
Лекция № клетка элементарная биологическая система. Структурно функциональная организация про- и эукариотических клеток
| Вид материала | Лекция |
- Нейроны и глиальные клетки: общая характеристика, разнообразие, функции. Серое и белое, 2972.29kb.
- Способы деления эукариотических клеток: митоз, мейоз, амитоз, 51.33kb.
- Структурно-функциональная организация палеоамигдалы: фундаментальные закономерности, 2391.63kb.
- Реферат по курсу «Анатомия центральной нервной системы» Тема: «Строение нейрона и нервного, 92kb.
- Повторение изучаемого, 127.15kb.
- Цитоплазма: а органоиды, 266.57kb.
- Клеточная инженерия Клетка, 255.38kb.
- Основы цитологии, 262.7kb.
- Программа по биологии для абитуриентов, поступающих в игму в 2010 г. Основы цитологии, 370.13kb.
- Вантеев А, 865kb.
ЛЕКЦИЯ № 2. КЛЕТКА – ЭЛЕМЕНТАРНАЯ БИОЛОГИЧЕСКАЯ СИСТЕМА. СТРУКТУРНО – ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПРО- И ЭУКАРИОТИЧЕСКИХ КЛЕТОК.
Клетка – основная структурно – функциональная единица всех живых организмов, элементарная живая система. Клетка может существовать как отдельный организм (бактерии, простейшие, некоторые водоросли и грибы) или в составе тканей многоклеточных животных, растений, грибов. Лишь вирусы представляют собой неклеточные формы жизни, способные осуществлять свой жизненный цикл только внутри клеток хозяина. Представление о клетке как элементарной структуре живых организмов, известное как клеточная теория, сложилось постепенно в XIX в. в результате микроскопических исследований.
- ^ Клеточная теория.
Клеточная теория – это обобщенные представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов.
Появлению и формулированию отдельных положений клеточной теории предшествовал длительный (более трехсот лет) период накопления наблюдений над строением различных одноклеточных и многоклеточных организмов растений и животных. Этот период связан с развитием и усовершенствованием различных оптических методов исследования.
Клеточная теория была сформулирована ботаником М. Шлейденом и зоологом Т. Шванном в 1838-1839 г.г. В 1858 г. Р. Вирхов обосновал принцип преемственности клеток путем деления («каждая клетка из клетки»). Создание клеточной теории стало важнейшим событием в биологии, одним из решающих доказательств единства живой природы.
Клеточная теория постулирует:
- Клетка – элементарная единица живого;
- Клетки разных организмов гомологичны по своему строению;
- Размножение клеток происходит путем деления исходной клетки;
- Многоклеточные организмы представляют собой сложные ансамбли клеток, объединенные в целостные, интегрированные системы тканей и органов, подчиненных и связанных между собой межклеточными, гуморальными и нервными формами регуляции.
С современных позиций можно добавить еще одно положение:
- В клетке содержится вся генетическая информация о строении и функциях организма.
- ^ Клетка – элементарная единица живого.
Живому свойственен ряд совокупных признаков, таких, как способность к воспроизведению (репродукции), росту, использование и трансформация энергии, метаболизм (ассимиляция и диссимиляция), возбудимость, раздражимость, изменчивость и др. Такую совокупность признаков можно обнаружить на клеточном уровне. Нет меньшей единицы живого, чем клетка. Можно выделить из клетки отдельные ее компоненты или молекулы и убедиться, что многие из них обладают специфическими функциональными особенностями, но только клетка в целом является наименьшей единицей, обладающей всеми свойствами живого.
- ^ Клетки разных организмов гомологичны по своему строению.
Термин «гомологичность» обозначает сходство по коренным свойствам и отличие по второстепенным. Гомологичность строения клеток наблюдается внутри каждого из типов клеток: прокариотическом и эукариотическом. Хорошо известно разнообразие клеток как бактериальных, так и высших организмов. Такое одновременное сходство строения и разнообразие форм определяется тем, что клеточные функции можно подразделить на две группы: обязательные и факультативные. Обязательные функции, направленные на поддержание жизнеспособности самих клеток, осуществляются специальными внутриклеточными структурами, сходными у разных типов клеток.
Различие клеток связано со специализацией их функций, с развитием особых клеточных аппаратов (например, фибриллярные компоненты в мышечных клетках, тигроид и отростки со специальными структурами передачи нервного импульса (синапс)).
- ^ Размножение клеток происходит путем деления исходной клетки.
Формулировка этого положения связана с именем Р. Вирхова. Размножение клеток прокариотических и эукариотических организмов происходит только путем деления исходной клетки, которому предшествует воспроизведение ее генетического материала (редупликация ДНК).
- Многоклеточные организмы представляют собой сложные ансамбли клеток, объединенные в целостные, интегрированные системы тканей и органов, подчиненных и связанных между собой межклеточными, гуморальыми и нервными формами регуляции
Действительно, клетка – это единица функционирования в многоклеточном организме. Но клетки объединены в функциональные системы, в ткани и органы, которые находятся во взаимной связи друг с другом. Специализация частей многоклеточного организма, расчлененность его функций, дают ему большие возможности приспособления для размножения отдельных индивидуумов, для сохранения вида.
- ^ В клетке содержится вся генетическая информация о строении и функциях организма.
Этот постулат появился после изучения строения и функций ДНК, которая является носителем генетической информациии клетки.
II. Сравнение прокариотических и эукариотических клеток.
В природе существует значительное разнообразие конкретных клеточных форм. Вместе с тем число основных типов клеточной организации ограничено. Выделяют прокариотический и эукариотичекий типы с подразделением второго на подтип клеток простейших организмов и подтип многоклеточных.
