
М. В. Ломоносова биологический факультет на правах рукописи никольская кира Алексеевна системно-информационные аспекты познавательной деятельности позвоночных диссертация
| Вид материала | Диссертация |
| Типология познавательной деятельности (ПД) А –сигнальный типа (I), отказавшийся решать проблемную ситуацию; В |
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- М. В. Ломоносова биологический факультет н а правах рукописи Столяров Андрей Павлович, 657.12kb.
- Самовольцева Кира Алексеевна Дылевич Елена Валерьевна закон, 168.26kb.
- На правах рукописи Гуськова ирина Алексеевна, 682.76kb.
- На правах рукописи, 772.97kb.
- На правах рукописи, 315.69kb.
- На правах рукописи, 372.38kb.
- Тема: «Системно-деятельностный подход - универсальный инструмент познавательной деятельности», 64.45kb.
- Самовольцева Кира Алексеевна Агеева Елена Валерьевна закон, 118.95kb.
- На правах рукописи, 544.87kb.
Типология познавательной деятельности (ПД)
Исследования индивидуальных особенностей ПД животных на примере изучения обучения крыс Вистар (n=200) показали, что в пределах социума, состоящего из 10 -12 особей устойчиво можно было выделить три подгруппы животных в соотношении 10%-30%-60%, различавшихся по характеру поведения в группе, динамике обучения, поведенческой тактики, репертуару психо-эмоциональных проявлений и характеру когнитивных процессов. Оказалось возможным сгруппировать животных не только по признаку "обучается – не обучается ", но и по содержанию – типу познавательной деятельности. По характеру аналитико-синтетической деятельности были выделены три типа, которые мы обозначили как: сигнальный, процедурный и концептуальный.
Наибольшую группу в популяции крыс Вистар (56.8 ± 4.7%) составляли особи, познавательная деятельность которых могла быть определена как сигнальная (тип I), осуществляющаяся по жесткому типу, поскольку они способны были формировать 4-х или 5-звенный навык только по методике жесткого обусловливания. В тоже время, в условиях свободного выбора они отказывались решать сходную пищедобывательную задачу (рис. 6 А) и самостоятельно могли сформировать лишь элементарную форму УР – побежку к кормушке. Высокая чувствительность животных к неопределенности ситуации, проявляющаяся в виде пассивных стрессовых проявлений и двигательной заторможенности в совокупности с подпороговыми значениями вероятности успеха из-за многочисленных ошибочных действий, указывали на значимые дефекты в мотивационной сфере у крыс I типа.
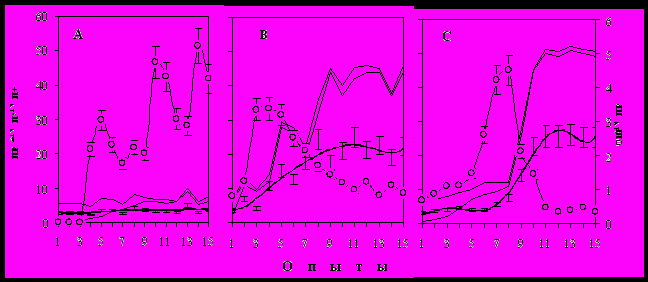 Рис. 6. Условнорефлекторные показатели обучения у крыс Вистар, различающихся по типу познавательной деятельности.
Рис. 6. Условнорефлекторные показатели обучения у крыс Вистар, различающихся по типу познавательной деятельности. А –сигнальный типа (I), отказавшийся решать проблемную ситуацию; В – процедурный, экспоненциальная кривая обучения (II тип) и С – концептуальный (III тип), характеризующийся логистической кривой обучения; серая линия – число посещений лабиринта (проб) в опыте (NПР); черная жирная линия – тренд NПР; тонкая с маркером – число ошибочных попыток в пробе (NОШ/NПР); верхняя тонкая - число подкреплений, доступное животному в пробах (2NП) с учетом двух подкрепляющих кормушек; тонкая линия в основании площади - фактическое число порций пищи, изъятое животным в опыте (NП+); заштрихованная область - несоответствие между доступным и фактическим числом подкреплений.
Совокупность выделенных черт свидетельствовали о подавленном характере ПД. Фармакологические исследования подтвердили правомочность сделанного заключения, поскольку препараты с ноотропными свойствами (опилонг, пирацетам, гепарин) в малых дозах, оказывали активирующее воздействие и в 68 % случаев активные особи смогли обучиться.
