
Особенности постнатального морфогенеза мышечной оболочки фундального отдела желудка белых крыс при длительном потреблении диспергированной пищи 03. 00. 25 гистология, цитология, клеточная биология
| Вид материала | Автореферат диссертации |
- Морфометрические особенности постнатального развития околоушной слюнной железы крыс, 388.58kb.
- Морфофункциональная характеристика слизистой оболочки кишечника потомства самок крыс, 303.36kb.
- Цитологические особенности вторичных миелодисплазий при лимфомах 03. 00. 25 гистология,, 526.98kb.
- Структурно-функциональная характеристика нонапептидергической гипоталамо-гипофизарной, 343.48kb.
- Формирование конечного мозга крыс после нарушения эмбрионального развития, вызванного, 370.65kb.
- Закономерности дегенерации и адаптации сетчатки глаз при экспериментальных ретинопатиях,, 745.16kb.
- Экспериментальное обоснование применения нейропептидов в комплексной терапии острого, 259.86kb.
- Морфофункциональная характеристика пульпы зуба и оценка иммунного статуса при кариесе,, 552.48kb.
- Молекулярные механизмы апоптоза при окислительном стрессе 14. 00. 16 патологическая, 606.37kb.
- Лимфоидные органы и миокард в системе мать-плод при вибрации, воздействии кадмием, 640.36kb.
СОДЕРЖАНИЕ РАБОТЫ
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом исследования послужили 204 самца беспородных белых крыс. На 21-е сутки после рождения животных произвольно разделяли на контрольную и две (I и II) опытные группы. Животных контрольной группы содержали в обычных условиях вивария на естественном для грызунов
корме, основу которого составляло цельное зерно пшеницы, а также
разрезанные на большие куски овощи. Животных I опытной группы с 21-х по 360-е сутки эксперимента кормили диспергированным кормом того же состава (измельченное на мельнице зерно, пропущенные через мясорубку овощи). Животные II опытной группы потребляли диспергированный корм до 120-х суток постнатального онтогенеза, после чего переводились до окончания эксперимента (360-е сутки онтогенеза) на естественный для
грызунов корм (корм животных контрольной группы). Кормление
осуществлялось два раза в сутки, при этом обеспечивался свободный доступ животных к корму и воде.
Для периодизации постнатального онтогенеза крыс использована
схема, предложенная В. И. Махинько и В. Н. Никитиным (1975). В возрасте 21-х (поздний молочный период), 45-ти (препубертатный период), 60-ти (препубертатный период), 120-ти (пубертатный период), 180-ти
(репродуктивный период), 270-ти (период возмужания), 360-ти (период
первой зрелости) суток животных контрольной и I опытной групп
взвешивали и декапитировали под эфирным наркозом. Декапитацию
животных II опытной группы, предварительно наркотизированных эфиром, проводили на 180-е, 270-е и 360-е сутки постнатального онтогенеза.
Объектом исследования послужил фундальный отдел желудка, участки которого фиксировали в 10% нейтральном формалине, затем обезвоживали в спиртах восходящей концентрации и заключали в парафин (Меркулов Г. А., 1969). Поперечные срезы толщиной 5-6 мкм изготавливали с помощью
санного микротома МПС-2, затем окрашивали гематоксилином и эозином (Елисеев В. Г., 1967). Окрашенные срезы заключали в бальзам.
Морфологические исследования включали: 1. Определение
абсолютной и относительной толщины продольного и циркулярного слоев, а также толщины мышечной оболочки в целом (мкм, %). 2. Определение
средних показателей площади продольного сечения ядер гладких миоцитов циркулярного и продольного слоев мышечной оболочки (мкм2), которые коррелируют с важнейшей интегративной характеристикой
морфофункционального состояния клеток – их средними объемами
(Автандилов Г. Г., 1992). 3. Определение количества ядер гладких миоцитов продольного и циркулярного слоев мышечной оболочки на стандартной площади среза (400 мкм2 при увеличении ´1000), в ходе которого ядра
гладких миоцитов подсчитывали в 50 произвольно выбранных участках гладкой мышечной ткани (%). 4. Вычисление среднего показателя ядерно-цитоплазматического отношения гладких миоцитов (%). Поскольку прямое вычисление ядерно-цитоплазматического отношения для гладких миоцитов (ЯЦОГМ) мышечной оболочки желудка оказалось невозможным из-за
нечеткости границ цитоплазмы, для его определения использовали формулу: ЯЦОГМ = (NS2/(S1 - NS2))´100%, где S1 – стандартная площадь среза (400 мкм2), S2 – средняя площадь сечения ядра гладкого миоцита, N – количество ядер гладких миоцитов на стандартной площади среза (S1). 5. Установление средней скорости (Vср.) прироста толщины продольного и циркулярного
слоев, а также мышечной оболочки в целом с помощью формулы: Vср. =
(L1-L2)/T (мкм/сутки), где L1 – средняя величина показателя в начальный
период времени, L2 – средняя величина показателя в конечный период
времени, Т - период времени.