Клетки прокариотического типа имеют особенно малые размеры – не более 0,5-5,0 мкм в диаметре. Содержимое прокариотической клетки одето плазматической мембраной, играющей роль активного барьера между собственно цитоплазмой клетки и внешней средой. Обычно снаружи от плазматической мембраны расположена клеточная стенка или оболочка – продукт клеточной активности. У них нет морфологически обособленного ядра, так как ядерный материал в виде ДНК не отграничен от цитоплазмы оболочкой. Генетический аппарат образован единственной кольцевой хромосомой, которая лишена основных белков – гистонов (нуклеоид). В клетке отсутствует развитая система мембран, хотя некоторые виды бактерий (например, фототрофные пурпурные бактерии) богаты внутриклеточными мембранными системами. Очень сильно цитоплазматические мембраны развиты у сине-зеленых водорослей. В основном веществе (или матриксе) цитоплазмы прокариотических клеток располагаются многочисленные рибосомы. У прокариот отсутствует клеточный центр. Для них не типичны внутриклеточные перемещения цитоплазмы и амебоидное движение. Время, необходимое для образования двух дочерних клеток из материнской, сравнительно мало и исчисляется десятками минут. Прокариотические клетки не делятся митозом. К этому типу клеток относятся бактерии и сине-зеленые водоросли.
Эукариотический тип клеточной организации представлен двумя подтипами: клетки простейших и клетки многоклеточных организмов (растительные и животные). Особенностью организмов простейших является то, что они, исключая колониальные формы, в структурном отношении представляют собой клетку, а в физиологическом – целый организм. В связи с этим в клетках некоторых простейших имеются миниатюрные образования, выполняющие на клеточном уровне функции органов, аппаратов и систем органов многоклеточного организма (цитостом, цитофарингс, порошица, сократительные вакуоли, генеративное и вегетативные ядра инфузорий). Основным отличительным признаком эукариотических клеток является наличие морфологически выраженного ядра. Кроме того, в цитоплазме таких клеток существует целый набор специальных структур, органелл, выполняющих отдельные специфические функции. К числу органелл относят мембранные структуры: ядро, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, митохондрии, пластиды (в клетках растений). Кроме того, для эукариотических клеток характерно наличие немембранных структур, таких, как центриоли (в клетках животных), рибосомы, микротрубочки, микрофиламенты и др. Эукариотические клетки обычно намного крупнее прокариотических (Таблица 1).
ТАБЛИЦА 1.^ Типы клеточной организации.
| Признак | Прокариоты | Эукариоты |
| 1. Размеры клеток | Диаметр 0,5 – 5 мкм | Диаметр примерно 40 мкм. Объем клетки в 1000-10000 раз больше, чем у прокариот. |
| 2. Ядро | нет | есть |
| 3. Ядерная мембрана | нет | есть |
| 4. Генетический аппарат | Одна кольцевая хромосома в зоне нуклеоида | Хромосомы |
| 5. Система цитоплазматических мембран | нет | есть |
| 6. Эдоплазмати-ческий ретикулум (ЭПР) | нет | есть |
| 7. Рибосомы | есть (70S) | есть (80S) |
| 8. Митохондрии | нет | есть |
| 9. Комплекс Гольджи | нет | есть |
| 10.Лизосомы | нет | есть |
| 11.Клеточный центр | нет | есть |
| 12.Микротрубочки | нет | есть |
| 13. Внутриклеточное перемещение цитоплазмы | нет | есть |
| 14. Органоиды движения | Жгутики. Жгутиковая нить состоит из белка флагеллина. | Реснички и жгутики включают в свой состав микротрубочки, построенные из белка тубулина. |
| 15. Наружная клеточная мембрана | есть | есть |
| 16. Клеточная стенка | Жесткая, содержит полисахариды, основной – муреин (пептидогликан) | У растений содержит целлюлозу, у насекомых и грибов – хитин, у животных отсутствует. |
| 17. Деление | Прямое | Митоз, мейоз |
^ Ш. Химический состав клетки.
- Клетки живых организмов сходны не только по своему строению, но и по химическому составу. Сходство в строении и химическом составе клеток свидетельствует о единстве их происхождения.
По составу входящие в клетку вещества делятся на органические и неорганические.
^ II. 1. Неорганические вещества.
На первом месте по массе в клетке стоит вода (примерно 2/3 массы клетки). Вода имеет огромное значение в жизнедеятельности клетки. Многие элементы в клетках содержатся в виде ионов. Чаще всего встречаются катионы: K+, Na+, Ca2+ Mg2+, и анионы: H2PO4-, Cl-, HCO3-. Содержание катионов и анионов в клетках обычно значительно отличается от содержания их во внеклеточной среде.
Минеральные соли (например фосфат кальция) могут входить в состав межклеточного вещества, раковин моллюсков и обеспечивать прочность этих образований.
^ III.2. Органические вещества.
Характерны только для живого. Органические соединения представлены в клетке простыми малыми молекулами (аминокислоты, моно- и олигосахариды, жирные кислоты, азотистые основания), и макромолекулами биополимеров (белки, липиды, полисахариды, нуклеиновые кислоты). Молекулы биополимеров состоят из повторяющихся низкомолекулярных соединений (мономеров), ковалентно связанных между собой.
IV. Строение эукариотической клетки.
Все эукариотические клетки имеют общий план строения. Собственно тело клетки и ее содержимое отделены от внешней среды или от соседних элементов у многоклеточных организмов плазматической мембраной. Кнаружи от плазматической мембраны расположена клеточная оболочка или стенка, особенно хорошо выраженная у растений. Все внутреннее содержимое клетки, за исключением ядра, носит название цитоплазмы. Цитоплазма эукариотических клеток не однородна по своему строению и составу и включает в себя: гиалоплазму, мембранные и немембранные компоненты. Мембранные органеллы представлены двумя вариантами: одномембранные и двумембранные. К первым относятся органеллы вакуолярной системы – эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы и другие специальзированные вакуоли, а также плазматическая мембрана. К двумембранным органеллам относятся митохондрии и пластиды, а также клеточное ядро. К немембранным органеллам принадлежат рибосомы, клеточный центр животных клеток, а также элементы цитоскелета (микротрубочки и микрофиламенты).
4.1. Цитоплазма.
4.1.1. Гиалоплазма.