В процессе исследовании выяснилось, что определяющую роль в характере поведения крыс I типа играли параметры основных нервных процессов. Наименьшей силой в группе характеризовался процесс возбуждения за счет слабой его выраженности как по времени, так и по его уровню, соотношение возбудительно-тормозных процессов было смещено в сторону значимого преобладания последнего. Максимальные в популяции значения длительности торможения давали основание говорить о том, что в ситуации высокой неопределенности торможение, в основном, выступало, согласно Э.А. Асратяну (1955), с охранительной ролью.
Анализ когнитивной деятельности показал, что особенности крыс с подавленными признаками ПД обусловлены очень низкой скоростью информационного процессинга (табл. 3).
Таблица 3.
Особенности аналитико-синтетической деятельности у крыс Вистар 1

1 – данные приведены: для I типа – 13й опыт, для II – 7й опыт, для IIIА и IIIИ типов непосредственно перед скачком;* - 0.05 и ** - 0.001 достоверные значения по сравнению с типом II в соответствие с критерием t-Стьюдента; # - 0.05 и ## - 0.001 при сопоставлении двух подгрупп крыс III типа.
Крысы сигнального типа обнаруживали серьезные трудности при формировании целостного представления о пространстве. Хотя количество выделенных синтаксических единиц было достаточно для построения модели пространства, низкая интенсивность ассоциирования, была обусловлена как слабой мотивационной активностью, так и повышенной чувствительностью к ситуации выбора. У большинства особей процесс распознавания новой семантической информации, как правило, сопровождался явлением отрицательной интерференции - после выполнения новой семантической связи, животное резко впадало в состояние глубокой двигательной заторможенности, подчас до конца опыта. Такие особенности крыс I типа как: низкая интенсивность ассоциативного процесса за счет пониженной двигательной активности, обусловленной высоким уровнем страха; когнитивные возможности, ограниченные уровнем прогнозирования результата: (если есть сигнал/среда, то будет пища), реализуемые в поведении в виде элементарного УР - побежки к кормушке; способность обучаться сложным формам поведения в условиях жесткого обусловливания позволили нам определить эту группу крыс как животных с сигнальным типом ПД.
Характерные черты поведения крыс процедурного типа (II тип; 11.5 ± 2.7 %) - высокая двигательная и исследовательская активности в незнакомой среде; чрезвычайно низкий уровень пассивно-оборонительных проявлений, высокая социальная активность особей. В условнорефлекторной ситуации они смогли решить предложенную задачу в условиях свободного выбора, кривая обучения развивалась по экспоненциальному закону, познавательная активность носила выраженный (для наблюдателя) характер, поскольку приращение поведенческой эффективности происходило по мере накопления опыта (рис. 6 В). Обучение осуществлялось на фоне высокой вероятности успеха, так как животные с первых проб брали подкрепление в обеих кормушка, несмотря на незнание структуры задачи. Ошибка, как правило, приводила к двигательной активации, которая сопровождалась различными активными психо-эмоциональными проявлениями, провоцируя маршрутную вариабельность. Отличительная особенность ПД этих крыс заключалась в том, что первая правильная и минимизированная ассоциация, соответствующая решению задачи (Вх К1 К2 Вых) у этих животных появлялась уже во втором опыте после М ± m = 12.4 ± 0.03 посещений лабиринта. Несмотря на экспоненциальное нарастание мотивационной активности (МА) и двигательной эффективности (ЭД), приращение организованности навыка (ОН) было затянуто за счет медленного угашения ошибочных действий (рис. 6).
Этап стабилизации характеризовался высокой неустойчивостью осуществления минимизированного решения, последовательное воспроизведение которого не превышало 2-3-х выполнений и наличием различного рода стрессовых и невротических проявлений. Отсутствие процесса оптимизации, высокая вариабельность значений всех УР показателей (КVAR=0.38), низкий уровень организованности навыка (РMIN=0.47) и большая доля активных психо-эмоциональных проявлений свидетельствовали о неустойчивости поведения.
