
Развивающийся мозг как эндокринный источник серотонина у крыс в онтогенезе
| Вид материала | Автореферат |
- Развитие иммунной системы в онтогенезе крыс: нейроэндокрино-иммунные взаимодействия, 310.15kb.
- Автор: Черников Василий, 2192.93kb.
- Влияние нарушения маточно-плацентарного кровообращения на структуру лимбической системы, 5266.12kb.
- Роль β-адренорецепторов в регуляции водно-солевого обмена у белых крыс, 748.01kb.
- «Анатомия цнс» Тема: Спинной мозг внешнее строение, 137.44kb.
- Метиленовый синий: риск развития серотонинового синдрома при совместном применении, 17kb.
- Укажите источник развития нервной системы, 1342.7kb.
- Женский мозг и мужской мозг(Серж Гингер), 110.44kb.
- Культурное разнообразие в обществе, институты и экономическое развитие: вопросы теории, 129.28kb.
- Что может наш мозг? Мозг и похудение, 148.35kb.
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт биологии развития им. Н.К. Кольцова
На правах рукописи
НАСЫРОВА ДИАНА ИЛЬДАРОВНА
Развивающийся мозг как эндокринный источник серотонина
у крыс в онтогенезе
Специальность 03.00.13 – физиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва 2007
Работа выполнена в лаборатории гормональных регуляций
Института биологии развития им. Н.К. Кольцова РАН
Директор института – доктор биологических наук,
профессор Николай Дмитриевич Озернюк
Научный руководитель:
академик Михаил Вениаминович Угрюмов
Официальные оппоненты:
доктор биологических наук, профессор
Борис Николаевич Манухин
доктор биологических наук, профессор
Владимир Вячеславович Раевский
Ведущая организация:
Институт эндокринологии Эндокринологического научного центра ФУ
Агентства по новым медицинским технологиям МЗ РФ
Защита состоится «____» мая 2007 г. в _________часов.
На заседании Диссертационного совета Д 002.238.01 при Институте биологии развития
им. Н.К. Кольцова РАН по адресу 119334, Москва, ул. Вавилова, д. 26.
Факс: (495) 135-80-12
E-mail: volina46@bk.ru
С диссертацией можно ознакомиться в библиотеке Института биологии развития им. Н.К. Кольцова РАН (119334, Москва, ул. Вавилова, д.26.)
Автореферат разослан «_____» апреля 2007 г.
Ученый секретарь
Диссертационного совета,
кандидат биологических наук Е.В. Волина
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Одной из наиболее актуальных задач современной физиологии является изучение механизмов нейроэндокринной регуляции, обеспечивающей интеграцию и поддержание постоянства внутренней среды взрослого и развивающегося организма [Угрюмов, 1999]. Важно подчеркнуть, что действие одних и тех же химических сигналов, вырабатывающихся элементами нейроэндокринной системы, на клетки и органы-мишени качественно различаются во взрослом и развивающемся организме. Во взрослом организме действие этих физиологически активных веществ (ФАВ) кратковременно и необратимо, тогда как в развивающемся организме они выступают в роли индукторов развития, оказывая необратимое морфогенетическое влияние на развивающиеся мишени [Lauder, 1993; Ugrumov, 1997]. Исследования становления нейроэндокринной регуляции в онтогенезе актуальны как для фундаментальной науки – физиологии и биологии развития, так и для медицины, поскольку нарушения метаболизма ФАВ на ранних стадиях развития нередко приводят к развитию врожденных заболеваний.
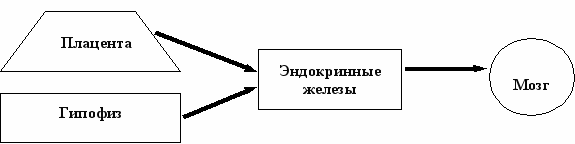
В начале 80-х годов прошлого столетия была сформулирована концепция гуморальной регуляции развития целостного организма и его нейроэндокринной системы, важнейшим звеном которой у млекопитающих является мозг и, главным образом, гипоталамус [Мицкевич, 1978; Dörner, 1981]. По этой концепции следовало, что мозг с момента образования нервной трубки и до окончания дифференцировки и формирования специфических нейрональных связей не оказывает влияния на развивающийся организм (Схема 1А). У незрелорождающихся животных (грызуны) этот период охватывает вторую половину внутриутробного развития. Учитывая, что именно в это время гормоны оказывают необратимое действие на мишени, был сделан вывод о том, что развитие мозга и висцеральных органов-мишеней контролируется гормонами рано развивающихся в онтогенезе эндокринных желез [Jacobson M., 1991]. И только после окончания дифференцировки нейронов и становления их эфферентных и афферентных связей формируется гипоталамический контроль функций гипофиза, а опосредованно и периферических эндокринных желез (Схема 1Б).
А
 Б
Б
Гипофиз
Схема 1. Нейроэндокринные регуляции у незрелорождающихся животных в пренатальном (А) и раннем постнатальном (Б) периодах развития (Dörner, 1977; Dörner, 1981)
Результаты последних исследований развития мозга и формирования нейроэндокринной регуляции в онтогенезе противоречат общепринятой концепции. Так, например, для нейронов, продуцирующих гонадотропин-рилизинг гормон, было показано, что они образуются одновременно с железистыми клетками эндокринных желез и сразу после этого приобретают способность к спонтанному или стимулированному выделению этого нейрогормона [Sétáló et al., 1978; Huhtaniemi, Warren, 1990]. Другими словами, дифференцирующиеся нейроны приобретают свойства секреторных клеток задолго до разрастания дендритов и аксонов и
 формирования межнейрональных синаптических связей (Схема 2).
формирования межнейрональных синаптических связей (Схема 2).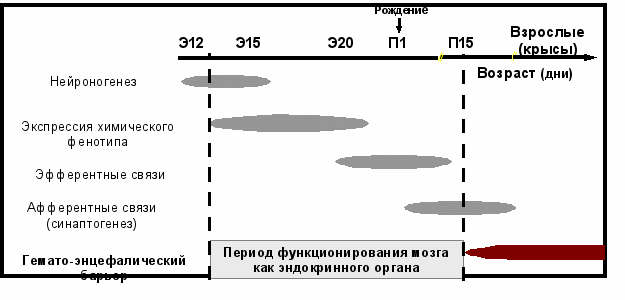
Схема 2. Хронология дифференцировки нейрона и развития мозга крысы в пре- и постнатальном периодах развития (Ugrumov, 2004).
Э – эмбриональный день; П – постнатальный день.
Учитывая отсутствие гематоэнцефалического барьера (ГЭБ) в период функционирования нейронов как секреторных клеток, т. е. до формирования межнейрональных связей [Sachs, Jonsson, 1996], можно предположить, что мозг именно в этот период развития играет роль эндокринного органа. Если гипотеза верна, то гуморальная регуляция развития мозга и висцеральных органов, включая эндокринные железы, должна быть взаимной, а не однонаправленной, как это утверждается общепринятой концепцией (Схема 3).