Термин гиалоплазма (от hyalihe – просвечивающийся, прозрачный), основная плазма или матрикс цитоплазмы, обозначает очень важную часть клетки, ее истинную внутреннюю среду. Гиалоплазма является сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые кислоты, полисахариды и т.д. В ней локализованы ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, в метаболизме сахаров. Эта система способна переходить из золеобразного (жидкого) состояния в гель и обратно. Важнейшая роль гиалоплазмы заключается в том, что эта среда объединяет все клеточные структуры и обеспечивает химическое взаимодействие их друг с другом. Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов: перенос аминокислот, жирных кислот, нуклеотидов, сахаров. В гиалоплазме идет постоянный поток ионов к плазматической мембране и от нее, к митохондриям, ядру и вакуолям. Гиалоплазма является основным вместилищем и зоной перемещения массы молекул АТФ. В гиалоплазме происходит отложение запасных продуктов: гликогена, жира. В цитозоле на расположенных там рибосомах синтезируются белки, транспортируемые в разные участки клетки, а также все белки клеточного ядра, большая часть белков митохондрий и пластид, основные белки пероксисом.
^ 4.1.2. Структура клеточных мембран.
Общей чертой всех мембран клетки (плазматической, внутриклеточных и мембранных органоидов) является то, что они представляют собой тонкие (6-10 нм) пласты липопротеидной природы (липиды в комплексе с белками), замкнутые сами на себя. В клетке нет открытых мембран со свободными концами. Мембраны клетки всегда ограничивают полости или участки, закрывая их со всех сторон и тем самым отделяя содержимое таких полостей от окружающей их среды (компартментализация).
Структурной основой мембраны является двойной слой липидов. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях (липофильность). Состав липидов, входящих в мембраны клеток, очень разнообразен. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды, сфингомиелины и холестерин (в растительных клетках не обнаружен). Характерной особенностью липидов мембран является разделение их молекулы на две функционально различные части: неполярные хвосты, состоящие из жирных кислот, и заряженные полярные головки. Обязательным компонентом клеточных мембран являются белки. В среднем они составляют 50% массы мембраны. Но количество белков в разных мембранах может быть различным. Так, в мембранах митохондрий на долю белков приходится около 75%, а в плазматической мембране клеток миелиновой оболочки – около 25%.
Сущесвуют три важных принципа строения мембраны:
- ^ Мембраны не однородны. Мембраны, окружающие внутриклеточные органеллы, и плазматическая мембрана отличаются по составу.
- Многие компоненты мембран находятся в состоянии непрерывного движения. Мембрана напоминает постоянно меняющуюся мозаику.
- ^ Компоненты мембран чрезвычайно асимметричны. Между наружным и внутренним слоями мембран имеется различие по относительному количеству и качественному составу липидов. Белки располагаются среди липидов асимметрично и имеют хорошо различимые вне- и внутриклеточные участки.
Важнейшими функциями мембран являются следующие:
- ^ Мембраны контролируют состав внутриклеточной среды. Основная функция мембраны – формирование вокруг цитоплазмы барьера, который избирательно пропускает молекулы, входящие в клетку и выходящие из нее. В значительной степени такое поведение мембраны обусловлено непроницаемостью ее липидов для воды и других гидрофильных молекул. В мембране находятся белки, которые образуют каналы и поры, принимающие участие в высоко избирательном транспорте молекул через мембрану.
- ^ Мембраны обеспечивают и облегчают межклеточную и внутриклеточную передачу информации. Мембрана – это место, где молекулярная информация воспринимается, преобразуется и передается далее в клетку.
- ^ Мембраны обеспечивают образование тканей с помощью межклеточных контактов. Многие белки, погруженные в мембрану, ковалентно связаны с углеводами (гликопротеины), расположенными на наружной поверхности мембраны. Гликопротеины и гликолипиды важны для осуществления межклеточных контактов.
В 1972 г. Сингер и Николсон предложили жидкостно – мозаичную модель мембраны, согласно которой белковые молекулы не располагаются сплошной пленкой на поверхности билипидного слоя, а образуют в нем своеобразную мозаику. Выявлено два типа мембранных белков. Белки первого типа, называемые периферическими белками, связаны с мембраной в основном ионными взаимодействиями. Примеры периферических белков – фибронектин (локализован на наружной поверхности большинства клеток, исключая циркулирующие клетки крови; обеспечивает взаимодействие между клетками и внеклеточным матриксом, обеспечивая таким образом контакт между внутриклеточной и окружающей средой) и спектрин (находится на внутренней поверхности большинства клеточных мембран, особенно в эритроцитах; связываясь с актином тропомиозином образует цитоскелет мембраны и поддерживает двояковогнутую форму эритроцита). Мембранные белки второго типа называют интегральными белками . Эти протеины или погружены в толщу липидного бислоя, или пронизывают мембрану насквозь (трансмембранные белки). Интегральные белки перемещаются в плоскости мембраны.
По биологической роли мембранные белки можно разделить на три группы: ферменты, рецепторные белки и структурные белки. Набор ферментов в составе мембран может быть очень велик и разнообразен. В разных мембранах существует характерный набор ферментов. Например, в плазматической мембране, как и во многих других, локализуется (К+/Na)-зависимая АТФаза., участвующая в транспорте ионов. В митохондриях специфическими являются набор белков – переносчиков электронов и фермент АТФ-синтетаза, обеспечивабщий окислительное фисфорилирование и синтез АТФ.
Рецепторные белки специфически связываются с теми или иными веществами и узнают их. Это белки-рецепторы для гормонов, для узнавания поверхности соседних клеток, вирусов и др. Рецепторные белки входят в состав любых мембран. Так, на внешней мембране митохондрий расположены рецепторы, участвующие в узнавании и транспорте митохондриальных белков, переносимых из цитозоля в митохондрии. На мембранах эндоплазматического ретикулума находятся рецепторы, узнающие и связывающие рибосомы и т.д. На плазматической мембране расположены как рецепторы, узнающие соседние клетки или даже отдельные ионы солей (переносчики), так и белки, узнающие белки цитоскелета в цитоплазме.
