
Биологическая фиксация азота в однолетних агроценозах лесостепной зоны Западной Сибири
| Вид материала | Автореферат |
СодержаниеСимбиотическая фиксация атмосферного азота зернобобовыми культурами Rhizobium leguminosarum |
- Эволюция культуры лесостепной части западной сибири второй четверти II − середины, 312.04kb.
- Мо Нитрогеназа Azotobacter Фиксация азота Mo,, 74.74kb.
- Перспективы развития рынка кредитования физических лиц Западной Сибири, 75.44kb.
- Программа дисциплины археология западной сибири дпп. Дс. 02 Цели и задачи дисциплины, 431.44kb.
- Программа дисциплины источниковедение и историография западной сибири для аспирантов, 223.01kb.
- История Западной Сибири) реферат (от лат еfеrо «сообщаю») краткое изложение, 69.17kb.
- Ареология флоры Западной Сибири, 22.43kb.
- Ареология флоры Западной Сибири”, 34.99kb.
- Урок 46. Природные ресурсы Западной Сибири. Проблемы их освоения тип урока, 57.84kb.
- Тюменский государственный нефтегазовый университет Гуманитарный институт Лаборатория, 27.71kb.
Анализ экспериментальных данных, полученных в опытах с мягкой пшеницей, свидетельствует о том, что в годы достаточной влагообеспеченности эффективность, как минеральных удобрений, так и бактериальных препаратов, особенно азоризина, довольно высока. Так, внесение N60P60K40 обеспечило формирование урожайности яровой пшеницы на уровне 29 ц/га, что на 43-52 % выше, чем на не удобренных посевах. Эффект инокуляции семян пшеницы азоризином в этих условиях составил 4,2-5,1 ц/га или 17-22 % к фоновому варианту.
Отмеченные закономерности формирования урожая пшеницы на различных фонах питания отражаются на выносе азота надземной биомассой. Так, применение минеральных удобрений повышает содержание азота в зерне пшеницы в среднем на 0,15 % при дозе N30 и на 0,36 %, при дозе N60. Существенное изменение этого показателя в соломе (+0,2%) наблюдается лишь при внесении 60 кг/га азота (табл. 4).
Таблица 4 – Содержание и вынос азота урожаем яровой пшеницы в зависимости от фона питания (в среднем за 2001-2003 гг.)
| Вариант | Содержание азота, % | Выносазота, кг/га | Дополнительный вынос (кг/га) в сравнении с: | ||||
| зерно | солома | зерно | солома | всего | контролем | фоном | |
| Без удобрений | 2,14 | 0,42 | 37,1 | 14,2 | 51,3 | – | – |
| N30P60K40 (фон) | 2,29 | 0,42 | 47,7 | 17,0 | 64,7 | 13,4 | – |
| Фон+азоризин | 2,33 | 0,50 | 56,9 | 23,1 | 80,0 | 28,7 | 15,3 |
| Фон+ризоагрин | 2,30 | 0,52 | 49,4 | 22,2 | 71,6 | 20,3 | 6,9 |
| Фон+Д-65 | 2,23 | 0,46 | 47,0 | 19,2 | 66,2 | 14,9 | 1,5 |
| N60P60K40 | 2,50 | 0,63 | 62,6 | 31,9 | 94,5 | 43,2 | 29,8 |
| НСР05 | 0,11 | 0,06 | 7,6 | 6,7 | 13,0 | | |
Рост урожайности и повышение концентрации азота в биомассе растений приводят к увеличению выноса элемента урожаем на 26-84%. При этом установлено, что из изучаемых биопрепаратов максимальный вынос азота обеспечивает азоризин (+15,3 кг/га к фону). Этот эффект примерно в 2 раза ниже, чем действие минерального азота (N60 в сравнение с N30), но следует учитывать, что если минеральные удобрения увеличивают вынос азота зерном и соломой в равной степени, то на фоне биопрепарата отмечается более высокий вынос элемента зерном. Это приводит к тому, что доля азота в зерне на варианте N60P60K40 составляет 66 % от общего выноса, а при замене части минерального азота на биологический (N30P60K40 + азоризин) этот показатель возрастает до 71 %.
Дополнительно усвоенный азот при инокуляции используется растениями, главным образом, на формирование прибавки урожая. Этот вывод подтверждается и данными по содержанию белка в зерне пшеницы (табл. 5). Если под воздействием минеральных удобрений этот показатель вырастает в среднем на 0,9 % (N30P60K40) или 2,1 % (N60P60K40), то существенных его изменений на вариантах инокуляции не зафиксировано.
