
Российская академия медицинских наук нии нормальной физиологии им. П. К. Анохина
| Вид материала | Автореферат диссертации |
СодержаниеРезультаты и обсуждение |
- Психосоциальная проблема, направленная на восстановление физического и психосоциального, 348.21kb.
- Донозологический контроль соматического здоровья населения 03. 00. 13- физиология, 836.84kb.
- Сергиев Владимир Петрович Член-корреспондент рамн, доктор медицинских наук, профессор, 829.91kb.
- Спонсоры конференции: Фармацевтическая фирма «Санофи-Авентис», 74.5kb.
- Acta Neurobiologia Experimentalis», «Biological Theory», а также членом диссертационных, 19.24kb.
- Составители: В. В. Зинчук, 610.47kb.
- Основание Петербургской академии наук, 49.85kb.
- Котов Сергей Викторович доктор медицинских наук, профессор Савин Алексей Алексеевич, 547.92kb.
- Академия медицинских наук СССР научный совет по физиологии человека, 4592.08kb.
- Цель настоящей программы состоит в изучении строения здорового организма и жизнедеятельности, 472.25kb.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Влияние тетродотоксина на динамику АСР и поведение замирания при введении в червь мозжечка животных с различным уровнем оборонительной мотивации. Сравнение с эффектами введения ТТХ в гиппокамп.
Для выбора адекватной модели обучения оценивали динамику АСР и поведения замирания у животных, которых при обучении помещали в экспериментальную обстановку впервые (неадаптированные крысы), а также у крыс, которых за 24 часа до обучения адаптировали к экспериментальной камере в течение 5 минут (адаптированные крысы).
Согласно литературным данным, брадикардическая реакция отражает ориентировочно-исследовательскую составляющую АСР, а тахикардия является показателем оборонительной компоненты реакции (Taylor, Printz, 1996, Gautier, Cook, 1997, Richardson et al., 1996, Lang & Davis, 2006). Поэтому у животных указанных групп проводили измерение частоты сердечных сокращений (ЧСС) с целью определения соотношения ориентировочно-исследовательской и оборонительной составляющих поведения замирания и АСР.
Обнаружено, что в день обучения ЧСС на фоне замирания перед началом звуковой стимуляции у адаптированных крыс была на 12 % ниже, чем у неадаптированных и на 8% меньше соответствующего показателя во время сеанса адаптации (p<0,05). Таким образом, предварительная адаптация животного к экспериментальному контексту снижает уровень оборонительной мотивации, вызванной изменением обстановочной афферентации.
Таблица 1. Основные показатели динамики АСР и поведения замирания у адаптированных и неадаптированных к обстановке крыс.
| Группы животных | Амплитуда первой АСР | Индекс восстановления АСР (Iсв) | Коэффициент регрессии b1 исходно | Коэффициент регрессии b2 через 24 часа | Время замирания исходно (с) | Время замирания через 24 часа |
| Неадаптиро-ванные (суммарно) n=36 | 17,9±2,1 | -3,54±2,8 | 0,70±0,13 | 0,56±0,15 | 61,6±7,8 | 118,6±7,8* |
| Неадаптиро-ванные (b>0) n=13 | 18,2 ±3,4 | -7,9 ±2,9 | 1,15 ±0,30 | 0,63 ±0,19 | 66,0± 9,6 | 108,2± 9,6* |
| Неадаптиро- ванные (b=0) n=23 | 17,4 ±3,0 | 2,5± 2,64 | 0,37± 0,19 | 0,49± 0,19 | 57,6± 9,4 | 129,6±12,2 |
| Адаптиро-ванные (суммарно) n=35 | 13,1±1,2# | 0,028±0,10 | 0,19±0,12# | 0,20±0,08# | 53,8±10,4 | 96,8±8,2#* |
| Адаптиро-ванные (b>0) n=11 | 11,9±1,8# | -2,82±1,9 | 0,79±0,14 | 0,69±0,16 | 52,8±7,4 | 130,2±9,8* |
| Адаптиро-ванные b=0 n=11 | 12,1±2,7# | 0,83±0,49 | -0,05±0,21 | 0,06±0,19 | 31,8±10,4 | 73,2±13,6#* |
| Адаптиро-ванные (b1<0) n=13 | 14,0±2,4 | 1,60±0,9 | -0,51±0,18 | -0,06±0,09 | 42,6±7,8 | 70,2±10,8* |
* - p<0.05 по сравнению с исходным уровнем, # p<0.05 по сравнению с неадаптированными крысами.