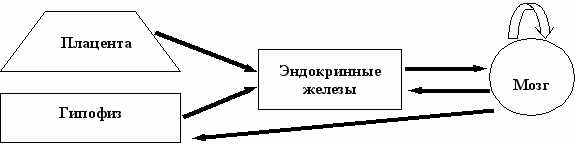
Схема 3. Замкнутая система регуляции в пренатальном и раннем постнатальном периодах онтогенеза (Угрюмов, 2004; Ugrumov, 2005)
Ранее нами было показано, что концентрация гонадотропин-рилизинг гормон и дофамина в крови высока в перинатальном периоде онтогенеза (до формирования ГЭБ) и падает практически до нуля после формирования ГЭБ. Кроме того, при удалении in utero переднего и среднего мозга плодов крыс уровень этих физиологически активных веществ в крови снижается на 50% [Ugrumov et al., 2005; Лаврентьева и др., 2006], что подтверждает гипотезу. В совокупности эти данные рассматриваются как серьезные аргументы в пользу гипотезы о развивающемся мозге как эндокринном органе.
Серотонин – один из важнейших и функционально значимых физиологически активных веществ в организме. В отличие от дофамина и гонадотропин-рилизинг гормон, серотонин в больших количествах синтезируется не только в мозге [Steinbusch, Nieuwenhuys, 1983; Montange, Calas; 1988], но и в энтерохромаффинных (ЭХ) клетках желудочно-кишечного тракта, особенно двенадцатиперстной кишки (12-пк), и поступает в общую систему циркуляции из периферических источников во все периоды онтогенеза [Pearse, 1974; Dockray, 1977; Rindi, 2004]. Учитывая это, серотонин использован как качественно новый маркер для проверки гипотезы о том, что до формирования ГЭБ мозг является значимым источником серотонина в крови.
У взрослых животных серотонин играет роль нейротрансмиттера в мозге и гормона на периферии [Fuller, Clemens, 1981; Weiner et al., 1988; Ugrumov, 1992, 1997]. Серотонинергическая система мозга участвует в регуляции циклов сна/бодрствования, общей двигательной активности, разных форм эмоционального поведения, процессов памяти и обучения. Сбои в ее работе ведут к развитию таких психических расстройств, как маниакально-депрессивный психоз, шизофрения, депрессии, тревожные состояния, нарушения пищевых мотиваций [Shopsin et al., 1984; Blier et al., 1999; Vickers et al., 2004; Lowry et al., 2005; Hough et al., 2006; Shmitt et al., 2006]. Периферическая серотонинергическая система участвует в регуляции в основном сердечно-сосудистой и пищеварительной систем, нарушения в ее функционировании приводят к серьезным патологиям. [De Clerck et al., 1990; Hayreh et al., 1999; MacLean et al., 2000; Yusuf et al., 2003; Eddahibi et al., 2006].
В определенные, так называемы «критические» периоды онтогенеза серотонин оказывает необратимое влияние на периферические клетки и органы-мишени, выступая в качестве индуктора развития [Azmitia et al., 1990; Whitaker-Azmitia, 1993; Lauder, 1993].
Принято считать, что до формирования синаптических контактов серотонин, синтезирующийся в ядрах шва головного мозга оказывает паракринное действие, влияя на дифференцировку нейронов-мишеней, в том числе и серотонинергических нейронов (ауторегуляция). В нашей лаборатории была выдвинута гипотеза, согласно которой помимо паракринной регуляции, серотонин из мозга попадает в кровь и оказывает эндокринное действие.
Цель исследования. Цель настоящей диссертационной работы состояла в проверке гипотезы о том, что развивающийся мозг является эндокринным источником серотонина у крыс.
Задачи исследования.
1. Оценить изменение уровня серотонина у крыс в онтогенезе:
а) в мозге и двенадцатиперстной кишке
б) в крови
2. Оценить изменение уровня серотонина в крови после «выключения» синтезирующих его нейронов мозга с помощью:
а) микрохирургической модели (декапитация плодов in utero)
б) фармакологической модели (ингибирование синтеза в мозге)
Научная новизна. Впервые проведен сравнительный анализ секреторной активности важнейших серотонин-продуцирующих систем организма у крыс в онтогенезе. Показано как изменяется концентрация и содержание серотонина в крови, мозге и 12-пк до и после формирования ГЭБ. Эти данные позволили предположить, что мозг до формирования ГЭБ является одним из источников серотонина в общей системе циркуляции. Кроме того, для проведения такой работы впервые была исследована динамика объема плазмы крови у крыс в онтогенезе. Эти результаты позволяют, зная концентрацию физиологически активных веществ в плазме, определить их общее содержание и, таким образом, оценить эффективность секреторных органов.
Впервые разработана модель фармакологического выключения синтеза серотонина в мозге при помощи стереотаксического однократного введения пара-хлорфенилаланина - ингибитора триптофангидроксилазы, первого скорость-лимитирующего фермента синтеза серотонина. Экспериментально была подобрана доза ингибитора, вызывающая максимальное ингибирование синтеза серотонина в мозге и не влияющая на синтез серотонина на периферии.
Впервые получено прямое доказательство того, что серотонин мозгового происхождения до формирования ГЭБ попадает в общую систему циркуляции и вносит существенный вклад в создание физиологически активной концентрации. Полученные данные подтверждают гипотезу о том, что нейроны до формирования межнейрональных связей и ГЭБ функционируют как секреторные клетки, а мозг в целом как эндокринный орган. Учитывая то, что существует мощный источник серотонина на периферии, серотонин был качественно новым маркером для проверки высказанной гипотезы.
Научная и практическая значимость работы.
- Подтвержден гипотеза о том, что развивающийся мозг до формирования ГЭБ в онтогенезе функционирует как эндокринный орган, внося существенный вклад в создание и поддержание «физиологически активной» концентрации ФАВ в крови, что принципиально меняет существующую концепцию развития организма.
- Работа вносит важный вклад не только в экспериментальную физиологию и нейроэндокринологию развития, но и в перинаталогию - область клинической медицины, изучающую развитие организма в пренатальном и раннем постнатальном периодах онтогенеза в норме, а также в патологии, связанной с нарушением развития. Результаты этой работы особенно важны для понимания механизмов патогенеза ряда врожденных заболеваний.
- Полученные данные используются для преподавания нейроэндокринологии и нейробиологии на старших курсах естественнонаучных и медицинских факультетов парижского Университета им П. и М. Кюри, Московской медицинской академии им. И.М. Сеченова и ряда других российских и зарубежных ВУЗов.
Апробация работы: Результаты диссертационной работы были представлены и обсуждены:
- на международных симпозиумах: International Symposium “Neuron differentiation and plasticity-regulation by intercellular signals”, Москва, 2003; I Съезд физиологов СНГ, Сочи, Дагомыс, 2005 г.; XIII международное совещание по эволюционной физиологии, Санкт-Петербург, 2006 г.; 5 th Forum of European neuroscience, Vienna, 2006 г.;
- на совместных коллоквиумах и семинарах: лаборатории гормональных регуляций Института биологии развития им. Н.К. Кольцова РАН, лаборатории нейрогистологии им. Б.И. Лаврентьева Института нормальной физиологии им. П.К. Анохина РАМН, кафедры нормальной физиологии человека и животных Казанского государственного медицинского университета, на конференции молодых ученых ИБР РАН, Москва, 2005.