Со стороны цитоплазмы мембраны связаны через примембраные или собственно мембранные интегральные белки с разнообразными белковыми структурами цитоплазмы. К ним относятся в первую очередь компоненты цитоскелета. Это позволяет не только сделать мембраны более жесткими, но и обеспечивает подвижность мембран, создавая возможности для их транспортных функций. Например, жесткость плазматической мембраны безъядерных эритроцитов создается за счет связывания сети цитоплазматических белков с интегральными белками плазмалеммы. В ее состав входит белок, который обеспечивает транспорт ионов через бислой, но одновременно через ряд белков связывается с сетью белков-спектринов, которые создают жесткую подмембранную сеть. Во многих эпителиальных клетках специальные белки плазматической мембраны связываются с элементами цитоскелета и участвуют в образовании целого ряда межклеточных соединений (десмосомы, адгезивный контакт и др.). С элементами цитоскелета связаны также оболочки клеточного ядра: внешняя ядерная мембрана тесно ассоциирована с промежуточными филаментами, которые фиксируют ядро в объеме цитоплазмы. Внутриклеточные вакуоли могут перемещаться в клетке только при взаимодействии с фибриллярными компонетами, такими как микротрубочки и микрофиламенты. Митохондрии перемещаются в клетке также за счет ассоциации с элементами цитоскелета.
Особое место среди клеточных мембран занимает плазматическая мембрана или плазмалемма. Плазматической мембраной называется барьер, который окружает цитоплазму, определяя границы клетки. Это поверхностная периферическая структура, ограничивающая клетку снаружи. Она имеет толщину около 10 нм и представляет собой самую толстую из клеточных мембран. Большая ее толщина обусловлена тем, что на ее внутренней стороне локализован довольно плотный слой периферических белков, а на наружной – слой углеводных компонентов. Плазматическая мембрана выполняет целый ряд важнейших клеточных функций, ведущими из которых являются функция разграничения веществ цитоплазмы от внешней среды и функции транспорта различных веществ как внутрь клетки, так и из нее. Различают активный и пассивный типы транспорта. К пассивным механизмам относят диффузию, облегченную диффузию и осмос, к активным – работу белковых ионных насосов, эндоцитоз и экзоцитоз. Кроме того, плазматическая мембрана участвует в выведении из клеток продуктов, образованных в клетке. На поверхности плазмалеммы располагаются различные рецепторные структуры, специфически взаимодействующие с внеклеточными факторами и с соседними клетками (передача сигналов внутрь клетки). Плазматическая мембрана принимает участие в межклеточных взаимодействиях у многоклеточных организмов (простой контакт, зубчатый контакт, десмосома, плотный контакт, щелевой контакт). Отдельные участки плазматической мембраны в специализированных клетках животных принимают участие в построении специальных отростков клетки, таких, как микроворсинки, реснички, рецепторные выросты и др. Кроме того, плазматическая мембрана играет важную роль при делении клетки.
^ 4.1.3.Эндоплазматический ретикулум (ЭПР).
Строение. ЭПР представляет собой систему уплощенных мембранных мешочков, канальцев, цистерн, пузырьков. Вся сеть объединена в единое целое с наружной мембраной ядерной оболочки и наружной клеточной мембраной. Выделяют два типа ЭПР: гранулярный (шероховатый) и агранулярный (гладкий).
Мембраны гранулярного эндоплазматического ретикулума со стороны гиалоплазмы покрыты мелкими (около 20нм) темными, округлыми гранулами. Эти гранулы представляют собой рибосомы. На мембранах рибосомы расположены в виде полисом (множество рибосом, объединенных одной информационной РНК).
Функции. Наличие полисом на мембранах однозначно показывает на то, что гранулярный ЭПР является важным местом синтеза белков. Но с элементами гранулярного ЭПР связан не весь синтез белка в клетке, а только его часть. Большое количество полиосом располагаются в гиалоплазме. Было показано, что активные в отношении белкового синтеза специализированные , переставшие делиться клетки (например, клетки поджелудочной, молочной желез, нервные клетки и др.) содержат очень мало свободных полисом, большая часть их рибосом связана с мембранами гранулярного ЭПР. Из этих наблюдений был сделан вывод, рибосомы в составе полисом гиалоплазмы синтезируют белки, необходимые для собственно клеточных нужд, для поддержания и обеспечения жизнедеятельности данной клетки. Рибосомы же, связанные с мембранами ЭПР, участвуют в синтезе белков, выводимых из данной клетки (клетки поджелудочной железы синтезируют и выделяют ферменты, участвующие в расщеплении пищи; слюнной железы – пищеварительные ферменты амилазу и др, молочные железы – казеин молока и т.д.). Многие синтезируемые ферменты при свободном выходе в гиалоплазму могут привести к самоперивариванию клетки (автолиз), к ее гибели. Однако этого не происходит, потому что сразу после синтеза белки переносятся через мембрану ЭПР в полость вакуолей и тем самым изолируются от содержимого гиалоплазмы и цитоплазматических структур. Таким образом, роль гранулярного ЭПР заключается не просто в участии в синтезе белков на рибосомах его мембран, но и в процессе сегрегации, обособления этих синтезированных белков клетки. Кроме того, многие секретируемые белки претерпевают различные изменения, «созревают» в вакуолях гранулярного ЭПР. Важнейшей функцией гранулярного ЭПР является способность синтезировать интегральные и периферические белки мембран, липидный компонент мембран и осуществлять сборку липопротеидных мембран.
^ Гладкий ЭПР представляет собой часть мембранной ретикулярной системы. В морфологическом отношении он также представлен мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярного на мембранах гладкого ЭПР нет рибосом. Чаще всего канальцы гладкого ЭПР образуют скопления или зоны. Гладкий и гранулярный ЭПР связаны между собой и переходят друг в друга. Считается, что гладкий ЭПР образуется из шероховатого путем потери рибосом. Несмотря на топографическую связь и вероятную общность происхождения, эти два представителя ЭПР резко отличаются друг от друга в функциональном отношении.
Функцией гладкого ЭПР является метаболизм липидов и некоторых внутриклеточных полисахаридов
4.1.4. Аппарат Гольджи (комплекс Гольджи, пластинчатый комплекс).