Описание, сравнительно-морфологический анализ и морфометрия структур проводились на постоянных микропрепаратах с помощью
бинокулярного микроскопа Axiostar plus («Carl Zeiss», Германия) при
увеличениях 10´10, 10´100. Для морфометрического исследования
применялась компьютерная видеотестсистема, включающая микроскоп
«Axiostar plus», цифровую фотокамеру Canon Power Shot G 5 («Canon»,
Германия) и компьютерную программу денситофотометрии «Месоs-С 1». При определении линейных размеров проводили не менее 50 измерений структур каждого животного, среднюю площадь исследуемой структуры
определяли по итогам измерения 500 структур у каждого из изученных
животных. Все возрастные группы были представлены в исследовании не менее пятью опытными и контрольными животными. Полученные
морфометрические данные подвергали статистической обработке с
определением критерия значимости (Т) по Стьюденту, уровень значимости был принят р<0,05 (Лакин Г. Ф., 1990). Изготовлено и изучено 500
постоянных гистологических препаратов, а также 25000 микрофотографий мышечной оболочки желудка. Результаты исследования запротоколированы, а также документированы таблицами, микрофотографиями, схемами.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
1. Морфологическая характеристика постнатального развития
мышечной оболочки желудка в норме
Мышечная оболочка (МО) фундального отдела (ФО) желудка белых крыс характеризуется интенсивным (более чем трехкратным) утолщением в раннем постнатальном онтогенезе (рис. 1, 2): в период с 21-х по 180-е сутки постнатального онтогенеза ее толщина возрастает с 32,50±1,45 мкм до 104,21±1,32 мкм (р<0,05). Наиболее интенсивное разрастание гладкой
мышечной ткани МО происходит с 21-х по 60-е сутки (рис. 3), когда
толщина МО увеличивается более чем в два раза (р<0,05). Показатели
среднего прироста МО оказываются в этот период самыми высокими в
исследованном отрезке постнатального онтогенеза (р<0,05), составляя
0,47 мкм/сутки с 21-х по 45-е сутки и 1,54 мкм/сутки с 45-х по 60-е сутки (табл. 1). Со 120-х по 180-е сутки средняя скорость прироста МО снижается до 0,38 мкм/сутки (р<0,05). Рост гладкой мышечной ткани МО практически приостанавливается со 180-х по 360-е сутки постнатального онтогенеза (табл. 1), в течение которых средняя скорость прироста МО составляет
0,01-0,03 мкм/сутки, а толщина МО относительно стабилизируется (р>0,05).
На всем исследованном отрезке постнатального онтогенеза степень
развития циркулярного слоя МО желудка превышает (р<0,05) таковую
продольного слоя (табл. 2, 3). Более существенны различия между
циркулярным и продольным слоями в средней скорости прироста (рис. 4, 5), которая оказывается более высокой (р<0,05) для циркулярного слоя в раннем постнатальном онтогенезе (21-180 сутки), достигая максимума с 45-х по 60-е
сутки (1,30 мкм/сутки). Примечательно, что если в циркулярном слое
средняя скорость прироста становится минимальной со 180-х суток и
сохраняет стабильность в последующем, то в продольном слое это
происходит раньше (с 60-х суток по 360-е сутки). Более значительными (р<0,05) эти различия оказываются в позднем постнатальном онтогенезе (табл. 1, 2). Обращает внимание тот факт, что период наиболее интенсивного прироста гладкой мышечной ткани в циркулярном слое продолжается с 21-х по 180-е сутки, тогда как интенсивный прирост продольного слоя заканчивается к 60-м суткам (рис. 7, 8).
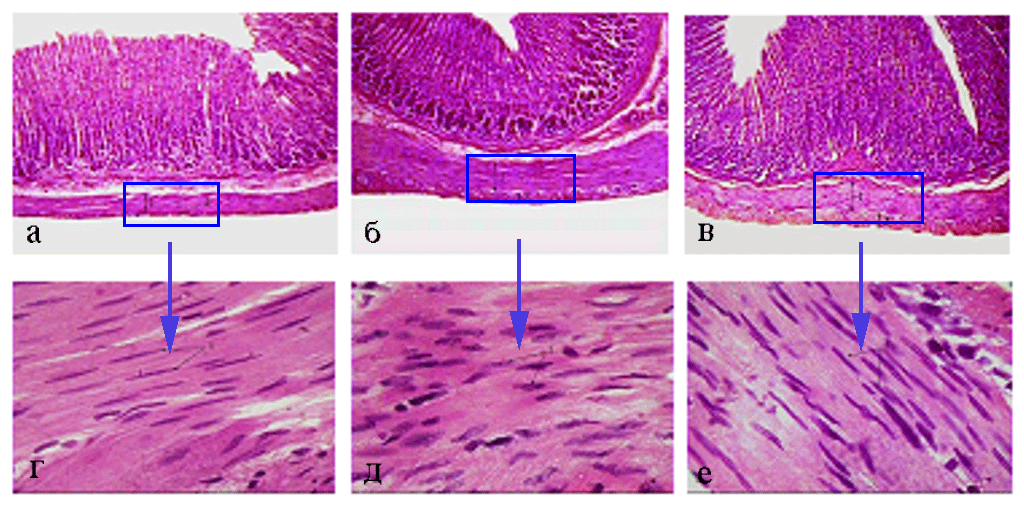
Рис. 1. Мышечная оболочка фундального отдела желудка (поперечный срез) 45- (а - 100; г - 1000), 180- (б - 100; д - 1000) и 360-суточных
(в - 100; е - 1000) животных контрольной группы. Окраска гематоксилином и эозином
В динамике значений ядерно-цитоплазматического отношения гладких миоцитов циркулярного слоя МО в постнатальном онтогенезе можно
выделить три этапа (рис. 13). На первом этапе (21-60 сутки) они достигают максимума (табл. 2), на втором этапе (120-180 сутки) значения ядерно-цитоплазматического отношения снижаются до минимума (8,46±1,13 %; 9,53±0,30 %) и на третьем (270-360 сутки) несколько возрастают (11,14±
0,43 %; 11,27±0,13 %). Следовательно, степень дифференцированности
гладких миоцитов циркулярного слоя (обратно пропорциональная значениям ядерно-цитоплазматического отношения) оказывается самой низкой (р<0,05) в период с 21-х по 60-е сутки. У 120- и 180-суточных животных этот
показатель достигает максимума (р<0,05) и несколько снижается в
последующем (270-360 сутки).