Публикации. По теме диссертации опубликовано 7 работ, из них 3 статьи в рецензируемых журналах, 4 публикации в сборниках тезисов.
Структура и объем диссертации. Диссертация состоит из введения, обзора литературы, описания материалов и методов исследования, результатов, обсуждения, выводов и списка литературы, включающего 183 источника. Работа изложена на 137 страницах машинописного текста, содержит 39 рисунков, 5 таблиц и 2 схемы.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Животные. Исследование проведено на крысах Вистар: плодах на 18-ый день эмбрионального периода развития (Э18), 3-й день постанатального периода развития (П3), П4, П15, П16 и П30. Животных всех возрастов, кроме Э18, дифференцировали по полу, по аногенитальному расстоянию. Для получения самок с датированным сроком беременности к 3-4 месячным самкам весом 200-250 г вечером подсаживали самцов, а утром брали влагалищные мазки. День обнаружения спермы в мазке считали первым днем беременности. День рождения крысят считали П1.
Эксперименты
Измерение объема плазмы крови у крыс в онтогенезе. Все манипуляции с животными проводили под пентобарбиталовым наркозом (40 мг/кг веса). Объем плазмы определяли методом «разведения красителя», используя азокраситель Evans Blue, Т-1824 [Gregersen et al, 1944; Nielsen N., Nielsen M., 1962]. Краситель разводили физиологическим раствором до концентрации 0,5% и вводили в бедренную вену из расчета 3 мг/кг веса. Кровь брали из сердца через 1 мин после введения раствора красителя. Через 10 минут плазму отделяли центрифугированием при 3000 об/мин в течение 10 минут. Собранную плазму десятикратно разводили физиологическим раствором и определяли оптическую плотность на спектрофотометре Specol 211 ("Carlzeiss Jena", Германия) при длине волны 640 нм. Для установки прибора на ноль использовали пробу без красителя, которую называли "чистой плазмой".
Дальнейшее определение объема плазмы проводили с использованием стандарта, для приготовления которого в известный объем чистой плазмы, десятикратно разведенной физиологическим раствором, вводили объем красителя, равный объему, введенному животному. После этого определяли оптическую плотность стандарта. Объем плазмы животного определяли по формуле:
V=P × V0 / P0
где P – оптическая плотность плазмы животного; P0 – оптическая плотность стандарта; V0 – известный объем чистой плазмы, использованный для приготовления стандарта.
Декапитация плодов крыс in utero. Помимо интактных животных, в экспериментах использовали плодов после декапитации in utero. Для этого самок на 18-ый день беременности под пентобарбиталовым наркозом (40 мг/кг веса) подвергали лапаротомии, высвобождали рог матки, пальцами фиксировали голову плода. Прошивали стенку матки и плодного пузыря по кругу в области головы плода и через разрез в стенке матки выводили голову плода, после чего, затягивая лигатуру вокруг шеи, отделяли голову. С контрольными плодами никаких манипуляций не проводили. По окончании операций на плодах брюшную полость самки зашивали и плоды находились в организме матери до Э21.
Стереотаксическое введение п-ХФА в боковые желудочки мозга. Для ингибирования синтеза серотонина у крыс in vivo использовали Д,Л-пара-хлорфенилаланин (п-ХФА) (pCPA, Sigma), конкурентный ингибитор триптофангидроксилазы (ТпГ).
Система для внутрижелудочкового введения представляла собой гамильтоновский шприц объемом 10 мкл, фиксированный на стереотаксическом аппарате. Крысят под холодовым наркозом фиксировали на стереотаксическом аппарате под углом в 15°.
100 и 150 мкг п-ХФА растворяли в 2 мкл 0,9% NaCl и вводили в латеральный желудочек мозга по следующим стереотаксическим координатам по отношению к брегме: 1.4 мм латерально, 2,0-2,5 мм вглубь [Ugrumov et Mitskevich, 1980]. Введение производили однократно со скоростью 1мкл/мин. Контрольным животным вводили 2 мкл 0,9% NaCl. Животных после инъекции содержали 24 часа в стандартных лабораторных условиях, после чего выделяли мозг, 12-пк и собирали кровь.
Системное введение п-ХФА. При системном введении использовали те же количества п-ХФА, что и при стереотаксическом введении: 100 и 150 мкг на животное. п-ХФА растворяли в 500 мкл 0,9% NaCl и внутрибрюшинно вводили животным однократно. Контрольным животным вводили 500 мкл 0,9% NaCl. Животных после инъекции содержали 24 часа в стандартных лабораторных условиях, после чего выделяли мозг, 12-пк и собирали кровь.
Взятие и обработка материала
Мозг и 12-пк. Для определения содержания серотонина ткань целого мозга и каудальный фрагмент 12-пк гомогенизировали при помощи ультразвукового гомогенизатора (L-666, MSE, Англия) в 0,1 Н HClO4, добавляли 10 мкл 0,2 Н HClO4, содержащей 1 нг альфа-метилсеротонина (АМГТ) и центрифугировали 20 минут при 14000 об/мин. Супернатант собирали и хранили при -80C до измерения на хроматографе.
Кровь. Кровь (300 мкл) брали из сердца и помещали в пробирку, содержащую 30 мкл 5% раствора ЭДТА (Sigma) и 10 мкл 10% раствора метабисульфита натрия (Sigma). Для определения серотонина в крови образцы центрифугировали при 1000 об/мин 10 минут для получения плазмы, обогащенной тромбоцитами. Для разделения тромбоцитов и плазмы проводили повторное центрифугирование при 3000 об/мин 10 мин.
В каждую пробу плазмы добавляли одну десятую объема 1 Н HClO4 до конечной концентрации 0,1 Н, а также 1 нг АМГТ (Sigma) в 10 мкл 0,1 Н HClO4. В каждую пробу с тромбоцитами добавляли фиксированный объем 0,1 Н HClO4, 10 мкл (1 нг) АМГТ и все пробы центрифугировали 20 минут при 14000 об/мин. Супернатант собирали и хранили при –80С до определения на хроматографе.
Для определения серотонина в тромбоцитах из каждой пробы плазмы, обогащенной тромбоцитами, брали 10 мкл для подсчета тромбоцитов в камере Горяева. Затем при помощи высокоэффективной жидкостной хроматографии определяли уровень серотонина в супернатанте, содержащем тромбоцитарный серотонин. Содержание серотонина оценивали на на 109 тромбоцитов.
