
Реферат по курсу «Анатомия центральной нервной системы» Тема: «Строение нейрона и нервного волокна»
| Вид материала | Реферат |
- Лекция 1 Введение. Общие сведения. Анатомия человека, 86.11kb.
- Учебно-методический комплекс дисциплины «анатомия центральной нервной системы» Специальности, 294.67kb.
- Учебно-методический комплекс по дисциплине «анатомия центральной нервной системы», 450.28kb.
- Как нервные клетки взаимодействуют друг с другом? Для согласованной работы нервной, 48.41kb.
- О. В. Анатомия центральной нервной системы: Программа, 315.76kb.
- «Анатомия и физиология центральной нервной системы и сенсорных систем», 256.06kb.
- Краткая учебно-методическая программа по специальности 030. 301. 65 Психология обоснование, 61.88kb.
- Программа дисциплины Анатомия и физиология центральной нервной системы для направления, 434.61kb.
- Практическая работа «Нейро-гуморальная регуляция функций организма человека», 83.83kb.
- Вопросы к экзамену по дисциплине «Функциональная анатомия центральной нервной системы», 52.3kb.
Университет Российской академии образования
Реферат по курсу
«Анатомия центральной нервной системы»
Тема: «Строение нейрона и нервного волокна»
1 курс
заочного обучения
факультета психологии
Москва
2009
План
- Нейрон ……………………...………………………………………………….3
- Структурная классификация нейронов ….…………………………………..4
- Функциональная классификация нейронов …………………………………5
- Развитие и рост нейрона ……………………………………………………...5
- Нервные волокна ……………………………………………………………...6
- Особенности строения и виды нервных волокон …………………………...6
- Механизм проведения возбуждения по нервному волокну ………………..7
- Классификация нервных волокон …………………………………………...8
- Литература …………………………………………………………………….9
Нейрон.
Нейрон (нервная клетка) – это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов.
Сложность и многообразие нервной системы зависит от взаимодействия между нейронами, которые, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
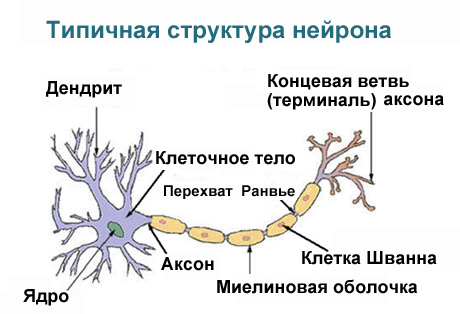
Нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро (с большим количеством ядерных пор) и другие органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), и отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
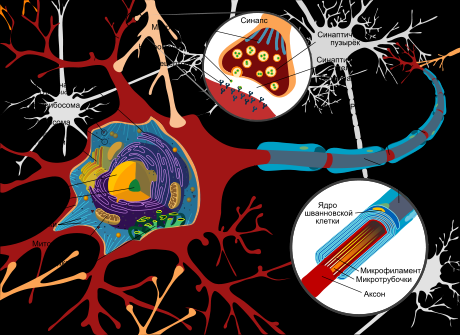
Схема строения нейрона
Аксон обычно — длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Си́напс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Структурная классификация.
На основании числа и расположения дейндритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны - небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны - нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны - нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах - сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
Мультиполярные нейроны - Нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе
Псевдоуниполярные нейроны - являются уникальными в своём роде. От тела отходит один остросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (т. е. находится вне тела клетки).
Функциональная классификация.
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны - ультиматные и предпоследние – неультиматные.
Ассоциативные нейроны (вставочные или интернейроны) - эта группа нейронов осуществляет связь между эфферентными и афферентными, их делят на комисуральные и проекционные (головной мозг).
Развитие и рост нейрона.
Нейрон развивается из небольшой клетки — предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным. Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Нервные волокна.
Нервные волокна имеют самую высокую возбудимость, самую высокую скорость проведения возбуждения, самый короткий рефрактерный период, высокую лабильность. Это обеспечивается высоким уровнем обменных процессов и низкой величиной мембранного потенциала.
Функция: проведение нервных импульсов от рецепторов к центральной нервной системе и обратно.
Особенности строения и виды нервных волокон.
Нервное волокно - аксон - покрыт клеточной мембраной.
Выделяют 2 вида нервных волокон.
Безмиелиновые нервные волокна - один слой швановских клеток, между ними - щелевидные пространства. Клеточная мембрана на всем протяжении контактирует с окружающей средой. При нанесении раздражения возбуждение возникает в месте действия раздражителя. Безмиелиновые нервные волокна обладают электрогенными свойствами (способностью генерировать нервные импульсы) на всем протяжении.