Оценка средне-групповых значений коэффициентов регрессии b1 и b2 показала (табл.1), что неадаптированные крысы в сравнении с адаптированными демонстрируют более выраженное снижение величины АСР во время обучения и при тестировании. Дальнейший анализ обнаруженных различий с оценкой индивидуальных величин коэффициента b1 позволил выявить в группе адаптированных животных три подгруппы, представленные практически в равной пропорции: 1 – крысы с доминированием процесса угашения (b1>0), 2 - крысы без выраженной тенденции в динамике АСР (b1 не демонстрирует достоверных отличий от нуля, поэтому принимаем b1=0), 3 – крысы с доминированием сенситизации (b1<0). В группе неадаптированных животных подгруппа с доминированием сенситизации отсутствовала, при этом около 2/3 от общего числа (23 из 36) составляли животные с b1=0 (табл.1.). Таким образом, предварительная адаптация к обстановке приводит к возникновению у части животных кратковременной сенситизации АСР.
А Б

В Г
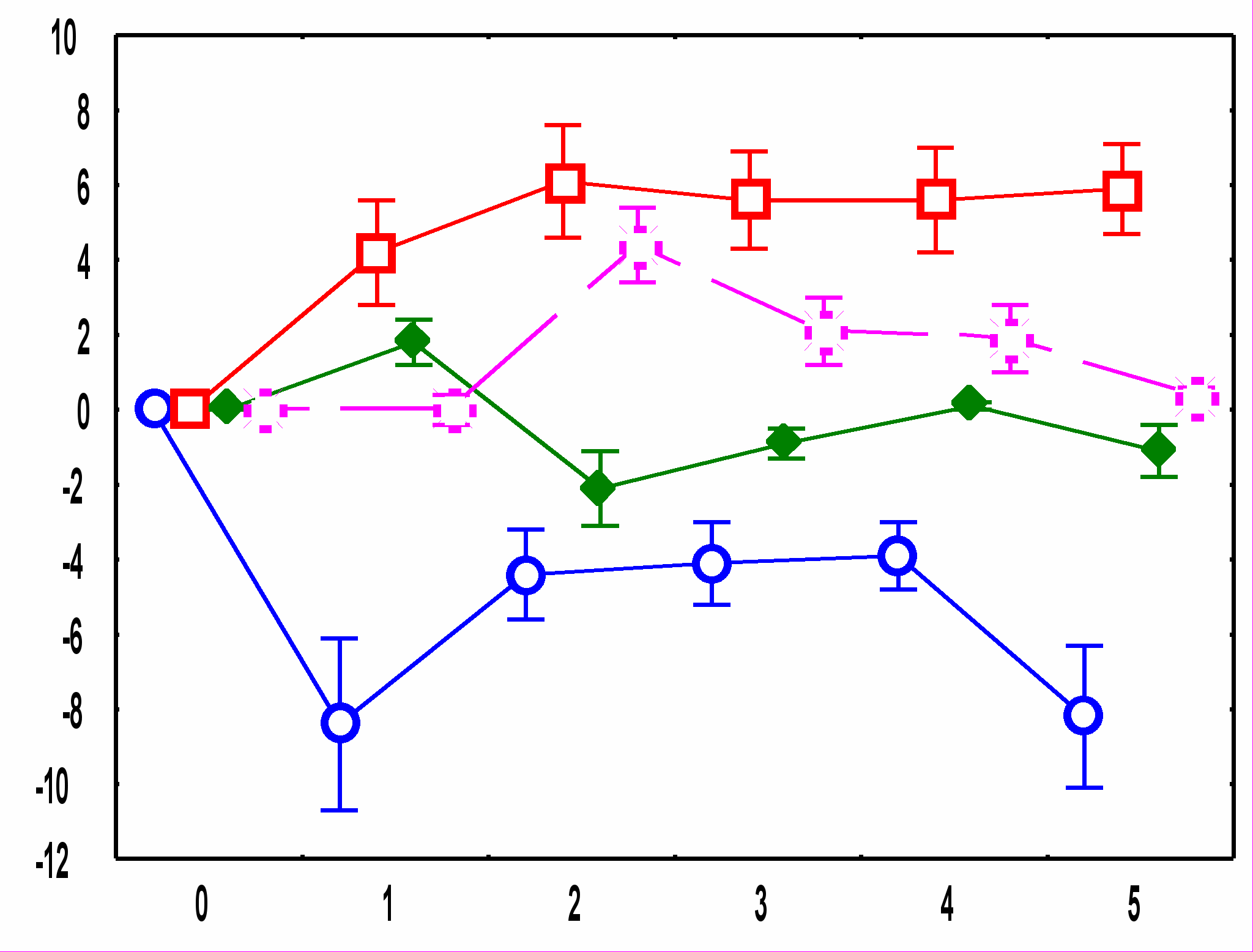

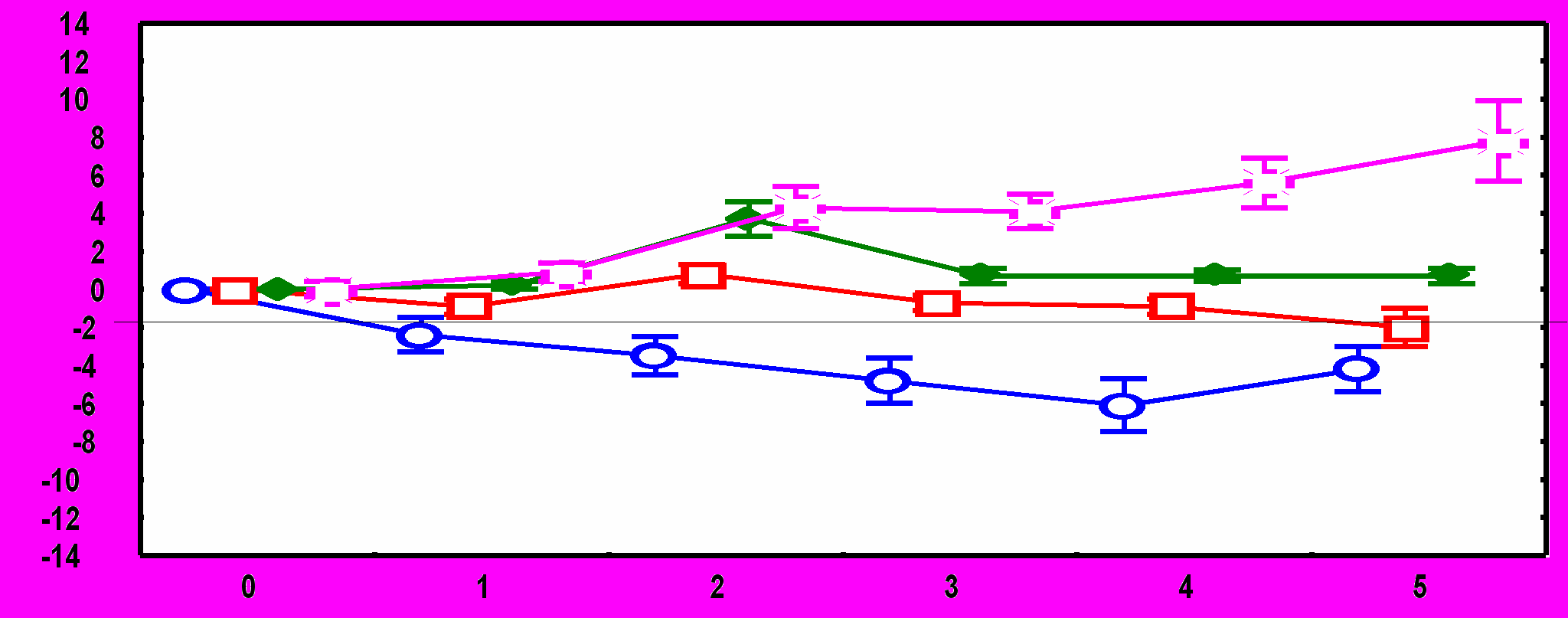
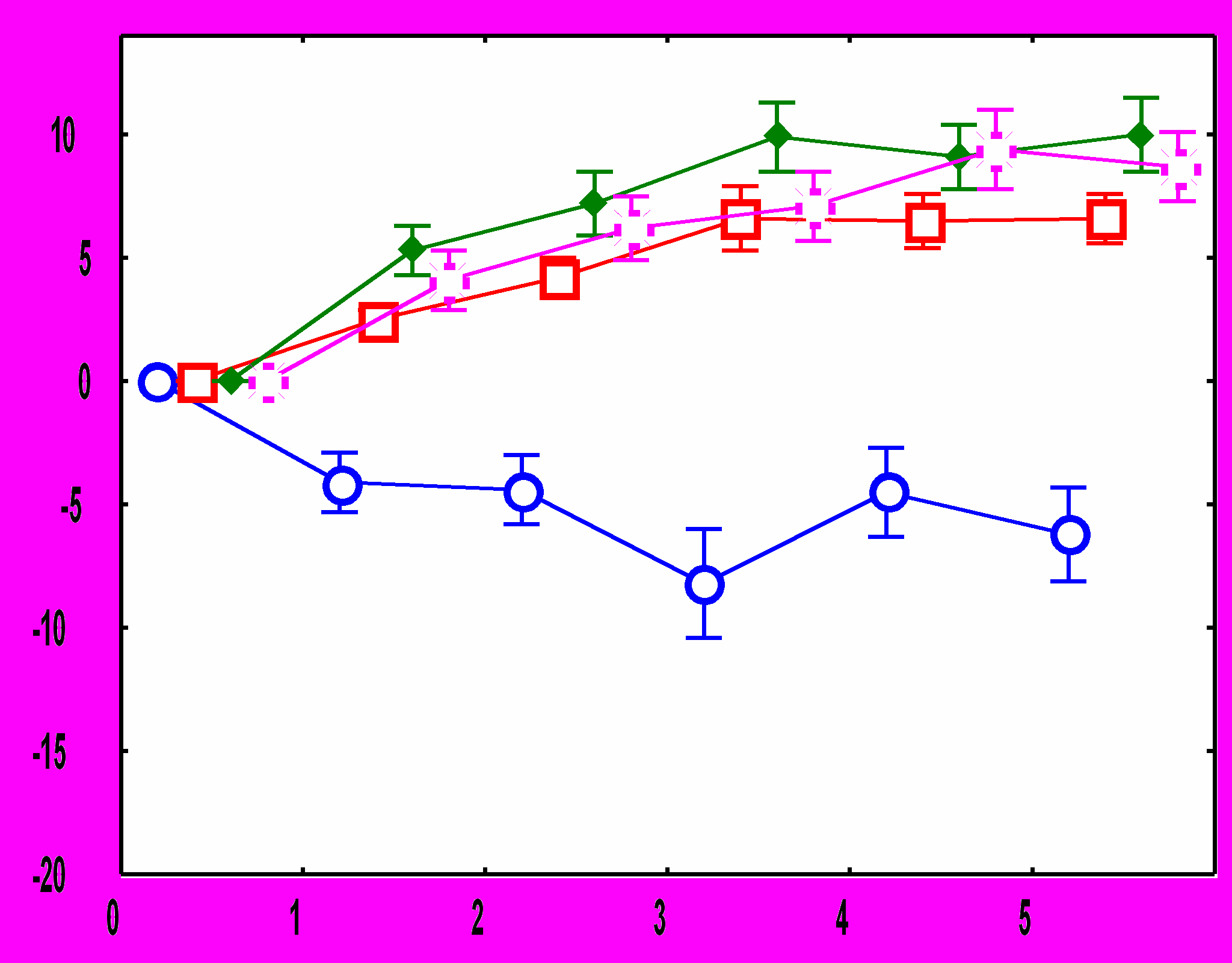