Высокоэффективная жидкостная хроматография с электрохимической детекцией. После 10-минутного центрифугирования при 1000 об/мин супернатанты исследовали при помощи обратно-фазной высокоэффективной жидкостной хроматографии с электрохимической детекцией (Amperometric detector LC-4B, Bioanalytical Systems, США) при потенциале 850 мВ. Подвижной фазой служил 0,1 М цитратно-фосфатный буфер, содержащий 0,3 мМ октансульфоната натрия (Sigma), 0,1 мМ ЭДТА (Sigma) и 8 % ацетонитрила (Sigma) (pH 3,2). В качестве стандарта использовали свежеприготовленный раствор следующего состава (нг/мл): серотонин (5-гидрокситриптамин, 5-ГТ), 100, АМГТ 100. Концентрацию каждого компонента вычисляли путем сравнения величины пика в пробе с величиной пика в стандарте по следующей формуле:
С 5-ГТстанд hАМГТстанд
С5-ГТпроб = hпроб * ----------------- * ----------------- ,
hстанд hАМГТтпроб
где С5-ГТпроб – концентрация серотонина в пробе, hпроб – высота пика серотонина в пробе, hстанд – высота пика серотонина в стандарте, С 5-ГТстанд – концентрация серотонина в стандарте, hАМГТстанд – высота пика АМГТ в стандарте, hАМГТпроб – высота пика АМГТ в пробе.
Содержание серотонина в крови рассчитывали как произведение его концентрации на объем плазмы, содержание в мозгу и 12-пк – как произведение концентрации на вес выделенной ткани.
Расчет средней скорости изменения концентрации и содержания серотонина в тканях и крови в изученные периоды проводили по формуле:
К 2 - К 1
VК = ----------------- ,
n
где VК - средняя скорость изменения концентрации в определенный возрастной период, К2 - концентрация серотонина на более поздний из выбранных сроков, К1 - концентрация серотонина на более ранний из выбранных сроков, n – количество дней между двумя выбранными сроками.
Аналогично рассчитывали среднюю скорость изменения содержания серотонина - VС
Статистическая обработка результатов
Статистический анализ полученных результатов проводили с помощью непараметрического критерия Уилкоксона.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Объем плазмы крови у крыс в онтогенезе
Концентрация серотонина в крови определяется скоростью его поступления из синтезирующих органов, скоростью ферментативного распада, а также объемом плазмы. Если концентрация серотонина в крови отражает его функциональную активность, то содержание - является показателем эффективности в целом синтезирующих его источников. Содержание серотонина в плазме рассчитывают как произведение концентрации на объем плазмы. Несмотря на огромное значение крови как транспортной системы, вовлеченной в общий метаболизм и регуляцию развивающегося организма, отсутствуют данные об изменении объема плазмы в онтогенезе млекопитающих. Учитывая выше сказанное, нами была исследована динамика объема плазмы у крыс в онтогенезе.
Объем плазмы на Э18 равен 0,197 ± 0,017 мл, а к Э21 он увеличивается до 0,380 ± 0,059 мл (в 1,93 раз) (Рис. 2). В перинатальном периоде развития крыс объем плазмы возрастает в 1,4 раза, достигая 0,520 ± 0,061 мл. Объем плазмы на П15 в 2,1 раза выше, чем на П3. В период с П15 по П30 объем плазмы у крыс увеличивается с 1,080 ± 0,085 мл до 3,701 ± 0,420 мл (в 3,4 раза). Не обнаружены половые различия по объему плазмы во всех возрастных группах (Рис. 1).
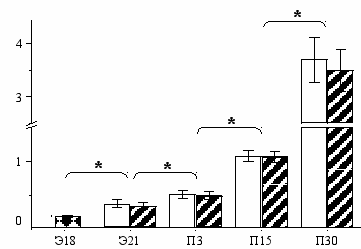
Р

 ис. 1. Объем плазмы крови, мл (по оси ординат) в эмбриональном (Э) и постнатальном (П) периодах развития (по оси абсцисс) у крыс: - самцы, - самки, - самцы + самки.
ис. 1. Объем плазмы крови, мл (по оси ординат) в эмбриональном (Э) и постнатальном (П) периодах развития (по оси абсцисс) у крыс: - самцы, - самки, - самцы + самки. * достоверные различия между выбранными параметрами, P<0.05.
Поскольку объем рассчитывается в миллилитрах, а вес тела - в граммах, чтобы привести эти параметры к одной единице измерения, мы взвешивали 1 мл плазмы, взятой у животных всех исследованных возрастов, и определили, что он соответствует 1 г. Это позволило нам рассчитать процент массы плазмы от веса тела.
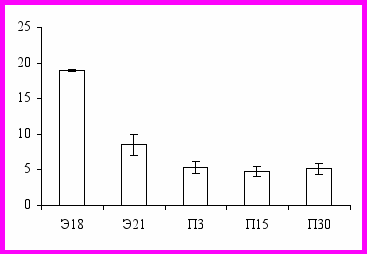
Рис. 2. Процентное соотношение массы плазмы крови и веса тела, % (по оси ординат) у крыс в эмбриональном (Э) и постнатальном (П) периодах развития (по оси абсцисс).
В эмбриональном периоде на Э18 объем плазмы составляет 19% от веса тела (Рис. 2). С Э18 по Э21 это соотношение снижается в два раза, что происходит за счет значительного прироста веса тела (в пять раз). Ранее было показано, что наиболее интенсивное увеличение веса тела происходит в эмбриональном и перинатальном периодах развития крыс (Западнюк, 1974; Нигматуллина и др., 1988; Szakmary et al., 2001), что согласуется с результатами данного исследования. Начиная с П3, это соотношение меняется незначительно (5,37 - 5,15%), что совпадает с результатами, полученными для новорожденных телят, взрослых крыс и кроликов (Prince, 1982; Gillen et al., 1994; Quigley et al., 1998). Таким образом, объем крови коррелирует с весом тела и степенью развития кровеносной системы.
Развитие центральной и периферической серотонин-продуцирующих
систем у крыс в онтогенезе
Изменение концентрации и содержания серотонина в мозге у крыс в онтогенезе.
Период с Э21 по П4. По нашим данным концентрация серотонина в мозге в этот период увеличивается в два раза (Рис.3А). К этому времени серотонинергические нейроны мозга становятся в значительной степени функционально зрелыми. Так, мРНК серотонинового транспортера (СТ) впервые выявляется в некоторых отделах мозга, в том числе в ядрах шва у крыс на Э11 [Hansson, 1998], а у мышей - на Э10 [Schroeter, Blakely, 1996]. В период с Э21 по П4 экспрессия мРНК СТ у крыс увеличивается и она выявляется во многих отделах мозга. Параллельно дифференцировке серотонинергических нейронов происходит экспрессия рецепторов к серотонину и формирование ферментативной системы его деградации. Так, на Э16 уже обнаружены 5-ГТ1А-, 5-ГТ1D-, 5-ГТ1С-, 5-ГТ1B-, 5-ГТ4-рецепторы. Их количество увеличивается до конца пренатального периода [Waeber, 1996; Bolaños-Jiménez, 1997; Moiseiwitsch, 1995]. Синтез мРНК МАО А, фермента деградации серотонина, впервые обнаружен у мышей на Э12. Уровень синтеза серотонина прогрессивно увеличивается до Э14 и остается на этом высоком уровне до П4 [Luque, 1995; Vitalis, 2002].