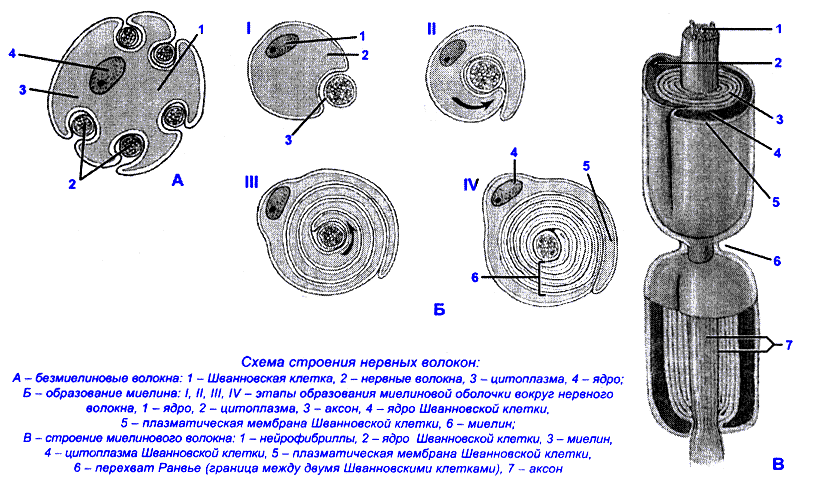
Миелиновые нервные волокна - покрыты слоями шванновских клеток, которые местами образуют перехваты Ранвье (участки без миелина) через каждые 1 мм. Продолжительность перехвата Ранвье 1 мкм. Миелиновая оболочка выполняет трофическую и изолирующую функции (высокое сопротивление). Участки, покрытые миелином не обладают электрогенными свойствами. Ими обладают перехваты Ранвье. Возбуждение возникает в ближайшем к месту действия раздражителя перехвата Ранвье. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом перехвате Ранвье происходит усиление нервных импульсов.
Перехваты Ранвье выполняют функцию ретрансляторов (генерируют и усиливают нервные импульсы).
Механизм проведения возбуждения по нервному волокну.
1885 г. - Л. Герман - между возбужденными и невозбужденными участками нервного волокна возникают круговые токи.
При действии раздражителя имеется разность потенциалов между наружной и внутренней поверхностями ткани (участки несущие различные заряды). Между этими участками возникает электрический ток (движение ионов Nа+). Внутри нервного волокна возникает ток от положительного полюса к отрицательному полюсу, т. е. ток направлен от возбужденного участка к невозбужденному. Этот ток выходит через невозбужденный участок и вызывает его перезарядку. На наружной поверхности нервного волокна ток идет от невозбужденного участка к возбужденному. Этот ток не изменяет состояние возбужденного участка, т. к. он находится в состоянии рефрактерности.
Доказательство наличия круговых токов: нервное волокно помещают в раствор NaCl и регистрируют скорость проведения возбуждения. Затем нервное волокно помещают в масло (повышается сопротивление) - скорость проведения уменьшается на 30 %. После этого нервное волокно оставляют на воздухе - скорость проведения возбуждения уменьшается на 50 %.
Особенности проведения возбуждения по миелиновым и безмиелиновым нервным волокнам:
1) миелиновые волокна - имеют оболочку обладающую высоким сопротивлением, электрогенные свойства только в перехватах Ранвье. Под действием раздражителя возбуждение возникает в ближайшем перехвате Ранвье. Соседний перехват в состоянии поляризации. Возникающий ток вызывает деполяризацию соседнего перехвата. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом следующем перехвате возникает чуть больший (по амплитуде) потенциал действия, за счет этого возбуждение распространяется без декремента и может перескакивать через несколько перехватов. Это сальтаторная теория Тасаки. Доказательство теории - в нервное волокно вводили препараты, блокирующие несколько перехватов, но проведение возбуждения регистрировалось и после этого. Это высоко надежный и выгодный способ, т. к. устраняются небольшие повреждения, увеличивается скорость проведения возбуждения, уменьшаются энергетические затраты;
2) безмиелиновые волокна - поверхность обладает электрогенными свойствами на всем протяжении. Поэтому малые круговые токи возникают на расстоянии в несколько микрометров. Возбуждение имеет вид постоянно бегущей волны. Этот способ менее выгоден: большие затраты энергии (на работу Nа-К-насоса), меньшая скорость проведения возбуждения.
Классификация нервных волокон
Нервные волокна классифицируются по:
- длительности потенциала действия;
- строению (диаметру) волокна;
- скорости проведения возбуждения.
Выделяют следующие группы нервных волокон:
- группа А (альфа, бета, гамма, дельта) - самый короткий потенциал действия, самая толстая миелиновая оболочка, самая высокая скорость проведения возбуждения;
- группа В - миелиновая оболочка менее выражена;
- группа С - без миелиновой оболочки.
Литература
- Физиология человека: Учебник / В двух томах. Т. I / В.М. Покровский, Г.Ф. Коротько, В.И. Кобрин и др.; Под ред. В.М. Покровского, Г.Ф. Коротько. — М.: Медицина, 1997. — 448 с.
- В.В. Жуков, Е.В. Пономарева. Анатомия нервной системы: Учебное пособие / Калинингр. ун-т. - Калининград, 1998. - 68 с.