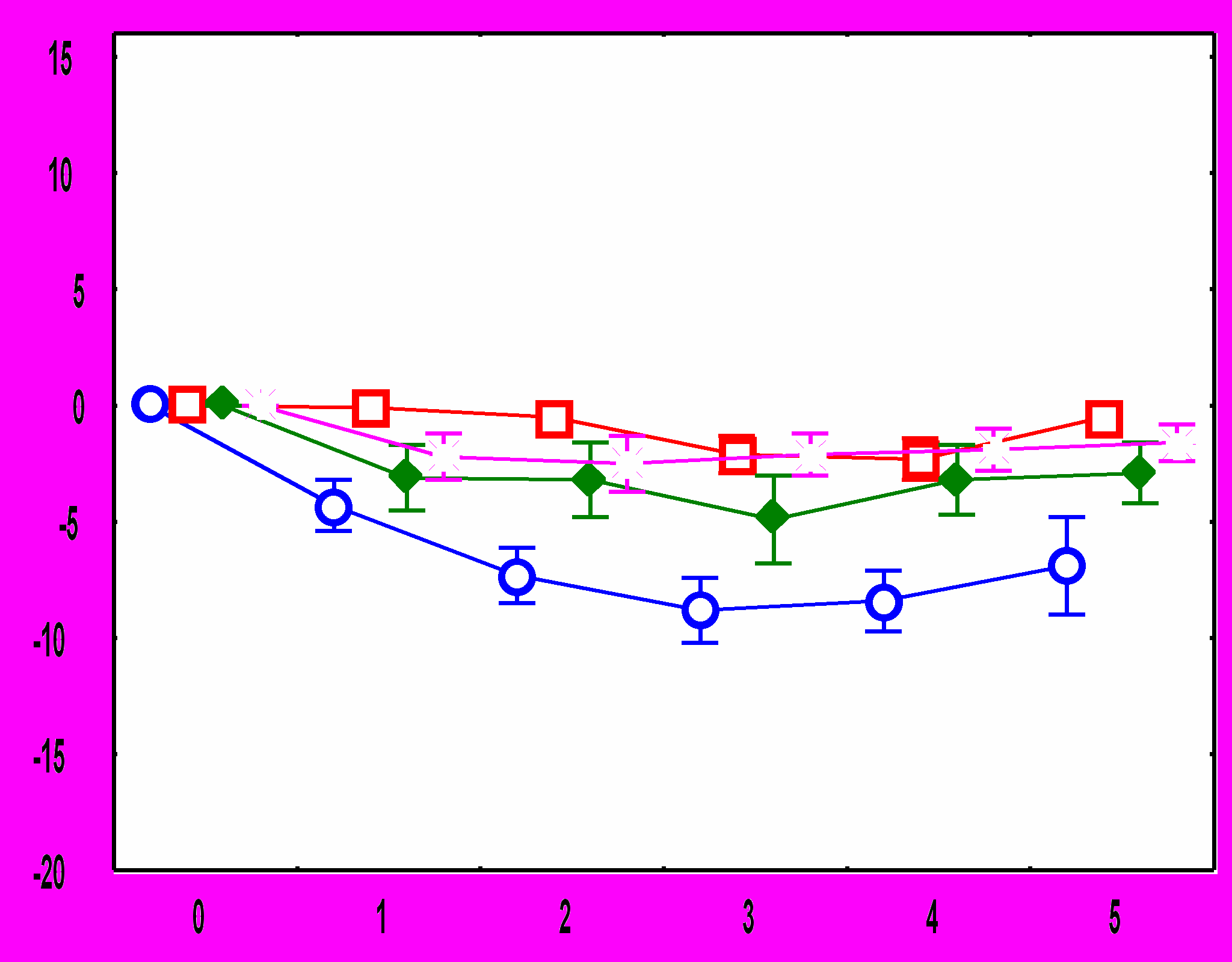
Рис.1. Изменения ЧСС после подачи звукового стимула: А - во время обучения у адаптированных крыс, Б – во время обучения у неадаптированных крыс, В – во время тестирования у адаптированных крыс, Г – во время тестирования у неадаптированных крыс. По оси абсцисс – время от начала стимула (с), По оси ординат – изменения ЧСС (1/с).