Исследования показали, что низкая организованность навыка у процедурного типа была обусловлена присутствием в поведении достаточно примитивных в когнитивном отношении тактик, таких как ситуационно-обусловленный обход (ориентация по внешним маркерам при осуществлении пищевого поведения), так и более простых вариантов, целенаправленного обхода – повторов, заглядываний и половинчатых решений, доля которых составляла 62 %. Поэтому, несмотря на высокий уровень мотивационной и двигательной активности, навык у этих животных характеризовался высокой хаотичностью как семантической (из-за многочисленных ошибок), так и синтаксической (из-за разнообразия маршрутов). Это обуславливало высокие значения когнитивных затрат на формирование программы поведения, при том, что на процесс восприятия условий задачи протекал наиболее легко у этих крыс (табл. 3).
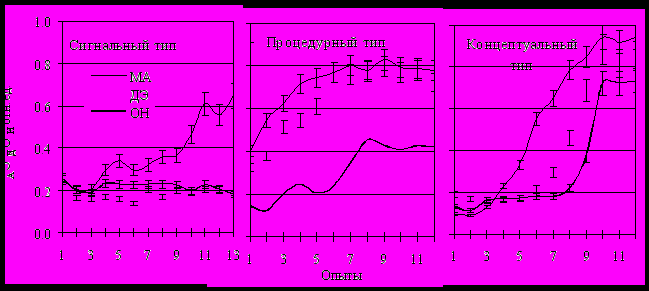
Рис. 6. Интегральные показатели обучения у крыс Вистар, различающихся типом познавательной деятельности.
По вертикали: МА – мотивационная активность, ЭД – условнорефлекторная двигательная эффективность, ОП – организованность навыка. Расчет показателей в разделе Методика.
При оценке основных нервных процессов выяснилось, что крысы с процедурным типом ПД характеризовались наибольшей в популяции выраженностью возбудительного процесса как по времени, так и по его уровню. Значения тормозного процесса, напротив, были наименьшими в популяции, проявляясь в поведении в низком уровне страха в незнакомой среде. Такие признаки как: высокая интенсивность ассоциативного процесса, нарастающая раздражительность, вызванная когнитивной деятельностью, гиперчувствительность к различным внешним стимулам, в том числе и к магнитному фактору, в ответ на действие психостимулирующих фармакологических препаратов указывали на склонность ЦНС этих животных к повышенной возбудимости. Результаты исследования свидетельствовали о том, что особенности основных нервных процессов, их явная неуравновешенность в сторону преобладания возбуждения, соответствуя по классификации Павлова, холерическому типу, обусловливали не только специфику поведения, но и характер ПД и особенности функционирования памяти у крыс II типа. Значимое преобладание возбудительного процесса определяло неустойчивость УР возбуждения как на этапе обучения, так и при воспроизведении памятного следа (рис. 8).
Благодаря системно-информационному подходу удалось установить, что неустойчивость функционирования памяти у крыс процедурного типа была обусловлена тем, что в каждом опыте истории формирования навыка в сжатом виде многократно повторялась (рис. 7). Опыт всегда начинался с извлечения инстинктивной программы (ВхКВых), вслед за этим на фоне нарастающего возбуждения, воспроизводились звенья структуры задачи, которые тут же объединялись в структуру решения. После этого решение распадалось на фрагменты, появлялись ошибки, а затем последовательность операций вновь повторялась.
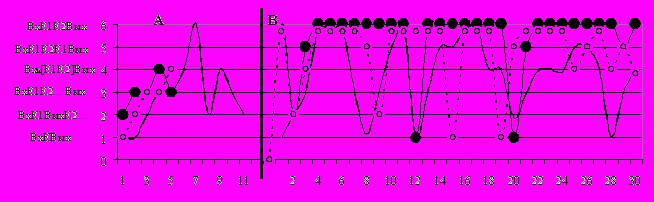 Рис. 8. Примеры динамики воспроизведения навыка на этапе стабилизации.
Рис. 8. Примеры динамики воспроизведения навыка на этапе стабилизации. По горизонтали – пробы, по вертикали – тактика выполнения решения: 1 – половинчатое решение, совпадающее по своей структуре с инстинктивной программой; 2 – ненаправленный обход; 3 – целенаправленный обход двух подкрепляющих кормушек; 4 – повтор (многократные повторные заглядывания в пустые кормушки); 5– однократное заглядывание в кормушку; 6 – минимизированный обход; сплошная линия – тип II, с черными маркерами – тип IIIА, пунктир – тип IIIИ. На рис. А – начальный этап обучения: у типа II – опыт 2, у типа IIIА – опыт 4, у типа IIIИ – опыт 6. На рис. В – этап стабилизации навыка (14-й опыт).