Строение. Как было показано выше, синтез и накопление многих выделяемых из клеток веществ, таких как белки, липопротеиды, стероиды, происходит в элементах эндоплазматического ретикулума. Однако выведение их за пределы клетки (экзоцитоз) происходит с участием другой мембранной системы – аппарата Гольджи. Комплекс Гольджи представляет собой скопление мембранных структур, собранными в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. Плоские мембранные мешки (цистерны) располагаются в виде стопки. Между стопками располагаются тонкие прослойки гиалоплазмы. В центре мембраны цистерн сближены, а на периферии могут иметь расширения (ампулы). Кроме плотно расположенных плоских цистерн в зоне аппарата Гольджи наблюдается множество вакуолей. Мелкие вакуоли отшнуровываются от расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный, или формирующийся цис-участок и дистальный, или зрелый, транс-участок. В секретирующих клетках аппарат Гольджи обычно поляризован: с одной стороны мембранные мешочки непрерывно образуются, а с другой – отшнуровываются в виде пузырьков. Цистерны аппарата Гольджи связаны с канальцами ЭПР. В некоторых клетках аппарат Гольджи имеет вид сложных сетей.
Функции. 1. Секреторная функция аппарата Гольджи.
Мембранные элементы аппарата Гольджи участвуют в сегрегации и накоплении продуктов, синтезированных в ЭПР, участвуют в их химических перестройках, созревании; в цистернах аппарата Гольджи происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов. Кроме того, аппарат Гольджи является источником клеточных лизосом. Главной функцией аппарата Гольджи является процесс выведения готовых секретов за пределы клетки. Синтезированный на рибосомах экспортируемый белок накапливается внутри цистерн ЭПР, по которым он транспортируется к зоне мембран аппарата Гольджи. Здесь от гладких участков ЭПР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей комплекса Гольджи. В этом месте вакуоли сливаются друг с другом и с плоскими цистернами комплекса Гольджи. Далее белковый продукт переносится уже внутри полостей цистерн аппарата Гольджи. По мере движения происходит модификация белков. На противоположном полюсе диктиосомы происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Мелкие пузырьки сливаются друг с другом, образуя секреторный вакуоли. Такие вакуоли движутся к поверхности клетки, соприкасаются с плазматический мембраной и сливаются с ней (экзоцитоз). Таким образом, содержимое вакуолей оказывается за пределами клетки.
^ 2. Модификация белков в аппарате Гольджи.
Белки по мере движения по цистернам аппарата Гольджи «созревают», т.е. подвергаются модификациям: некоторые их аминокислоты фосфорилируются, ацетилируются и др. Подвергаются модификации и олигосахаридные цепи белков. При этом возникает специальный комплекс олигосахаридов. Такие превращения осуществляются с помощью ферментов – гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи. Так как каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека («этажа в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов.
В ряде специализированных клеток в аппарате Гольджи осуществляется синтез собственно полисахаридов. У растений это полисахариды матрикса клеточной стенки, у животных – синтез полисахаридных цепей глюкозааминогликанов.
- ^ Сортировка белков в аппарате Гольджи.
Через АГ проходит большой поток белков: поток гидролитических ферментов в компартмент лизосом; поток выделяемых белков, которые накапливаются в секреторных вакуолях и выделяются из клетки только при получении специальных сигналов; поток постоянно выделяемых секреторных белков. Разделение белковых потоков происходит в транс-участке АГ. Этот процесс доконца не расшифрован, но известно, что в сортировке принимают участие трансмембранные рецепторы.
1.5.Лизосомы.
Представляют собой пузырьки, ограниченные одиночной мембраной с разнородным содержимым внутри. Все лисосомы содержат ферменты гидролазы, что позволяет им участвовать в процессах внутриклеточного переваривания.
4.1.6. Пероксисомы (микротельца).
Строение. Это небольшие вакуоли (0,3 – 1,5 мкм), окруженные одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина. В зоне сердцевины располагаются кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Пероксисомы вероятно образуются из расширенных концов цистерн ЭПР.
Функции. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода (основной – каталаза). Так как перекись водорода является токсичным веществом, эти ферменты выполняют важную защитную роль.
4.1.7. Митохондрии.
Строение. Двумембранный органоид эукариотических клеток. Внешняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет ровные контуры, не образует впячиваний или складок. Ее толщина около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себе. Внешнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10 – 20 нм. Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрии, ее матрикс. Внутренняя мембрана образует впячивания внутрь митохондрии (кристы). Матрикс митохондрий содержит рибосомы, кольцевую молекулу ДНК, отложения солей магния и кальция.
Имея в своем составе молекулу ДНК, митохондрии обладают полной системой авторепродукции, т.е. на митохондриальной ДНК синтезируются информационные, транспортные и рибосомальные РНК и рибосомы, осуществляющие синтез митохондриальных белков. ДНК в митохондриях представлена циклическими молекулами, не образующие связь с гистонами, т.е. напоминают бактериальные нуклеоиды. У человека митохондриальная ДНК содержит 16,5 т.п.н., она полностью расшифрована. Все митохондриальные ДНК представлены множественными копиями, собранными в группы (1-50 циклических молекул на клетку). сиНтез митохондриальной ДНК не связан с синтезом ДНК в ядре. Митохондрии могут как делиться, так и сливаться друг с другом. Рибосомы митохондрий отличаются от рибосом цитоплазмы, они более мелкие (50S). ДНК митохондрий кодирует не все белки митохондрий, а только их часть (в основном структурные белки). Большинство митохондриальных белков синтезируется на рибосомах цитоплазмы. Совокупность митохондрий клетки называется хондриомом.
Функции Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
4.1.8. Пластиды
Строение. Двумембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). У высших растений найден целый ряд различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластид в другой.
Функции. Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты и синтезу сахаров и к выделению кислорода.. В других пластидах (лейкопласты, амилопласты) происходит отложение крахмала и каратиноидов (хромопласты).
4.1.9. Вакуоли растительных клеток.
Строение. Мембранная структура, представляющая собой мешок, образованный одинарной мембраной. Мембрана вакуоли носит название тонопласта. Полость вакуоли заполнена так называемым клеточным соком представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Функции. Одной из важнейших функций центральной вакуоли является поддержание тургорного давления. Вода поступает в концентрированный клеточный сок путем осмоса через избирательно проницаемый тонопласт. В результате в клетке развивается тургорное давление и цитоплазма прижимается к клеточной стенке. Осмотическое поглощение воды играет важную роль при растяжении клеток во время их роста.
В вакуолях растения могут накапливаться отходы жизнедеятельности и некоторые вторичные продукты его метаболизма (например, кристаллы оксалата кальция).