Содержание серотонина в этот период увеличивается почти в 5 раз, что объясняется продолжающимся прорастанием аксонов ядра шва в отделы мозга-мишени и накоплением в них серотонина [Wallace, Lauder 1983; Ugrumov, 1986] (Рис. 3Б).
Период с П4 по П16. Концентрация серотонина увеличивается в два раза (Рис. 3А), что совпадает по времени с достижением пика экспрессии 5-ГТ4-рецепторов. Незначительное количество этих рецепторов в эмбриогенезе говорит об их незначительной роли в развитии, однако в постнатальном периоде (особенно до П20) экспрессия этих рецепторов очень высока, что, вероятно, связанно с их участием в регуляции поведения. [Waeber, 1996]. Кроме того, в этот период прогрессивно снижается уровень экспрессии МАО А в мозге [Vitalis, 2002].
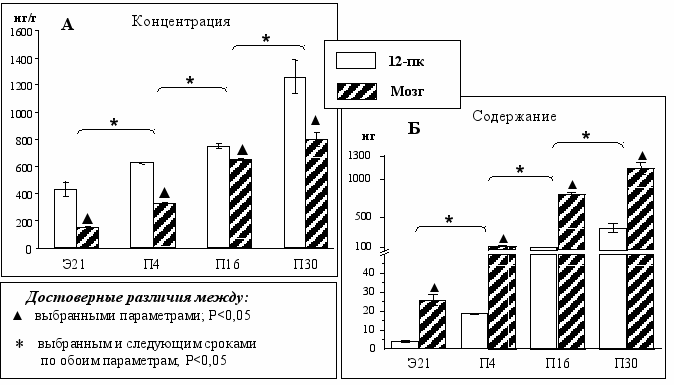
Рис. 3. Концентрация (А) и содержание серотонина (Б) в мозге и двенадцатиперстной кишке у крыс в онтогенезе.
В этот же период отмечено самое значительное увеличение содержания серотонина в мозге (в 6,5 раз) (Рис. 3Б). Это можно объяснить тем, что в течение первых трех недель после рождения продолжается иннервация мозга, в том числе коры [Mustari, Lund, 1976; Wallace, Lauder 1983], происходит формирование ГЭБ [Loizou, 1970], при этом серотонин перестает поступать из мозга в общую систему циркуляции. Все это позволяет полагать, что резкое увеличение уровня серотонина вызвано скорее «накоплением» серотонина в мозге, связанным с формированием ГЭБ, чем с усилением его синтеза. Это подтверждается еще и тем, что в этот же период активность ТпГ в мозге увеличивается незначительно по сравнению с увеличением содержания серотонина [Huether, 1992].
Период с П16 по П30. В этот период, в отличие от предыдущего, происходит небольшое увеличение, как концентрации, так и содержания серотонина в мозге, по сравнению с другими периодами (Рис. 3А, 3Б). К П30 уровень серотонина достигает уровня взрослых животных, что коррелирует с результатами предыдущих морфологических и биохимических работ [Wallace, Lauder 1983; Rodriguez, 1989; Takada, 1995]. Активность ТпГ в мозге в этот период увеличивается незначительно (в 1,1 раза), а к П30 она выходит на плато [Huether, 1992]. К П21 активность МАО А в мозге достигает уровня взрослых и в дальнейшем изменяется незначительно [Vitalis, 2002].
Таким образом, полученные результаты свидетельствуют о том, что эффективность мозга, как источника серотонина высока в эмбриональном периоде, резко увеличивается до П16 и затем снижается.
Изменение концентрации и содержания серотонина в двенадцатиперстной кишке у крыс в онтогенезе.
Период с Э21 по П4. По нашим данным концентрация серотонина в 12-пк увеличивается примерно в 1,2 раза, тогда как содержание - в 4,5 раза (Рис. 3А, 3Б). ЭХ клетки, являющиеся основным источником серотонина на периферии, впервые обнаруживаются у крыс на Э15, в это же время в них начинается синтез серотонина [Pearse, Takor-Takor, 1976; Branchek, Gershon, 1989].
Период с П4 по П16. Концентрация серотонина увеличивается также как и в предыдущий период. Увеличение уровня синтеза серотонина может достигаться за счет увеличения числа ЭХ клеток. Действительно, после рождения ЭХ клетки помимо 12-пк появляются и в других отделах желудочно-кишечного тракта, например, в желудке. Их число невелико и увеличивается только до 14-го постнатального дня [Hakanson et al., 1967; Pearse et al., 1976; Dockray, 1977; Pham et al., 1991].
Период с П16 по П30. В этот период при полностью сформировавшемся ГЭБ, наблюдается самое значительное (по сравнению с другими периодами) увеличение концентрации серотонина в 12-пк (Рис. 3А). К П30 концентрация равна 1,5 мкг/г, а вес животных в среднем - 70 г. Показано, что у взрослых крыс весом 200 г концентрация серотонина в 12-пк достигает 4,5 мкг/г [Kleinrok, 1984].
Полученные результаты свидетельствуют о том, что эффективность 12-пк, как источника серотонина не очень высока в эмбриональном периоде, но резко увеличивается после П16, т.е. после «выключения» мозга как источника серотонина в крови, обусловленного формированием ГЭБ. В этот период и позднее 12-пк - наиболее мощный источник серотонина в крови.
Изменение концентрации и содержания серотонина в крови у крыс в онтогенезе.
Период с Э21 по П4. По нашим данным в этот период наблюдается самое значительное повышение концентрации серотонина в плазме крови (Рис. 4А). Содержание серотонина в плазме увеличилось в 4 раза, в тромбоцитах - почти в 5 раз (Рис. 4А, 4Б). Вероятно, одна из причин такого резкого увеличения уровня серотонина в этот период состоит в том, что в отсутствие ГЭБ серотонин поступает в общую систему циркуляции не только из периферических источников, но и из мозга, который также вносит свой вклад в создание физиологически активной концентрации серотонина в крови – 10-8 М (уже на Э21), достаточной для адекватного действия на периферические клетки и органы-мишени. Показано, что у морских свинок 5-ГТ4-рецепторы в кишечнике чувствительны к серотонину в концентрации 10-8-10-7 М, а 5-ГТ3-рецепторы – к более высоким концентрациям - 10-7 -10-5 М [Craig, 1990a,b]. У грызунов на Э8 выявлена экспрессия мРНК 5-ГТ2В-рецептора в зачатке сердца, к Э9,5 экспрессия в области сердца увеличивается и продолжает расти [Choi, 1997]. На Э14 впервые выявляются 5-ГТ2В-рецепторы в желудке и кишечнике [Florica-Howells, 2000]. У мышей с нокаутом по гену 5-ГТ2В-рецептора 25 % эмбрионов погибают, остальные не доживают до половозрелого возраста [Nebigil, 2000]. Это объясняется тем, что именно через этот тип рецепторов серотонин регулирует морфогенез сердечно-сосудистой системы, в частности, дифференцировку и пролиферацию кардиомиоцитов.