Когнитивные трудности крысы II типа испытывали не только при обучении, но и при переделке сформированного навыка. Благодаря этой пробе удалось установить, что ПД животных не выходила за рамки процедурного понимания задачи. Несмотря на то, что в «истории» обучения животного сходная комбинация действий ранее осуществлялся с вероятностью М ± m = 0.13 ± 0.03, процесс изменения исходного значения на новое требовал «нового» обучения, проходил поэтапно по принципу последовательного перемещения «с конца» нового семантического элемента к начальному звену навыка.
Анализ когнитивной деятельности свидетельствовал о высоких ассоциативных возможностях крыс II типа, позволяющих животному быстро овладеть незнакомым пространством и свободно ориентироваться в нем (табл. 3). Несмотря на это, интегративный процесс протекал намного труднее, поскольку "истинное" распознавание структуры задачи, которое имело бы признаки поведенческой программы, происходило чрезвычайно медленно. Устойчивое неслучайное ее воспроизведение всегда было существенно отставлено от его первой реализации. Несоответствие между ассоциативными и интегративными процессами наиболее четко выявилось при психостимулирующих воздействиях на организм, поскольку под их влиянием усиливалось рассогласование между ассоциативными и интегративными процессами: облегчение ассоциативного процесса сочеталось с угнетением интегративного. Полученные в работе факты указывали на то, что когнитивный процесс у этих животных заключался в увеличении исходной информационной емкости единицы информации только до уровня отдельных звеньев (4-8-граммы), представляющие собой УР 1-го порядка. Реализация программы целенаправленного поведения у крыс этого типа протекала по механизму принятия решения в рамках оперативной памяти. Анализ поведения показал, что для процедурного типа организация цепного УР происходила с опорой на наличный стимул. Каждое условнорефлекторное действие привязывалось к конкретному контекстному стимулу среды - пустой кормушке (44%), выходу или ложной кормушке (12%). Включение их в структуру решения позволяло организовать поведение на более простом в когнитивном отношении уровне, поскольку структура задача допускала решать ее не только на основе прогнозирования результата действия, т.е. установления причинно-следственной связи (УР 2-го порядка) – S1 → S2 , но и на основе прогнозирования действия: S →R.
Таким образом, такие особенности крыс II типа как: (1) высокие ассоциативные возможности за счет одновременного образования множества ассоциаций (сенситизация) благодаря высокой двигательной активности и низкой тревожности; (2) низкие прогностические способности за счет одинаковой значимости всех связей, независимо от их прагматической ценности; (3) экспоненциальное нарастание эффективности поведения за счет быстрого перебора всех связей, провоцирующих в случае ошибки психомоторную активацию (обучение по методу проб и ошибок); (4) реализация программы поведения на основе прогнозирования каждого последующего действия, входящего в структуру задачи, позволили нам определить эту группу крыс как животных с процедурным типом ПД.
В третью группу были отнесены крысы, для которых был характерен концептуальный (компилятивный) тип ПД (III тип, 29.8 ± 1.3 %). В условиях высокой неопределенности они проявляли пассивно-оборонительные черты, хотя степень их выраженности была меньше, чем у крыс I типа. Самостоятельный выход из клетки содержания в «жилую комнату» был отставлен на 7-10 мин от начала опыта и происходил только после ее посещения крысами II типа. В условнорефлекторной ситуации крысы III типа смогли решить предложенную задачу, кривая обучения, в отличие от процедурного типа, развивалась по логистическому закону (рис. 6 С). Медленное приращение эффективности поведения (по числу проб в опыте) в течение 5-9 опытов (этап плато) и последующий резкий выход на высоко организованное и стереотипное поведение (этап скачка) свидетельствовало о накопительном характере научения. Познавательная деятельность носила явно скрытый характер, поскольку никакие видимые (для наблюдателя) поведенческие признаки не указывали на возможность скачка, это всегда происходило неожиданно для экспериментатора. Ошибочные действия, в отличие от процедурного типа, провоцировали всегда тормозный эффект – последующее за ошибкой непродолжительное подавление или снижение двигательной активности М ± m = 0.42 ± 0.03 мин. Однако угашение ошибочных действий происходило очень быстро (рис. 6 С).