В вакуолях могут накапливаться пигменты антоцианы и родственные им соединения, которые придают цветкам и плодам красную, желтую, синюю или пурпурную окраску.
В вакуолях могут накапливаться запасные вещества, такие как сахара и белки.
В вакуолях растений иногда содержатся гидролитические ферменты. В этом случае, вакуоли действуют как лизосомы.
4.1.10. Клеточная стенка.
Строение. Встречается у прокариотических клеток и клеток растений. Это плотная многослойная структура, расположенная снаружи клеток. Клеточная оболочка является продуктом жизнедеятельности клетки. Клеточная стенка состоит из двух компонентов: аморфного пластичного гелеобразного матрикса с высоким содержанием воды и опорной фибриллярной системы. Часто для придания жесткости, несмачиваемости и др., в состав оболочек входят дополнительные полимерные вещества и соли. Главными компонентами клеточных стенок являются полисахариды. В состав матрикса оболочек входят полисахариды, растворяющиеся в концентрированных щелочах, гемицеллюлозы и пектиновые вещества. Волокнистые компоненты клеточных оболочек состоят обычно из целлюлозы. Во время деления клеток образуется первичная клеточная стенка. Позже в результате утолщения она может превратиться во вторичную клеточную стенку. Клеточные стенки соседних клеток скрепляет срединная пластинка. Некоторые клетки растений претерпевают интенсивную лигнификацию (одревеснение).
Функции Клеточные стенки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации.
Относительная жесткость клеточных стенок обуславливает тургорисцентность клеток, когда в них осмотическим путем поступает вода.
Клеточные стенки придают растительным клеткам определенную форму.
В клеточных стенках есть небольшие поры, сквозь которые проходят цитоплазматические тяжи – плазмодесмы. Плазмодесмы связывают содержимое отдельных клеток в единую систему – симпласт.
По клеточной стенке происходит передвижение воды и минеральных солей.
4.1.11. Рибосомы.
Строение Немембранный органоид клетки. Рибосомы – это сложные рибонуклеопротеидные частицы, в состав которых входят белки и молекулы рРНК примерно в равных соотношениях. Состоят из двух субединиц – большой и малой. Кроме ЭПР и гиалоплазмы обнаруживаются в митохондриях и пластидах (здесь они гораздо мельче). Эукариотические рибосомы имеют коэффициент седиментации 80 ед. Сведберга (80 S), рибосомы прокариот – 70S. Большая субъединица эукариотической рибосомы имеет коэффициент седиментации 60S, малая – 40S (у прокариот – 50S и 30S соответственно).
Функции Рибосомы являются местом синтеза белка в клетке.
4.1.11. Микротрубочки, микрофиламенты.
Строение. Микротрубочки располагаются в матриксе цитоплазмы. Это цилиндрические неразветвленные органеллы. Это очень тонкие трубочки диаметром приблизительно 24 нм. Их стенки толщиной около 5 нм. Построены из спирально упакованных глобулярных субъединиц белка тубулина. В длину они могут достигать нескольких микрометров. Растут микротрубочки с одного конца путем добавления тубулиновых субъединиц.
Функции. Микротрубочки входят в состав центриолей, базальных телец, ресничек, жгутиков. Микротрубочки участвуют также в перемещении других клеточных органелл, например пузырьков Гольджи. Кроме того, микротрубочки образуют опорную систему клетки – цитоскелет.
Микрофиламентами называются очень тонкие белковые нити диаметром 5 –7 нм. Эти нити состоят из белка актина и образуют цитоскелет, подобно микротрубочкам. Нередко микрофиламенты образуют сплетения или пучки непосредственно под плазматической мембраной. По-видимому, микрофиламенты участвуют также в экзо- и эндоцитозе. В клетке обнаруживаются и нити миозина (их количество значительно меньше). Взаимодействие актина и миозина лежит в основе сокращения мышц.
4.1.12. Клеточный центр.
Строение. Немембранный органоид клетки. Состоит из двух центриолей. Центриоли характерны и обязательны для клеток животных, их нет у высших растений, низших грибов и некоторых простейших. Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующие таким образом полый цилиндр. Обычно центриоли располагаются под прямым углом друг к другу.
Функции. В делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток эпителия и располагаются вблизи аппарата Гольджи.
4.2. Ядро.
Строение. Клеточное ядро, обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, кариоплазмы (или ядерного сока). Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариотических организмов. Ядро - самая крупная органелла, заключенная в оболочку из двух мембран. Оболочка ядра пронизана порами. Поры не являются просто сквозным отверстием, а представляют собой сложные комплексы мембран и белков. Внешняя ядерная мембрана непосредственно переходит в мембраны ЭПР, от нее могут отделяться вакуоли, переходящие в систему аппарата Гольджи.
Основным компонентом интерфазного ядра является хроматин, в состав которого входит ДНК в комплексе с белком. В делящихся клетках хроматиновые нити спирализуются и образуют хромосомы.
Внутри ядра практически всех эукаритических клеток видны одно или несколько округлой формы телец. Это ядрышки. Ядрышко – не самостоятельная структура или органоид. Оно – производное хромосомы, один из ее локусов, активно функционирующий в интерфазе. В состав ядрышка также входят рРНК и белки.
Все ядерные структуры погружены в ядерный сок (кариоплазма, нуклеоплазма).
Функции Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую - с ее реализацией, с обеспечением синтеза белка. В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК, что сохраняет строение молекул ДНК практически неизменными в ряду поколений. Кроме того, в ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые в качественном и количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но и транскрипция всех видов транспортных и рибосомных РНК. В ядре (ядрышке) также происходит образование субъединиц рибосом, путем сборки синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро. Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится.
4.3. Включения.
Кроме постоянных в клетке могут находиться временные компоненты цитоплазмы, продукты ее жизнедеятельности. Такие временные компоненты называются включениями.