Известно, что в норме 90% серотонина у человека и животных находится в тромбоцитах, которые высвобождают его при необходимости. Например, высвобожденный серотонин является одним из важнейших факторов при агрегации тромбоцитов и тромбообразовании [Da Prada et al., 1967; Da Prada et al., 1972], участвует в формировании ощущения боли, с последующей гипертензией и тахикардией [Kanai et al., 1993; Egermayer et al., 1999].
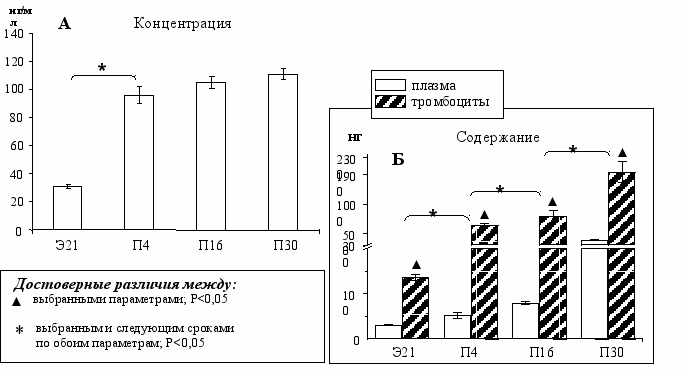
Рис. 4. Концентрация (А) и содержание серотонина (Б) в крови у крыс в онтогенезе.
Таким образом, в этот период стабильно высокая концентрация серотонина в плазме необходима для нормального развития животных. Этому способствует раннее появление всех необходимых типов рецепторов [Choi, 1997; Florica-Howells, 2000; Nebigil, 2000].
Период с П4 по П16. В плазме крови содержание серотонина увеличилось в 2,1 раза, а в тромбоцитах - в 1,3 раза. Уже к П4 концентрация серотонина в крови достигает уровня взрослых животных [Takada, 1995] и до П30 изменяется незначительно (Рис 4А).
Период с П16 по П30. Содержание серотонина в плазме увеличилось в 3,5 раза, в тромбоцитах - почти в 4 раза, т.е. содержание в плазме и тромбоцитах в этот период растет параллельно (Рис. 4Б). Известно, что у взрослых мышей содержание серотонина в тромбоцитах 3,2 мкг на 109 тромбоцитов [Lesurtel, 2006], нами показано, что на П30 содержание серотонина в тромбоцитах у крыс достигает такого же уровня.
Соотношение содержания серотонина в плазме и тромбоцитах не меняется в онтогенезе и составляет 1:10, что согласуется с результатами других работ по отдельным срокам [Takada, 1995]. Это может свидетельствовать о том, что уже в конце пренатального периода развития белки-транспортёры функционально активны и тромбоциты выполняют свою функцию – «депо» серотонина – также как у взрослых животных.
Средняя скорость изменения содержания серотонина
Средняя скорость изменения содержания серотонина в мозге и 12-пк, которая является показателем изменения их секреторной активности, в период с Э21 по П4 составляет 20 нг/дн. С П4 по П16 скорость увеличения содержания в мозге 60 нг/день, то есть в 3 раза выше, чем в предыдущий период, а в следующий период снижается до исходного значения (Рис. 5А).
В 12-пк средняя скорость увеличения содержания ниже, чем в мозге во все изученные периоды. Однако, в период с П16 по П30 наблюдается резкое увеличение этого показателя, что говорит об усилении секреторной активности 12-пк после формирования ГЭБ. Вероятно, это - компенсаторная реакция в ответ на прекращение поступления серотонина из мозга на периферию, направленная на поддержание физиологически активной (надпороговой) концентрации серотонина в крови. Именно серотонин периферического происхождения участвует в регуляции моторики желудочно-кишечного тракта, уровня глюкозы в крови, тромбообразования, сосудистого тонуса и артериального давления [Cote, 2004], сердечного ритма, силы сердечных сокращений [Frishman, Grewall, 2000; Manivet, 2000].
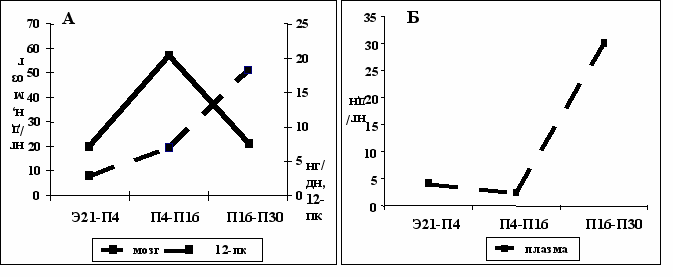
Рис. 5. Средняя скорость изменения содержания серотонина в мозге и 12 пк (А) и плазме (Б) у крыс в онтогенезе.
Средняя скорость изменения содержания серотонина в крови – показатель эффективности синтезирующих его источников. В крови в период становления ГЭБ (с П4 по П16) происходит достоверное снижение этого показателя почти в 2 раза, а затем резкое увеличение почти в 4 раза. Таким образом, несмотря на то, что содержание серотонина в крови растет, скорость его увеличения достоверно падает в период становления ГЭБ, когда мозг перестает быть источником серотонина в периферической крови.
Таким образом, полученные данные показывают, что во все периоды онтогенеза в создании физиологически активной концентрации серотонина в крови участвуют оба его источника, однако до формирования ГЭБ ведущую роль играет мозг, а после закрытия ГЭБ – 12-пк.
Мозг до формирования ГЭБ в онтогенезе - важнейший источник серотонина
в крови у крыс
Мы показали, что большое количество серотонина синтезируется не только периферическим источником – 12-пк, но и мозгом. Следовательно, теоретически мозг может быть источником серотонина в крови до закрытия ГЭБ. Эту гипотезу можно проверить, если создать модель, при которой мозг «выключается» как источник синтеза серотонина.
Существует два способа такого моделирования:
- Грубая неспецифическая модель микрохирургического удаления мозга, то есть внутриутробная декапитация плодов;
- Минимально травматичная специфическая модель фармакологического ингибирования синтеза серотонина в мозге с помощью п-ХФА.
Для получения прямого доказательства поступления серотонина из мозга в общую систему циркуляции до установления ГЭБ сначала была использована экспериментальная модель удаления мозга (декапитация плодов in utero), с измерением концентрации и содержания серотонина в периферической крови и 12-пк. Декапитация плодов плодов in utero, разработанная Жостом [Jost, 1947] в 70-80-е годы прошлого столетия успешно применялась для оценки формирования гипоталамического контроля функций гипофиза [Mitskevich and Rumyantzeva, 1972; Chatelain and Dupouy, 1981; Mitskevich and Sapronova, 1982], причем полученные с ее помощью результаты позднее были полностью подтверждены с помощью не столь травматичного и более селективного методического подхода – пассивной иммунизации к тем или иным нейрогормонам [Daikoku et al., 1981; Lalau et al., 1990; Zakharova et al., 2000].