 1-й стимул,
1-й стимул,  5-й стимул,
5-й стимул,  7-й стимул,
7-й стимул,  10-й стимул
10-й стимулСущественные межгрупповые отличия обнаружены и в динамике реакций ЧСС на звуковые стимулы. У адаптированных крыс в день обучения в ответ на первый звуковой сигнал наблюдается снижение ЧСС.
Ко второму стимулу брадикардическая реакция угасает, а с 4 по 10 стимул имеет место возрастание ЧСС. В день тестирования снижение ЧСС происходит в ответ на 1-й стимул, на 4-6-й - наблюдается тахикардический ответ; который угасает при дальнейшей стимуляции (рис.1). У неадаптированных животных в день обучения снижение ЧСС наблюдается в ответ на 1-й и 2-й стимулы, затем брадикардическая реакция угасает, но в отличие от адаптированной группы, не сменяется тахикардической: уровень ЧСС после предъявления 3-10 звуковых стимулов не отличается от фоновых величин. В день тестирования у животных этой группы брадикардический ответ наблюдается только на 1-й стимул, а к 7-му стимулу развивается тахикардическая реакция (рис 1).
Таким образом, предварительная адаптация к экспериментальной обстановке повышает выраженность оборонительной составляющей в структуре АСР и вызывает у части животных (около 30%) кратковременную сенситизацию реакции.
Аппликация ТТХ на червь мозжечка неадаптированных крыс за 1 час до обучения вызывала достоверное возрастание времени замирания при первом помещении в экспериментальную камеру (109,0±15,8 с по сравнению с 66,6±6,8 с в контроле, р<0,05). Повышения данного показателя в день тестирования по сравнению с днем обучения в экспериментальной группе не наблюдалось (115,6 ±11,2 с) в то время как в контрольной группе его значение возрастало почти вдвое (до 114,2±8,6). Влияния ТТХ на исходный уровень и динамику АСР в этих условиях обнаружено не было.
Аппликация ТТХ на червь мозжечка адаптированных крыс не влияла на исходный уровень АСР и замирания (табл.2), однако приводила к достоверному уменьшению числа животных, демонстрирующих кратковременную сенситизацию АСР и возрастанию численности подгруппы животных без выраженной динамики реакции. При этом в подгруппе с b1=0 индекс спонтанного восстановления реакции был достоверно отрицательным, т.е. имела место долговременная сенситизация АСР. Во всех подгруппах экспериментальных крыс наблюдалось также нарушение формирования условного обстановочного замирания. При оценке долговременного привыкания АСР по величине коэффициента b2 достоверных различий между группами не обнаружено.
Таблица 2. Влияние тетродотоксина на угашение АСР и поведение замирания при аппликации на червь мозжечка адаптированных к обстановке крыс.