ТАБЛИЦА 2. Строение клетки.
| Структура и схематическое изображение | Строение | Функции |
 | Тонкие пласты (6-10 нм), состоящие из бислоя липидов и молекул белка, расположенных на поверхности, погруженных или пронизывающих его. | Избирательно проницаемый барьер, регулирующий обмен между клеткой и средой. Плазмалемма отделяет цитоплазму от внешней среды, регулирует транспорт веществ в клетку и из нее, осуществляет межклеточные взаимодействия, принимат участие в делении клетки. |
| II. Органоиды | Специализированные структуры цитоплазмы, которые строением и выполняют жизнедеятельности постоянные компоненты обладают определенным определенные функции в клеток. | |
| II.1. Мембранные органоиды | | |
| II.1.1. Ядро 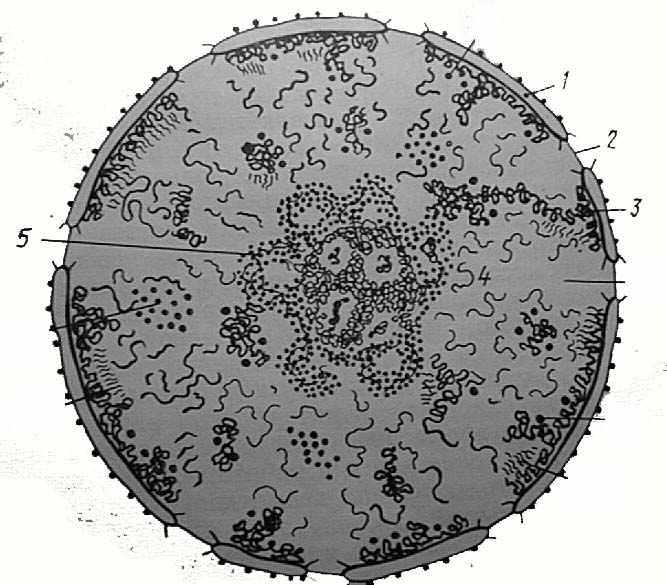 | Самая крупная органелла. Заключена в оболочку из двух мембран, пронизанную порами. Включает в себя хроматин (в интерфазе) или хромосомы (при делении), ядрышко и нуклеоплазму . | Хранение и реализация генетической информации. |
| II,1.2. Эндоплазматический ретикулум (ЭПР) | Система уплощенных мембранных мешочков – цистерн, узких каналов. Различают шероховатый (гранулярный) ЭПР, имеющий на мембранах рибосомы и гладкий (агранулярный) не имеющий их в своем составе. | Обеспечивает транспорт веществ как внутри клетки, так и между соседними клетками. Делит клетку на отдельные секции, компартменты, в которых одновременно происходят различные физиологические процессы и химические реакции.Функцией шероховатогоЭПР является синтез, сегрегация и созревание белков, гладкого – метаболизм липидов, стероидов и некоторых внутриклеточных полисахаридов. |
| II.1.3. Аппарат Гольджи | Стопка уплощенных мембранных мешочков –цистерн. На одном конце стопки мешочки непрерывно образуются, а с другого – отшнуровываются пузырьки. Стопки могут быть собраны в одной зоне (диктиосомы) или образуют пространственную сеть. | Накопление и модификация продуктов (г.о. белков), синтезированных в ЭПР. Транспорт этих веществ в пузырьках и выведение их за пределы клетки. Является источником лизосом. |
| II.1.4. Лизосомы | Простой сферический мембранный мешочек, заполненный пищеварительными (гидролитическими) ферментами | Расщепление, переваривание каких-либо клеточных структур или молекул; накопление непереваренных остатков; аутофагоцитоз (отбор и уничтожение измененных, дефектных клеточных элементов) |
| II.1.5. Пероксисомы (микротельца) | Небольшие вакуоли, окруженные одинарной мембраной. Содержимое имеет зернистую структуру, в центре иногда располагается кристаллоподобное образование. | Содержат ферменты метаболизма перекиси водорода (основной – каталаза). |
| II.1.6. Митохондрии | Окружены оболочкой из двух мембран. Внутренняя мембрана образует складки (кристы). Содержит матрикс, в котором находятся рибосомы, одна кольцевая молекула ДНК, включения. | Являются «энергетическими станциями» клетки – местом синтеза АТФ. При аэробном дыхании в кристах происходит окислительное фосфорилирование и перенос электронов, а в матриксе работают ферменты, участвующие в цикле Кребса и в окислении жирных кислот. |
| II.1.7. Пластиды (хлоропласты, хромопласты, лейкопласты, амилопласты) | Двумембранные органоиды, характерные для растительных клеток. Основная пластида – хлоропласт. Это крупная содержащая хлорофилл пластида. Обладает сложной системеой внутренних мембран. Выделяют ламеллы стромы и мембраны тилакоидов. Тилакоиды собраны в стопки - граны. Ламеллы свзывают граны друг с другом. В матриксе обнаруживаются рибосомы и кольцевая ДНК. | Функцией хлоропластов является фотосинтез. В других пластидах (лейкопласты, амилопласты) откладываются крахмал и каратиноиды (хромопласты). |
| II.1.8. Вакуоли | Крупные центральные вакуоли встречаются только у растительных клеток. Более мелкие встречаются как в клетках растений, так и в клетках животных. Представляет собой мешок, образованный одинарной мембраной, которая называется тонопластом. В вакуоли содержится клеточный сок – концентрированный раствор различных веществ, таких как минеральные соли, сахара, пигменты, органические кислоты и ферменты. | Здесь храняться различные вещества, в том числе и конечные продукты обмена. От содержимого вакуоли в сильной степени зависят осмотические свойства клетки. Иногда вакуоль выполняет функции лизосом. |
| II.1.9. Клеточная стенка | Плотная многослойная структура, расположенная снаружи клетки. Характерна для клеток растений и прокариот. Является продуктом жизнедеятельности клетки. Состоит из аморфного гелеобразного матрикса с большим содержанием воды, и опорной фибриллярной системы. Главным компонентом клеточной стенки являются полисахариды. В матриксе преобладают гемицеллюлозы и пектиновые вещества, волокнистым компонентом является целлюлоза. Соединяет клетки срединная пластинка, состоящая из пектиновых веществ. Может подвергаться вторичному утолщению и лигнификации. | Обеспечивает механическую опору и защиту, поддерживает форму клетки. Благодаря ей возникает тургорное давление, способствующее усилению опорной функции. По клеточной стенке происходит передвижение воды и минеральных солей. За счет пор и плазмодесм объединяет протопласты соседних клеток в единую непрерывную систему – симпласт (осуществляет транспорт веществ между этими клетками. |
| II.2. Немембранные органоиды | | |
| II.2.1. Рибосомы | Очень мелкие органеллы, состоящие из двух субъединиц – большой и малой. Содержат белок и РНК. Связаны с мембранами ЭПР или свободно лежат в цитоплазме. | Место синтеза белка в клетке. |