Показано, что в опыте содержание серотонина в плазме и тромбоцитах плодов снизилось на 40 %, при этом содержание серотонина в 12-пк не изменилось (Рис. 6А, 6Б). Это указывает на то, что мозг является важнейшим источником серотонина в крови и согласуется с результатами наших предыдущих работ. Так, ранее нами было показано, что при удалении in utero переднего и среднего мозга плодов крыс, уровень гонадотропин-рилизинг гормона и дофамина в крови падал на 50% [Лаврентьева и др., 2004, Лаврентьева и др., 2006].
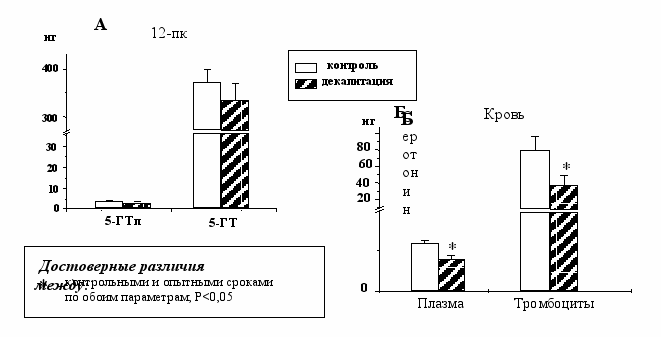
Рис. 6 Содержание серотонина и 5-гидрокситриптофана в 12-пк (А) и содержание серотонина в крови (Б) у плодов крыс после декапитации in utero
При декапитации «выключается» не только серотонинергическая система мозга, но и все остальные. Для «выключения» синтеза именно серотонина необходимо было разработать специфическую фармакологическая модель, которая позволила бы направленно заблокировать синтез серотонина. Кроме того, требовалось разработать фармакологическую модель ингибирования синтеза серотонина максимально в мозге при отсутствии ингибирования на периферии.
Поскольку такая работа проводилась впервые, то способ введения, оптимальное время и доза п-ХФА подбиралась эмпирически, учитывая, однако, результаты предыдущих работ [Koe, Weissman, 1966; Jequier, 1967; Koe, Weissman, 1968].
Путем ингибирования синтеза серотонина или в центральном источнике, или на периферии можно оценить, существенен ли вклад мозга в общее содержание серотонина в крови. Известно, что основные скопления серотонинергических нейронов и отростков располагаются около боковых желудочков мозга. Учитывая это, наиболее удобным оказалось ингибирование синтеза серотонина с помощью стереотаксических инъекций п-ХФА в мозг.
Ранее было показано, что при пероральном введении п-ХФА в дозе 300 мг/кг через 24 часа уровень серотонина в мозге снижался на 20%, а в 12-пк – на 55% [Koe, Weissman, 1966]. То есть при данном способе введения ингибирование было более эффективным в периферическом источнике. Также оценивались активность ТпГ и уровень серотонина в мозге крыс в зависимости от времени после внутрибрюшинной инъекции ингибитора в дозе 300 мг/кг [Jequier, 1967]. Ингибитор проникал через ГЭБ и спустя 4 часа обнаруживался в мозге в максимальной концентрации – 300 нмоль/г. Через 24 часа концентрация снижалась в 2 раза и через 6 дней падала до неопределяемой величины. Активность ТпГ оценивалась по уровню гидроксилированного триптофана, который максимально снижался через 24 часа после введения п-ХФА до 5% от нормы. Так как в норме 5-гидрокситриптофан (5-ГТп) сразу же после образования подвергается воздействию фермента неспецифической декарбоксилазы ароматических L-аминокислот и превращается в серотонин, то вслед за его снижением наблюдалось падение и уровня серотонина до 30% от нормы [Gal, 1970]. Таким образом, при внутрибрюшинном введении эффект в мозге был более выражен, по сравнению с пероральным.
Однако последующие работы показали, что при однократном внутрибрюшинном введении п-ХФА в дозе 300 мг/кг его максимальная концентрация через 6 часов после инъекции обнаруживается именно в кишечнике, в то время как в мозге она в 3 раза ниже [Jequier, 1967]. Следовательно, при данном введении ингибитора эффект максимально выражен в кишечнике, а не в мозге. Поэтому в настоящей работе использование внутрибрюшинного пути введения п-ХФА не позволило бы достигнуть поставленной цели.
Исходя из данных литературы, первая тестируемая доза п-ХФА равнялась 300 мг/кг веса мозга, т.е. 150 мкг на мозг крысы на П3. Сбор материала производили спустя сутки после инъекции, так как именно в это время по литературным данным был максимальный эффект после внутрибрюшинного введения п-ХФА [Gal, 1970].
При стереотаксическом введении п-ХФА в боковые желудочки мозга в дозе 300 мг/кг наблюдалось снижение содержания серотонина в мозге на 90% (Рис. 7А), что свидетельствует о проникновении п-ХФА почти во всю толщу мозга. При ведении любой дозы ингибитора в мозг, возможно диффундирование п-ХФА и попадание в общую систему кровообращения. Поэтому падение содержания серотонина в 12-пк на 17% (Рис. 7А) при этой дозе может быть обусловлено:
1) воздействием п-ХФА на уровень синтеза этого моноамина;
2) компенсаторным увеличением выделения серотонина из 12-пк в ответ на снижение его уровня в общей системе кровообращения.
Уровень 5-ГТп является показателем действия п-ХФА, так как его содержание напрямую зависит от активности ТпГ. Снижение его содержания в 12-пк на 50% (Рис. 8А) свидетельствует о воздействии ингибитора на синтез серотонина. Достоверное снижение содержания в 12-пк 5-гидроксииндолуксусной кислоты – основного метаболита серотонина - также свидетельствует о нарушении синтеза этого моноамина. Следовательно, снижение общего содержания серотонина в крови на 32% (Рис. 7Б) вызвано воздействием п-ХФА и на мозг, и на периферическую серотонин-продуцирующую систему. Поэтому для избегания периферического эффекта п-ХФА необходимо было снизить дозу вводимого ингибитора, чтобы уменьшить количество п-ХФА попавшего путем диффузии в общую систему кровообращения так, чтобы при разведении в крови он уже не смог бы подействовать на периферии.

Рис.7 Содержание серотонина у животных интактной, контрольной и опытной групп при введении п-ХФА в дозе 300 мг/кг в мозге и в двенадцатиперстной кишке (А), в крови (Б).
При введении п-ХФА в боковые желудочки мозга крысам на П3 в дозе 200 мг/кг (100 мкг на мозг) снижение содержания серотонина в мозге составило 68%, а в 12-пк – 12% (Рис. 9А). Однако уровень 5-ГТп на периферии не изменился (Рис. 8Б). Следовательно, п-ХФА не воздействовал на синтез серотонина в 12-пк, а снижение серотонина в 12-пк можно объяснить компенсаторным увеличением его выделения в ответ на снижение в общей системе кровообращения.