Таблица 5 – Влияние минеральных и бактериальных удобрений на содержание белка в зерне пшеницы, %
| Вариант | 2001 г. | 2002 г. | 2003 г. | В среднем за 2001-2003 гг. |
| Без удобрений | 13,9 | 12,3 | 11,6 | 12,6 |
| N30P60K40 (фон) | 14,6 | 13,1 | 12,7 | 13,5 |
| Фон + азоризин | 15,3 | 13,1 | 12,5 | 13,6 |
| Фон + ризоагрин | 15,5 | 12,9 | 12,1 | 13,5 |
| Фон + Д-65 | 14,6 | 13,0 | 11,7 | 13,1 |
| N60P60K40 | 15,7 | 14,4 | 13,9 | 14,7 |
| НСР05 | 0,88 | 0,59 | 0,65 | 0,64 |
Таким образом, по результатам проведенных исследований можно заключить, что применение бактериальных удобрений на основе корневых диазотрофов позволяет в 1,5-2,0 раза увеличить азотфиксирующую активность в ризосфере зерновых культур и повысить продуктивность агроценозов на 15-40 %.
При этом отмечается увеличение содержания белка в зерне на 0,4-1,0 %, в зависимости от препарата, выращиваемой культуры и условий увлажнения.
Различная ответная реакция культур на тот или иной биопрепарат, что связана с биологическими особенностями культур, с адаптацией применяемых штаммов микроорганизмов к сообществу микрофлоры почвы и с их устойчивостью к изменениям гидротермического режима в период вегетации растений. Это вызывает необходимость подбора наиболее эффективного в конкретных условиях ассоциативного комплекса: штамм микроорганизма – культура.
Отличительной особенностью действия биопрепаратов является преимущественное поступление дополнительно усвоенного азота в репродуктивные органы растений и пропорциональное увеличение их массы, в результате чего существенно возрастает урожай зерна и в гораздо меньшей степени – содержание в нем азота.
Симбиотическая фиксация атмосферного азота
зернобобовыми культурами
Зернобобовые культуры являются одним их основных источников производства растительного белка. Но для обеспечения высокой урожайности и белковой продуктивности этих культур необходимо достаточное азотное питание, источником которого могут служить доступные растениям формы почвенного азота, фиксированный клубеньковыми бактериями азот атмосферы и азотсодержащие органические и минеральные удобрения. При этом как с экономических, так и с экологических позиций в большинстве стран мира предпочтение отдается азоту биологическому.
Наблюдения за формированием симбиотических отношений в посевах зернобобовых культур, что в естественных условиях на черноземах лесостепи юга Западной Сибири среди изученных растений корневые клубеньки образуются у гороха, вики, чечевицы и кормовых бобов.
Это свидетельствует о том, что в почве присутствуют только клубеньковые бактерии Rhizobium leguminosarum, а специфичных для фасоли, нута, люпина и сои азотфиксаторов в почве нет. Поэтому формирование симбиотического аппарата у последних возможно лишь при заражении почвы соответствующими бактериями.
Известно, что конечный результат взаимодействия растений с клубеньковыми бактериями определяется величиной симбиотического аппарата, его удельной азотфиксирующей активностью и продолжительностью активного симбиоза.
С использованием этих показателей нами и проведена оценка азотфиксирующей способности различных зернобобовых культур, обобщенные результаты которой представлены в таблице 6.
Таблица 6 – Средние характеристики симбиоза клубеньковых бактерий
с зернобобовыми культурами
| Культура | Период вегетации, дней | Период симбиоза, дней | Масса клубень-ков, мг/раст. | Нитрогеназная активность, мкг N2 | Уровень фиксации азота за вегетацию, кг/га | |
| на 1 г клубень-ков за час | на 1 расте-ние за час | |||||
| Чечевица | 80 | 35 | 160 | 310 | 45 | 40 |
| Вика | 80 | 45 | 160 | 330 | 50 | 60 |
| Горох | 75 | 40 | 190 | 475 | 90 | 50 |
| Бобы | 110 | 65 | 410 | 370 | 150 | 75 |
| Фасоль | 80 | 55 | 310 | 440 | 130 | 50 |
| Нут | 100 | 65 | 320 | 330 | 100 | 50 |
| Соя | 100 | 75 | 360 | 490 | 150 | 90 |
| Люпин | 100 | 70 | 630 | 370 | 210 | 110 |
Установлено, что по продолжительности симбиоза зернобобовые культуры имеют существенные различия, связанные главным образом с длиной вегетационного периода. Показано, что у изученных культур этот показатель в среднем меняется от 30-40 дней у чечевицы до 70-80 и более дней (в зависимости от условий увлажнения) у сои и люпина. При этом в зависимости от срока сева, обусловленного биологическими особенностями рассматриваемых культур, период максимальной азотфиксирующей активности клубеньков смещается со второй половины июня (культуры раннего срока сева) на июль и даже на начало августа (нут, соя, люпин узколистный). Следовательно, даже в один и тот же год клубеньковые бактерии разных культур работают в различных условиях увлажнения и температуры, что также вносит существенные поправки в уровень усвоения ими азота воздуха.