| Группы животных | Амплитуда первой АСР | Индекс восстановления АСР | Коэффициент регрессии b1 исходно | Коэффициент регрессии b2 через 24 часа | Время замирания исходно | Время замирания через 24 часа |
| Контроль (суммарно) n=32 | 12,3±1,1 | 0,031±0,9 | 0,17±0,13 | 0,29±0,09 | 54,4±5,3 | 88,6±11,8* |
| Контроль (b1>0) n=10 | 11,5±1,7 | -2,51±1,8 | 0,62±0,14 | 0,58±0,14 | 50,6±6,1 | 70,4±6,3* |
| Контроль (b1<0) n=12 | 13,5±2,1 | 1,51±0,9 | -0,57±0,12 | -0,04±0,08 | 37,0±8,6 | 65,4±12,2* |
| Контроль (b1=0) n=10 | 11,8±2,4 | 0,96±0,51 | -0,04±0,13 | 0,21±0,14 | 20,6±4,9 | 64,4±16,2* |
| ТТХ (суммарно) n=32 | 12,8± 1,6 | -2,9± 1,1 | 0,38± 0,16# | 0,45± 0,12 | 58,8±10,0 | 77,2± 9,4 |
| ТТХ (b1>0) n=14 | 14,1±2,6 | -2,52±1,9 | 0,86±0,28 | 0,41±0,18 | 51,6±16,8 | 75,6±14,2 |
| ТТХ (b1<0) n=3 | 12,5±1,9 | 1,51±0,8 | -0,93±0,31 | 0,34±0,44 | 53,0±14,6 | 55,0±12,4 |
| ТТХ (b1=0) n=15 | 13,3±2,3 | -4,2±1,6# | -0,05±0,24 | 0,52±0,24 | 53,4±13,8 | 79,2±18,8 |
У адаптированных крыс с введением ТТХ в червь мозжечка обнаружена отрицательная корреляционная зависимость между временем замирания в день обучения до начала звуковой стимуляции и амплитудой первой АСР (r=–0,59, р=0,0017 по сравнению с r=–0,12 р>0,1 в контроле) и положительная корреляционная зависимость (r=0,36, p<0,05) между индексом восстановления реакции и изменением времени замирания в день тестирования по сравнению с днем обучения, (в контрольной группе r=0,19, p>0,1).
Таблица 3. Влияние ТТХ на динамику АСР и поведение замирания при введении в гиппокамп неадаптированных к экспериментальной обстановке крыс.
| Группы животных | Амплитуда первой АСР | Индекс восстановле-ния АСР | Коэффициент регрессии b1 исходно | Коэффициент регрессии b2 через 24 часа | Время замирания исходно | Время замирания через 24 часа |
| Контроль (суммарно) n=24 | 19,5±2,3 | -5,4±2,1 | 0,67±0,16 | 0,70±0,13 | 66,8±6,8 | 114,2±8,6* |
| Контроль (b>0) n=8 | 23,7 ±2,7 | -8,2 ±3,8 | 1,11 ±0,10 | 0,94 ±0,37 | 74,2± 13,2 | 116,0± 16,2* |
| Контроль (b=0) n=16 | 17,4±2,3 | -3,6± 2,6 | 0,07± 0,10 | 0,43± 0,28 | 58,8±11,6 | 105,2 ±18,2* |
| ТТХ (суммарно) n=18 | 27,1± 2,5# | -13,2± 4,7# | 1,51± 0,34 | 0,15± 0,33# | 53,4 ±11,2 | 67,4± 16,2 |
| ТТХ (b>0 n=16 | 26,0± 2,62 | -16,5± 2,7# | 1,74± 0,13# | 0,06± 0,31# | 59,8± 22,8 | 70,4± 15,7 |
| ТТХ b=0 n=4 | 31,0± 4,89# | - 8,0±5,9 | 0,59±0,41 | 0,54±0,39 | 51,6± 7,8 | 48,0± 8,2 |
* - p<0.05 по сравнению с исходным уровнем, # p<0.05 по сравнению с контролем
У неадаптированных крыс введение ТТХ в гиппокамп приводило к возрастанию исходного уровня АСР, стимуляции кратковременного угашения и возникновению долговременной сенситизации реакции, (оцениваемой по показателю спонтанного восстановления и величине коэффициента b2), а также нарушало формирование условного замирания (табл.3).