В крови суммарно содержание серотонина снизилось на 23%, падение в плазме составило 16%, а в тромбоцитах – 26% (Рис. 9Б). Вероятно, наиболее выраженное снижение содержания серотонина в тромбоцитах – компенсаторная реакция в ответ на снижение серотонина в плазме, возможная либо за счет усиления выделения, либо – уменьшения обратного захвата. Следовательно, мы получили прямое доказательство гипотезы о том, что мозг до формирования ГЭБ функционирует как эндокринный орган
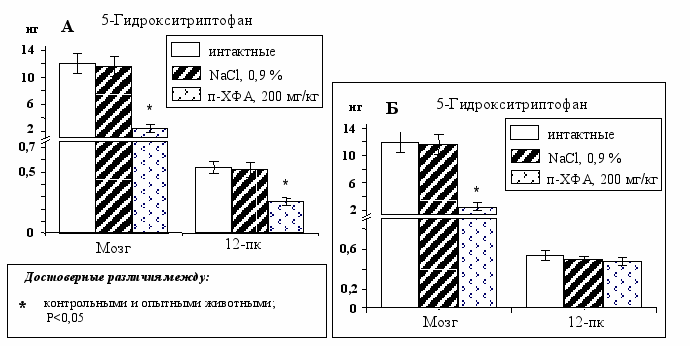
Рис. 8. Содержание 5-гидрокситриптофана у животных интактной, контрольной и опытной групп при введении п-ХФА в дозе 300 мг/кг (А) и 200 мг/кг (Б).
При оценке полученного снижения – 23% следует обратить внимание на то, что:
- это падение скомпенсировано примерно на 12% 12-пк;
- ингибирование синтеза серотонина в мозге произошло только на 60%
Учитывая эти факты, можно предположить, что вклад мозга значительно больше показанного нами.
Таким образом, разработанная фармакологическая модель направленного выключения синтеза серотонина в мозге у крыс подтвердила гипотезу о том, что до формирования ГЭБ значительная часть серотонина в общей системе циркуляции имеет мозговое происхождение, хотя мозг и не единственный источник его синтеза.
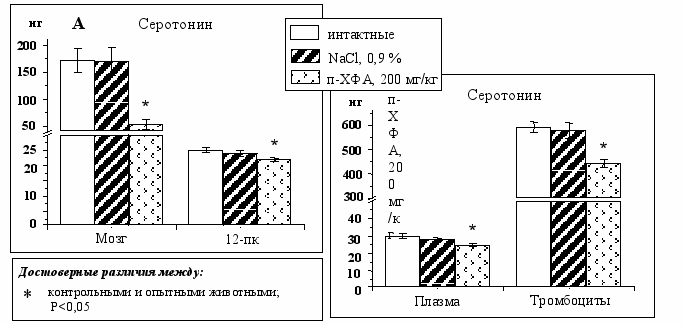
Рис. 9. Содержание серотонина у животных интактной, контрольной и опытной групп при введении п-ХФА в дозе 200 мг/кг в мозге и в двенадцатиперстной кишке (А), в крови (Б).
Итак, нейроны до формирования специфических связей функционируют как секреторные клетки, а мозг в этот период, предшествующий становлению ГЭБ, играет роль эндокринного органа. Формирование межнейрональных связей и ГЭБ приводят к качественному скачку в развитии мозга, после чего эндокринные свойства мозга начинают носить ограниченный характер. Однако по отношению к серотонину это в полной мере компенсируется периферическими источниками.
ВЫВОДЫ
- Впервые проведено сравнительное исследование развития центральной и периферической серотонин-продуцирующих систем у крыс в онтогенезе, до и после формирования ГЭБ:
а) до формирования ГЭБ (с Э21 по П4) как мозг, так и 12-пк синтезируют значительное количество серотонина, причем уровень серотонина в крови в этот период резко возрастает, достигая к П4 его значений у взрослых животных.
б) в период формирования ГЭБ (с П4 по П16) уровень серотонина в мозге резко увеличивается, в то время как в 12-пк увеличивается незначительно.
в) после формирования ГЭБ (с П16 по П30) уровень серотонина резко увеличивается в 12-пк, в то время как в мозге меняется незначительно.
2. Получены прямые доказательства поступления серотонина из мозга в общую систему циркуляции до формирования ГЭБ:
а) после декапитации плодов in utero уровень серотонина в крови плодов снизился на 40 %.
б) фармакологическое ингибирование синтеза серотонина в мозге с помощью п-ХФА привело к снижению уровня серотонина в крови на 23%
3. Полученные данные подтверждают гипотезу о том, что развивающийся мозг до формирования межнейрональных связей и гемато-энцефалического барьера функционирует как эндокринный орган, внося существенный вклад в поддержание высокой концентрации ФАВ, в частности серотонина, в общей системе циркуляции.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ.
Статьи.
1. Ugrumov M. V., Sapronova A. Y., Melnikova V., Proshlyakova E. V., Adamskaya E. I., Lavrentieva A. V., Nasirova D. I., Babichev V. N. Brain is an important source of GnRH in general circulation in the rat during prenatal and early postnatal ontogenesis // Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005. V. 141. P. 271-279.
2. Насырова Д. И., Сапронова А. Я., Нигматуллина Р. Р., Угрюмов М. В. Изменение объема плазмы крови у крыс в онтогенезе // Онтогенез. 2006. Т. 37. №4. С.1-5.
3. Насырова Д.И., Развитие центральной и периферической серотонин-продуцирующих систем у крыс в онтогенезе // Журнал эволюционной физиологии и биохимии. 2007. Т. 43. №2.
Тезисы докладов.
4. Nasyrova D. Serotonin increases stroke volume in the earl postnatal ontogenesis in rats. Neuron Differentiation and Pasticity - Regulation by Intercellular Signals. Moscow, 2003, p.56.
5. Насырова Д.И., Сапронова А.Я., Балбашев А.В., Нанаев А.К., Угрюмов М.В. Развитие центральной и периферической серотонин-продуцирующей системы у крыс в онтогенезе. I Съезд физиологов СНГ. Сочи, Дагомыс. 19-23 сентября. 2005. С. 166-167
6. Насырова Д.И., Сапронова А.Я., Уртикова Н.А., Угрюмов М.В. Становление серотонин-продуцирующей системы у крыс в онтогенезе // XIII международное совещание по эволюционной физиологии. 23-28 января 2006 года. С.-П. С. 151-152
7. Nasyrova D. I., Sapronova A Ya., Ugrumov M.V. Development of central and peripheral serotonin-producing systems in rats ontogenesis. 5 th Forum of European neuroscience, July 8-12. 2006. Vienna. P. 402
СПИСОК СОКРАЩЕНИЙ
ГЭБ – гематоэнцефалический барьер
ФАВ - физиологически активные вещества
12пк – двенадцатиперстная кишка
Э – день эмбриональный день
П – постнатальный день
п-ХФА- пара-хлорфенилаланин
5-ГТп – 5-гидрокситриптофан