По количеству и массе корневых клубеньков зернобобовые культуры также существенно различаются. Так, если у гороха, вики и чечевицы наблюдаются близкие показатели (по 8-15 клубеньков на 1 растении со средней массой одного клубенька в пределах 5-10 мг), то у кормовых бобов, сои и фасоли эти показатели в 2,0-2,5 раза выше. Отличительной особенностью люпина является формирование небольшого числа (4-8) очень крупных (150-250 мг) клубеньков.
По удельной азотфиксирующей активности клубеньков изученные культуры различаются в меньшей степени: минимальна она у чечевицы (310 мкг азота / г клуб.· час), а максимальна – у сои (490 мкг азота / г клуб. · час).
В конечном итоге по азотфиксирующей активности растений зернобобовые культуры можно расположить в следующий ряд (в порядке возрастания): чечевица – вика – горох – нут – фасоль – бобы и соя – люпин. При этом минимальные и максимальные значения прихода азота атмосферы в агроценозы различаются в 2-3 раза.
Анализ экспериментальных данных показывает, что с ростом массы клубеньков на 1 растении их азотфиксируюшая активность повышается, если увеличение массы клубеньков связано с фенологией растения. Если же измерения проведены в одно и то же время, а различия в формировании симбиотического аппарата обусловлены условиями произрастания растений или применяемым штаммом бактерий удельная активность клубеньков понижается (рис.5.).
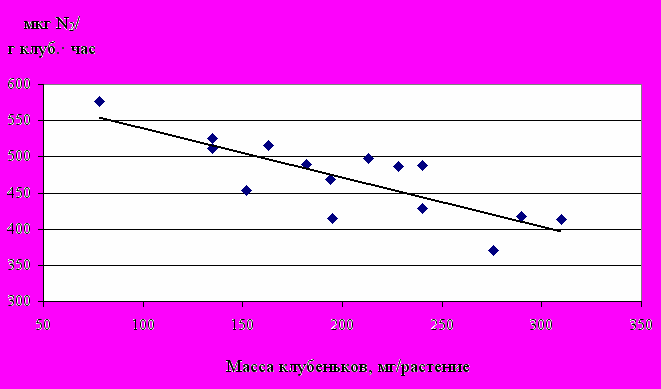
Рис. 5. Азотфиксирующая активность клубеньков гороха в зависимости от их массы на 1 растении (в среднем за 1996-1998 гг.)
В частности, у растений гороха, согласно рассчитанному по 3 летним экспериментальным данным уравнению регрессии y = -0,68x + 607,4 (n = 15; r = 0,81±0,35), с увеличением массы клубеньков у одного растения на 1 мг, удельная активность связывания азота снижается в среднем на 0,68 мкг/г клубеньков.
Особенно резко падает активность клубеньков в условиях дефицита влаги (1997 год). Это связано, по-видимому, с тем, что в процессе онтогенеза рост массы клубеньков сопровождается нарастанием площади листьев и значительной активизацией фотосинтеза, в то время как при изменении экологических факторов, способствующих усилению клубенькообразования, ассимиляция растениями СО2 увеличивается в меньшей степени и ухудшается обеспечение клубеньков продуктами фотосинтеза.
Аналогичные зависимости, характеризующиеся достаточно высокими коэффициентами корреляции, характерны и для других зернобобовых культур (табл. 7).
Таблица 7 – Зависимости удельной нитрогеназной активности клубеньков зернобобовых культур (y) от их массы на одном растении (x)
| Культура | Коэффициент корреляции | Уравнение регрессии (1996-1998 гг.) | |||
| 1996 г. | 1997 г. | 1998 г. | 1996-1998 гг. | ||
| Чечевица | -0,81 | -0,88 | -0,97 | - 0,92 | y = -0,93x + 471,6 |
| Вика | -0,52 | -0,83 | -0,61 | - 0,81 | y = -0,68x + 444,6 |
| Горох | -0,95 | -0,91 | -0,91 | - 0,80 | y = -0,68x + 607,4 |
| Бобы | -0.97 | -0.99 | -0.96 | - 0,96 | y = -0,26x + 484,7 |
| Фасоль | -0,93 | -0,85 | -0.84 | - 0,95 | y = -0,51x + 607,9 |
| Нут | -0,94 | -0,92 | -0,93 | - 0,90 | y = -0,24x + 403.9 |
| Соя | -0,99 | -1,00 | -0.86 | - 0,93 | y = -0,57x + 711,4 |
| Люпин | -0,95 | -0,96 | -0,98 | - 0,87 | y = -0,28x + 546,1 |