс Вистар за 30 опытов) по формуле: КБРi = 1/Р_NБРi – (Рi – вероятность возникновения реакции).
При оценке функционирования памяти на этапе стабилизации навыка использовали три параметра: готовность как показатель скорости извлечения памятного следа (Г – величина, обратная числу выполнений, сделанных до появления первого минимизированного решения); мобилизационная активность как параметр реконструкции памятного следа (Мак – время до выполнения последовательности из трех минимизированных решений); устойчивость как параметр стереотипной реализации в поведении (У – отношение максимальной длины минимизированных выполнений к их сумме - Nmin, i/ΣNmin).
Оценку основных свойств ВНД осуществляли на начальном этапе обучения (1-2-й опыты) с помощью следующих параметров: длительность возбуждения (ДВ) (при заходе в лабиринт) как среднее время непрерывной локомоторной активности (ДА), в течение которого значения ДА в пределах одной минуты не снижались более чем на 30%; уровень возбуждения (ИВ) – среднее число отсеков лабиринта, пройденных за период непрерывной локомоторной активности; под силой возбуждения (СВ) понимали среднюю скорость перехода из пассивного состояния (минимальное значение ДА) в активное и вычисляли по формуле СВ = d(ДAmax-ДAmin)/dT(мин); длительность торможения (ДТ) как средняя продолжительность минимальной локомоторной активности, в течение которой значения ДА не поднимались в пределах одной минуты более чем на 30%; глубина торможения (ИТ) как среднее число пройденных отсеков лабиринта за период минимальной активности. В качестве силы торможения (СТ) рассматривали среднюю скорость падения ДА до минимальных значений – СТ = d(ДAmax-ДAmin)/dT(мин).
Проявление основных процессов ВНД в условнорефлекторной деятельности оценивали на этапе стабилизации сформированного навыка как среднюю трех опытов. Длительность возбуждения (ДУРВ) определяли как время опыта, в течение которого частота заходов в лабиринт была не менее двух выполнений задачи в мин; интенсивность возбуждения (ИУРВ) как суммарное количество проб, совершенных за период ДУРВ, сила возбуждения (СУРВ) как продолжительность ДУВР, отнесенной ко всему времени опыта; длительность торможения (ДУРТ) как время, в течение которого животные последовательно выполняли минимизированные (безошибочные) пробы, интенсивность торможения (ИУРТ) как количество безошибочных проб, совершенных за период ДУРТ, под силой торможения (СУРТ) - отношение ДУРТ к ДУРВ.
При оценке биохимического состояния мозга крыс Вистар изучали особенности: (1) нейромедиаторного обмена (выполненного Е.Г. Доведовой) в сенсомоторной коре и хвостатом ядре; содержание дофамина (ДА), норадреналина (НА), серотонина (5-ОТ) и 5-гидро-ксиндолуксусной кислоты (5-ОИУК) флуориметрически (MPF-4, "Hitachi", Япония) определяли по методу Когана с соавт. (1979), спектрофотометрически (спектрофотометр Gilford, Франция) определяли активность МАО А при 250 нм по методу Попова с соавт., (1971), МАО Б при 540 нм по методу Горкина с соавт., (1968) и АХЭ при 540 нм по методу Hestrin (1949), радиометрически (счетчик 1211-Racbeta, Швеция) оценивали активность ацетилхолинтрансферазы ХАТ по методу Fannum (1969); (2) ферментативной активности мозга (выполненного А.В. Сергутиной) на примере активности глутаматдегидрогеназы (ГДГ) и глюкозо-6-фосфатдегидрогеназы (Г-6ФДГ) по методу Rubinstein et all.(1962), аминопептидазы (АМП) по методу Нахласа в модификации Герштейн (1965) в сенсомоторной коре (слои III и V), в дорсальном гиппокампе, хвостатом ядре и n.accumbens. Продукт цитохимических реакций фотометрировали на микроскопе ЛЮМАМ-И 3 (Россия) (диаметр зонда 2,5 мкм): оценку активности ГДГ и Г-6ФДГ проводили при 589 нм, АМП при 550 нм. В каждом образовании ЦНС животных активность ферментов измеряли в 100 нейронах; (3) состояния белкового обмена (выполненного А.В. Савоненко, В.Н. Шпиньковой) у крыс Вистар, обучавшихся в течении 25 опытов; интерферометрически (микроскоп ИНТЕРФАКО) определяли содержание и концентрацию белка в ядрах и цитоплазме нейронов с одновременным измерением площади профильных полей тела клетки и ядра в нейронах сенсомоторной коры (слои III и V), дорсальном гиппокампе и амигдале (базолатеральное ядро – n. ABL). У каждого животного было изучено по 40-60 нейронов в каждой структуре мозга. Результаты обрабатывали по программе "Protein", созданной в НИИ Мозга РАМН.