| II.2.2. Микротрубочки, микрофиламенты | Опорные структуры, располагаются в цитоплазме. Микротрубочки представляют собой цилиндрические неразветвленные трубочки, диаметром примерно 24 нм, длиной до нескольких мкм. Построены из спирально упакованных глобул белка тубулина. Микрофиламенты - тонкие белковые нити диаметром 5-7 нм, состям из белков актина или миозина. | Микротрубочки формируют цитоскелет, входят в состав центриолей, ресничек, жгутиков, участвуют в перемещении других клеточных органоидов. Микрофиламенты также образуют цитоскелет, участвуют в экзо- и эндоцитозе. Взаимодействие актина и миозина лежит в основе мышечного сокращения. |
| II.2.3. Клеточный центр | Состоит из двух центриолей. Характерны для животных клеток. Основой строения центриолей являются расположенные по окружности девять триплетов микротрубочек, образующих полый цилиндр. Расположены под прямым углом друг к другу. | Принимают участие в формировании веретена деления, располагаются на его полюсах. |
| III. Включения | Временные компоненты жизнедеятельности | цитоплазмы, продукты клетки |
| III.1. Трофические | Гликоген и жир в клетках животного организма | Запасные питательные вещества |
| III.2. Секреторные | Продукты жизнедеятельности железистых клеток (гормоны, ферменты,секреты) | Участвуют в работе внутренних органов (напрмер, ферменты кишечника, в регуляции обмена веществ) |
| III.3. Экскреторные | Конечные продукты обмена (соли, мочевая кислота). | Предназначены для выведения из организма. |
^ V. ЗНАЧЕНИЕ ЦИТОЛОГИИ ДЛЯ МЕДИЦИНЫ.
Знания структуры и функций клеток разных тканей, их субструктур, их взаимодействаия, протекающих в них процессов роста, развития, размножения и гибели, патологических нарушений их жизнедеятельности определяют клинические проявления болезней и имеют важнейшее значение для диагностики и лечения.
В настоящее время известно большое количество заболеваний, причиной которых являются дефекты клеточных органелл и связанное с этим изменение их функций.
Например, описано множество мутаций генов, кодирующих ферменты окислительного фосфорилирования митохондрий. Нарушение работы ферментов приводит к тяжелым порокам развития. Окислительное фосфорилирование служит основным источником энергии в клетках. Следовательно, дефекты на любом этапе этого пути могут приводить к нарушению работы мышц или головного мозга. Эти заболевания имеют множество фенотипических проявлений, в том числе миопатию, энцефалопатию и кардиомиопатию. Мутациями в митохондриальной ДНК также обусловлены наследственная атрофия зрительных нервов Лебера, сахарный диабет и глухота, доброкачественная опухоль онкоцитома и другие заболевания.
Некоторые наследственные заболевания связаны с нарушением функции пероксисом. Например, синдром Цельвегера обусловлен почти полной потерей пероксисомной функции. При этом синдроме в пероксисоме отсутствует большое число важных ферментов. Пациенты с данным синдромом умирают в детском возрасте. Даже нарушение функций одного пероксисомного фермента приводит к тяжелым заболеваниям (например адренолейкодистрофия).
Описано множество генетических заболеваний, связанных с лизосомными гидролазами. Часть из них, обобщенно называемая болезнями накопления мукополисахаридов, обусловлена недостаточностью особых лизосомных ферментов, разрушающих мукополисахариды. Неполностью разрушенные мукополисахариды накапливаются в тканях и окружающем межклеточном пространстве, выделяются с мочей. Эти заболевания вызывают прогрессирующую деградацию и приводят к смерти больного до 10-ти летнего возраста.
Известны заболевания, возникающие из-за изменений в стоении мебранных белков и липидов. Например, при муковисцедозе сложный мембранный белок, который находится в плазматической мембране и участвует в переносе хлоридов, испытывает изменение в одной аминокислоте. Однако это изменение оказывает сильное воздействие на локализацию белка в клетке, потому что изменившийся белок не может правильно складываться в эндоплазматической сети. Вместо своенго нормального перехода на клеточную мембрану, белок остается в эндоплазматической сети и распадается в цитоплазме. В результате нарушается транспорт хлоридов в эпителиальных клетках, они усиленно выводятся. Следствием этого является гиперсекреция густой слизи в клетках эндокринной части поджелудочной железы, эпителии бронхов, слизистой оболочки ЖКТ. Ферменты поджелудочной железы не поступают в просвет кишечника, закупориваются мелкие бронхи, что приводит к присоединению инфекции.
Знания в области молекулярной биологии клетки позволяют делать попытки лечения причины заболевания. Например, медикаментозная терапия онкологических больных заключается в разрушении митотического веретена веществами, нарушающими полимеризацию тубулина (например, колхицином). Применение этих препаратов обусловлено тем, что митотическое веретено, способствующее расхождению хромосом, состоит из микротрубочек. Антимитотические лекарственные средства ингибируют образование митотического веретена и уничтожает преимущественно быстро делящиеся раковые клетки. Посредством антигенов гистосовместимости и антигенов групп крови плазматическая мембрана обеспечивает генетическую индивидуальность клетки. Совместимость поверхностных клеточных антигенов во многом определяет успех переливания крови и трансплантации тканей.
За последние 20 лет знания, приобретенные в области молекулярной и клеточной биологии, полностью революционизировали представления о том, как функционируют клетки и предоставляют возможность манипулировать их функциями. Технологии рекомбинантных ДНК позволяет вводить естественные и модифицированные гены в клетки различных организмов, в том числе и человека, что имеет для врача глубокий смысл, потому что коренным образом изменяет нашу способность понимать, диагностировать и лечить болезни.