При введении ТТХ в гиппокамп адаптированных крыс (табл.4) достоверно возрастала численность подгруппы без выраженной динамики АСР (р=0,037) за счет уменьшения количества крыс с кратковременным угашением реакции. В экспериментальной группе этой серии наблюдалось повышение исходного времени замирания в день обучения (табл.4), однако не было обнаружено дальнейшего возрастания времени замирания в день тестирования.
Таким образом, независимо от адаптации к обстановке, введение ТТХ как в гиппокамп, так и в мозжечок, нарушает формирование условной реакции замирания. Эффекты «инактивации» мозжечка и гиппокампа на другие показатели оборонительного поведения зависят от адаптации к экспериментальной обстановке и, в ряде случаев, имеют противоположную направленность: изменения, наблюдаемые при депрессии активности мозжечка у адаптированных животных,
сходны с эффектами, наблюдаемыми при депрессии гиппокампа у неадаптированных крыс, и наоборот (табл.5).
Таблица 4. Влияние ТТХ на динамику АСР и поведение замирания при введении в гиппокамп адаптированных к экспериментальной обстановке крыс
| Группы животных | Амплитуда первой АСР | Индекс восстановле-ния АСР | Коэффициент регрессии b1 исходно | Коэффициент регрессии b2 через 24 часа | Время замирания исходно | Время замирания через 24 часа |
| Контроль (суммарно) n=24 | 13,5±3,5 | 0,21±0,6 | 0,25±0,14 | 0,24±0,11 | 53,2±8,7 | 99,2±12,8* |
| Контроль (b>0) n=9 | 14,9±3,7 | 1,58±1,4 | 0,71±0,16 | 0,61±0,23 | 98,5±10,7 | 136,3±18,1* |
| Контроль (b=0) n=8 | 12,8±2,9 | 0,83±0,42 | -0,11±0,19 | -0,09±0,10 | 25,9±6,2 | 59,2±15,8* |
| Контроль (b<0) n=7 | 13,7±4,2 | - 1,47±0,5 | -0,43±0,14 | -0,10±0,08 | 38,2±7,2 | 70,9±8,4* |
| ТТХ (суммарно) n=24 | 10,1 ± 3,4 | 0,23± 0,7 | 0,18± 0,19 | 0,31± 0,09 | 112,6±22,8 | 88,0 ±18,4 |
| ТТХ (b>0) n=3 | 11,8± 4,9 | -1,64±1,1 | 0,94± 0,37 | 0,52± 0,18 | 40± 9,9 | 58,8± 12,2 |
| ТТХ b=0 n=15 | 9,9 ± 3,0 | 0,48± 0,29 | 0,07± 0,08 | 0,08± 0,09 | 158,2 ±8,4 | 88,0 ±22,4 |
| ТТХ (b<0) n=7 | 10,6± 2,7 | 1,02±0,85 | -0,81± 0,39 | -0,32± 0,19 | 82,0 ±26,2 | 107,5 ±39,9 |
*- p<0.05 по сравнению с исходным уровнем,
Учитывая данные о соотношении ориентировочно-исследовательской и оборонительной составляющих поведения замирания и АСР у адаптированных и
Таблица 5. Эффекты нарушений функций мозжечка и гиппокампа на поведение замирания и угашение АСР у адаптированных и неадаптированных животных.
| Структура, в которую вводили ТТХ/опыт адаптации к контексту | Кратковре-менное угашение АСР | Спонтанное Восстановле- ние АСР | Замирание исходно | Возрастание замирания в день тестирования |
| Мозжечок, неадаптированные | 0 | 0 | + | - |
| Гиппокамп, недаптированные | + | + | 0 | - |
| Мозжечок, адаптированные | + | + | 0 | - |
| Гиппокамп, адаптированные | - | 0 | + | - |
неадаптированных крыс, можно сделать вывод, что подавление активности мозжечка оказывает влияние на эти формы поведения в условиях более высокого уровня оборонительной составляющей, а при доминировании ориентировочной компоненты более выраженные эффекты наблюдаются при подавлении активности гиппокампа..
Исходя из полученных результатов, дальнейшие эксперименты проводили на адаптированных к обстановке животных