Для статистической обработки полученных данных использовали регрессионный анализ, непараметрический критерий Манна-Уитни, t-критерий Стьюдента, для оценки различий распределения критерий λ Колмогорова-Смирнова, метод Монте-Карло для определения значения случайного события.
Результаты исследований
-
системно-информационного подход. Его роль в изучении познавательной деятельности позвоночных
Идея возможности изучения ментальных процессов была сформулирована еще во второй половине позапрошлого столетия (Сеченов, 1878). Ее сущность заключалась в привлечении лингвистического подхода для изучения психической деятельности человека и животных, в центре внимания которого находится язык – система знаков, играющая важную роль в репрезентации информации. В дальнейшем идея рассмотрения языка как ментального феномена, как особой когнитивной способности, благодаря которой обработка информации есть оперирование символами, высказывалась учеными неоднократно (Витгенштейн, 1958; Бернштейн, 1965, Напалков, Целкова, 1974, Новиков, 1968; и др.), но реального воплощения в экспериментальной психофизиологии так и не получила.
Для реализации этой идеи в нашей работе в качестве экспериментального пространства была использована лабиринтная среда различной топологии (рис. 1) как аналог проблемной ситуации (Арбиб, 1972). Во всех случаях эта среда конструктивно была организована таким образом, чтобы животное в процессе обучения должно было посетить две в информационном отношении различные области: (1) простую в виде открытого поля (названной нами «свободным полем»), целостное представление о которой животное могло составить в пределах одного пробега и (2) сложную (лабиринт) за счет многочисленных разветвлений, обеспечивающих высокую неопределенность относительно поиска адекватного решения. Наличие двух сред и предоставленная животному возможность самому управлять своим поведением (в случае свободы выбора) позволяли исключить из процедуры обучения традиционно принятый межстимульный интервал и изучать познавательный процесс в зависимости от особенностей его мотивационной активности. Внесение фактора симметрии при организации пространства лабиринта позволило, с одной стороны, создать определенную трудность как в отношении пространственной ориентации, так и в распознавании структуры задачи, а с другой стороны, обеспечило условия для изучения способности улавливать этот признак и использовать при реализации аналитико-синтетической деятельности. Привлечение ряда положений теории графов (Swamy и Thulasiraman, 1981) и теории информации (Шеннон, 1963) позволило количественно оценивать пространственную сложность экспериментальной среды и делало возможным изучение влияния этого фактора на характер протекания ПД. В качестве меры использована величина неопределенности выбора маршрутной реализации при осуществлении поведения, которая учитывала как количество альтернативных зон в лабиринте, так и коэффициент разветвления.
Под познавательной деятельностью мы понимали аналитико-синтетические процессы, обусловливающие распознавание информации для решения проблемной ситуации и обеспечивающие на ее основе организацию целенаправленного поведения. Для углубленного изучения ПД животных мы воспользовались положением семиотики о свойствах знаковой системы как носителе трех видов информации в случае возникновения коммуникативной ситуации (внешняя среда – животное): синтаксической, семантической и прагматической (Черри, 1964; Смирнова, 1990). Согласно этому экспериментальную среду мы рассматривали как носитель знаков, а животное как носитель языка. В рамках развиваемого нами системно-информационного подхода задача животного при столкновении с проблемной ситуацией состояла в способности распознать три вида информации: синтаксическую - о топологии среды, (отвечающей на вопрос - что такое?), (2) семантическую - о структуре или логике задачи (что делать?) и (3) прагматическую - о структуре поведения (как делать?). Для создания знаковой системы каждому отсеку лабиринта и оперантным элементам (полка, двери, кормушки) присваивали свой знак, в результате чего пространству среды ставилась в соответствие матрица буквенных символов - от 14 до 28 (синтаксическая информация), часть из которых выделялась в роли организующих узлов пространства - семантических элементов задачи. Таким образом, регистрация действий животного в совокупности с закодированными психо-эмоциональными проявлениями на всех этапах обучения представляла собой меняющуюся последовательность знаков (текст).