При сходстве основных специфических черт обучения, группа III типа оказалась неоднородной по выраженности рефлекса осторожности, психо-эмоциональному рисунку и длительности скрытого периода. В первую подгруппу (IIIА - 35.7%) вошли активные животные, у которых продолжительность скрытого периода не превышала М ± m = 6.6 ± 0.1 опытов, пассивно-оборонительные черты, типичные на начальном этапе обучения, быстро сменялись активными проявлениями, индуцированные когнитивным процессом. Во вторую подгруппу (IIIИ - 64.3% ) вошли крысы с инертными фенотипическими проявлениями, у которых продолжительность скрытого периода была достоверно больше - М ± m = 8.8 ± 0.1 опытов за счет преобладания рефлекса осторожности в виде различных пассивных проявлений, на фоне которых осуществлялась ПД. Реализация найденного решения задачи в поведении сопровождалась более резким, чем в подгруппе IIIА возрастанием его эффективности (dNПР/NОП – 9.72 и 7.15 соответственно) не только в отношении организованности (уровня оптимизации), но и локомоторной активности. Значения когнитивных затрат позволяли говорить о том, что процесс преобразования распознанной информации в программу поведения у инертных животных протекал легче (табл. 3).
Анализ когнитивной деятельности показал, что интенсивность ассоциативного процесса у крыс III типа находилось в прямой зависимости от того, как быстро у крыс снимались ограничения в отношении исследовательской активности (возбудительного процесса), обусловленные рефлексом осторожности. Поэтому накопление различных информационных единиц в виде двигательных последовательностей, их усложнение, а также распознавание элементов структуры задачи происходило последовательно (табл. 3) и не сопровождалось явлением сенситизации. Процессы распознавания различных видов информации протекали синхронно и подчинялись логистической закономерности, в то время как у II типа они осуществлялись последовательно. Своеобразие когнитивной деятельности у концептуального типа заключалось в том, что накопление новых единиц, увеличение их информационной емкости, так же как и переход на более высокий уровень обобщения происходили скачкообразно. Этому всегда предшествовал период многократного апробирования в пределах одной пробы раннее выделенных единиц, причем признак «навязчивого» повторения был присущ исключительно животным III типа и встречался на всех этапах ПД. Анализ символьной записи поведения животного позволил вскрыть содержательную сторону аналитико-синтетических процессов и сущность скрытого характера обучения. Выяснилось, что после относительно длительного периода накопления первичной информации о значимых узлах пространства (2х- и 3х-граммы, табл. 3) фрагментарные «знания» скачкообразно объединялись в единую замкнутую структуру типа семантической сети.
Результаты исследования показали, что ошибочная информация у концептуального типа играла важную, определяющую роль при выяснении значимости семантических связей, вследствие чего обучение осуществлялось, в отличие от процедурного типа, на фоне пороговых значений вероятности успеха (0.3). Анализ динамики становления структуры решения свидетельствовал о том, что изначально аналитико-синтетический процесс был ориентирован на формальные признаки структуры задачи (схему) – построение различных 4-звенных последовательностей, независимо от биологической значимости элементов, входящих в эту структуру. Равновероятное осуществление различных комбинаций, в том числе и все варианты с ложными кормушками, как в пределах одной пробы, так и опыта обеспечивало условия для осуществления в оперативном режиме процесса сравнения и последующего выбора адекватной комбинации.
Высокий уровень афферентной генерализации в этот период коррелировал с низкой вероятностью физического подкрепления, которая увеличивалась как только поведение приобретало признаки устойчивой целенаправленности. Важная роль сенсорного подкрепления на этапе восприятия информации свидетельствовала о том, что реализация аналитико-синтетического процесса осуществлялась на основе УР 1-го порядка, обеспечивая возможность образования каузального УР – УР 2-го порядка. Одна из отличительных особенностей ПД концептуального типа состояла в том, что результат когнитивной деятельности мог быть реализован в поведении в виде устойчивого целенаправленного навыка только после того, когда не только была распознана структура задачи (семантика), но и найден конкретный оптимальный вариант ее двигательной реализации (рис. 6С). Показатель непрерывного пробега на этапе завершения обучения, значения которого соответствовали длине решения (М ± m = 11.3 ± 0.25), отражал способность этих животных делать долгосрочный прогноз относительно результата будущих действий, по крайней мере на 3 - 4 звена вперед.
Таким образом, концептуальный тип, уступая процедурному типу в отношении когнитивных затрат, потребовавшихся на распознавание информации (табл. 3), оказывался более эффективным в количественном и качественном отношении относительно использования этой информации для организации целенаправленного поведения (навыка). Правомочность сделанного предположения была подтверждена данными, полученными в тесте по переделке информационного значения кормушек. Процесс переделки для крыс III типа состоял в перепроверке ранее сформированного правила поведения за счет извлечения из памяти различных семантических комбинаций, в том числе и с ложными кормушками. Быстрый переход к новому варианту навыка (один опыт) отражал, по существу, концептуальное понимание задачи: независимо от мест получения пищи, правило поведения заключалось в смене двух информационно различных сред: лабиринт (пищевая) ↔ свободное поле (непищевая).
Проведенные исследования позволили установить прямую зависимость когнитивной деятельности и особенностей поведения III типа от характера основных нервных процессов. В целом, в отличие от процедурного типа, животные характеризовались большой силой возбудительно-тормозных процессов и их относительной уравновешенностью. При этом у активных крыс возбуждение преобладало над торможением, в то время как у особей с инертными проявлениями оно было смещено в противоположную сторону. Внутригрупповые особенности основных нервных процессов отражались на характере разворачивания исследовательской активности и угашения ошибочных действий. По этим показателям активные крысы III типа занимали промежуточное положение между возбудимыми (II тип) и инертными (IIIИ тип) животными, проявляя сходную с первыми высокую исследовательскую активность, а со вторыми – большую скорость угашения ошибок, что позволяло, в соответствии с павловской классификацией, определить их как «сангвиники». Крайне медленное и постепенное разворачивание исследовательской активности в сочетании со способностью очень быстро вытормаживать ошибочные действия позволили нам подгруппу крыс IIIИ типа определить как «флегматики».
При сходстве основных черт обучения крысы III типа наиболее значимо различались по показателю устойчивости воспроизведения навыка (рис. 8). Немаловажную роль в работе механизмов, обеспечивающих длительное поддержание уровня внимания (концентрацию УР возбуждения) на этапе стереотипного осуществления навыка, как оказалось, играл такой фактор как направленность неуравновешенности нервных процессов. У «сангвиников» устойчивость навыка в константных условиях оказывалась низкой - не более 5 реализаций минимизированных решений подряд, а у «флегматиков» при сходном с «сангвиниками» уровне УР возбуждения (NПР = 27.4 ± 0.18; М ± m) устойчивость последнего была высокой (до 8 -12 минимизированных решений подряд, а в ряде случаев до 20-24 раз) Особенности нервных процессов определяли характер функционирования памяти у крыс III типа, который принципиально отличался от такового у процедурного типа (рис. 8). Достоверно более высокие значения готовности, мобилизационной активности и устойчивости памятного следа, а, главное, факт воспроизведения минимизированного решения с первой пробы в опыте указывали на то, что реализация решения у всех крыс III типа осуществлялась по механизму извлечения целостной программы поведения из долговременной памяти. Факт независимости эффективности работы животных обеих групп от их внутреннего состояния, выявленный в пробе на сытость, подтверждал правомочность высказанного предположения.
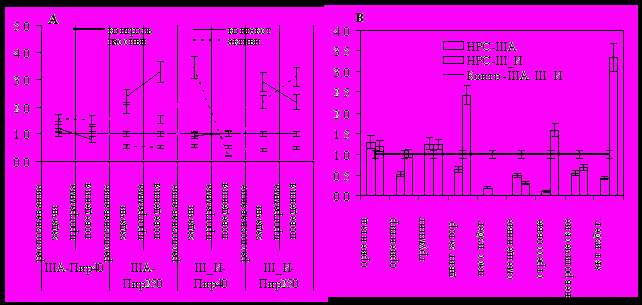
Рис. 9. Особенности психо-эмоциональных проявлений у III типа, подвергнутого фармакологическому воздействию (А) и разрушению дорсального НРС (В).