Принципиальная особенность разработанного нами информационного подхода, состояла в том, что каждый знак, соотнесенный с определенным видом информации, потенциально отражал не только где находится животное, но и что и как оно делает. Вследствие этого запись опыта представляла собой историю (процесс) ПД животного в зашифрованном для экспериментатора виде, в котором содержалась информация об аналитико-синтетических процессах, направленных на распознавание трех видов информации. Полный набор символов рассматривали в качестве исходной матрицы синтаксической информации. Символы, кодирующие оперантные объекты (кормушки, выход, вход, полка и т.п.), часть из которых входили в структуру задачи, была отнесена нами к семантической информации, а совокупность оперантных действий определяли как прагматическая информация. Последовательность из двух действий (двуграмму) рассматривали в качестве наименьшей информационной единицы. Текстовая запись поведения животных позволяла на основе ее фрагментации и последующей обработки осуществить реконструкцию познавательного процесса, выделить основные операции, связанные с распознаванием информации, получить сведения об ассоциативных, аналитических и синтетических процессах ВНД. При этом информационный подход не исключал возможность получения количественных и скоростных показателей обучения, таких как число проб, ошибок, подкреплений, скорость обучения и т.п.
Были сформулированы критические требования к поведенческой модели: (1) ответ животного должен представлять не одиночный рефлекторный акт (выбор налево или направо, побежка к кормушке и т.п.), а поведение, которое, в соответствии с представлениями Бериташвили (1932), должно состоять из последовательности действий, требующих их организации и отражать понимание экспериментальной ситуации т.е. обладать признаками целенаправленности относительно задачи в соответствии с исходной потребностью; (2) правило поведения не должно определяться фазическими (детерминирующими ответ) условными раздражителями, включаемые экспериментатором; (3) изменения в среде должны возникать только в ответ на действия животного (например, выходная дверь открывается только в том случае, если животное подходит к ней и закрывается, если отходит от нее); (4) реализация правила поведения должна удовлетворять условию многоальтернативности; число равновозможных вариантов реализации решения задачи должно быть таково, чтобы при формировании правила необходима была проверка множества рабочих гипотез, и животное могло интерпретировать его в зависимости от своих индивидуальных когнитивных особенностей и приобретенного знания; (5) в пределах пробы животному должна быть предоставлена возможность коррекции ошибочных действий.
Для получения свидетельств об аналитико-синтетических процессах, требования к структуре задачи были следующими: (1) семантика когнитивной задачи по сложности должна быть модельным вариантом интеллектуальной деятельности человека, требующей наивысшего уровня интеллекта - прогнозирования результата действия, (Анохин, 1962; Бернштейн, 1963) и соответствовать многозвенной логике по типу: если совершить n-действий, то цель будет достигнута всегда; (2) объем предъявляемой информации о задаче должен превышать симультанность восприятия, объем которой, согласно Миллеру (1964), составляет 7±2 единиц информации; (3) число семантических элементов должно быть избыточно относительно тех, которые входят в структуру задачи;
Учитывая значимость сложных форм условного рефлекса для изучения различных сторон проявления ВНД (Воронин, 2002), а также методологическое требование соблюдения принципа подобия при моделировании процессов мышления у животных (Славская, 1968), всем изучаемым позвоночным предлагалось решить когнитивную задачу, структура которой соответствовала 4-звенной циклической логической конструкции. Сложность ее определялась тем, что для распознавания структуры задачи необходимо было сформировать как минимум пять условнорефлекторных зависимостей разной степени сложности: (1) свободное поле (СП) непищевая среда; (2) лабиринтное поле (ЛП) пищевая среда; (3) всякий заход из свободного поля в лабиринт позволяет получить подкрепление; (4) не всякий подход к подкрепляемой кормушки приводит к подкреплению, и (5) при отсутствии пищи в лабиринте необходимо выйти в СП. Введение избыточных семантических элементов (двух ложных кормушек) обеспечивали дополнительную когнитивную сложность для распознавании семантической информации, поскольку животные должны были из имеющихся семи семантических элементов найти правильную 4-звенную последовательность – (число размещений из 7 по 4 = 840 возможных комбинаций). Наличие фактора симметрии и возможность коррекции ошибочных действий создавали условия для объективного изучения у животных способности сформировать представление о свойствах предмета (согласно Леонтьеву, 1981) = в зависимости от собственных действий кормушка может изменять свои свойства - быть полной или пустой, оставаясь при этом подкрепляющей в отличие от ложной.
Применение системного подхода к изучению ПД позволило изучить различные проявления ВНД животного: когнитивные, условнорефлекторные, психо-эмоциональные и физиологические в пределах одного эксперимента. При этом оказалось возможным не просто зафиксировать их, а представить в виде целостного описания ПД с учетом общих закономерностей и индивидуальных особенностей. Важно отметить, что традиционный условнорефлекторный и системно-информационный подходы изучения ПД продуктивно дополняли друг друга.