Указанные изменения степени дифференцированности гладких
миоцитов циркулярного слоя МО обусловлены увеличением площади
(и, следовательно, объема) ядер гладких миоцитов, но, в большей мере,
увеличением объема их цитоплазмы (рис. 11). Так, увеличение объема ядер (о котором судили на основании показателя их площади сечения)
происходит с 21-х по 60-е сутки (р<0,05), после чего показатель
относительно стабилизируется (табл. 2).
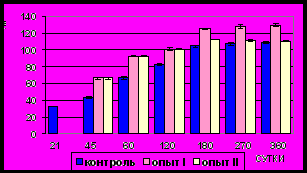
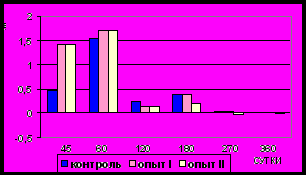
Рис. 2. Толщина (мкм) мышечной оболочки фундального отдела желудка в норме, при потреблении диспергированной пищи и в период адаптации к питанию недиспергированной пищей
Рис. 3. Средняя скорость прироста (мкм/сутки) мышечной оболочки
фундального отдела желудка в норме, при потреблении диспергированной пищи и в период адаптации к питанию недиспергированной пищей
Количество ядер гладких миоцитов циркулярного слоя на стандартной площади среза, позволяющее косвенно судить об объеме гладких миоцитов в целом, уменьшается (р<0,05) с 21-х по 180-е сутки (рис. 16), после чего
несколько возрастает (270-360 сутки). Эти особенности динамики площади сечения ядер и их плотности расположения обусловили отмеченные выше
изменения ядерно-цитоплазматического отношения гладких миоцитов
циркулярного слоя МО в постнатальном онтогенезе.
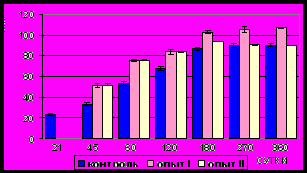
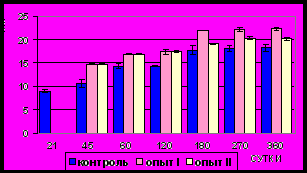
Рис. 4. Толщина (мкм) циркулярного слоя мышечной оболочки фундального отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
Рис. 5. Толщина (мкм) продольного слоя мышечной оболочки фундального отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
В динамике ядерно-цитоплазматического отношения гладких миоцитов продольного слоя МО выделяются два этапа (рис. 14, табл. 3): если на
первом этапе (21-120 сутки) значения ядерно-цитоплазматического
отношения уменьшаются (р<0,05), то на втором этапе (180-360 сутки) они
увеличиваются (р<0,05). В промежутке между этими этапами (120-180
сутки) степень дифференцированности гладких миоцитов оказывается
наиболее высокой (р<0,05). Такой характер изменений ядерно-цитоплазматического отношения обусловлен, с одной стороны, увеличением площади сечения (а, следовательно, и объема) ядер гладких миоцитов (р<0,05) в период с 45-х по 180-е сутки, а с другой, уменьшением (р<0,05) плотности их расположения (измеряемой как количество ядер на
стандартной площади среза) с 45-х по 120-е сутки, свидетельствующей об увеличении объема цитоплазмы гладких миоцитов (рис. 14, 17).
Таким образом, циркулярный и продольный слои МО ФО желудка
различаются динамикой ядерно-цитоплазматического отношения гладких миоцитов, указывающей на специфику морфогенетических процессов,
протекающих в каждом из слоев МО в норме. При этом в каждом слое имеет место чередование периодов интенсивного роста и дифференцировки
гладкой мышечной ткани, обусловливающее особенности постнатального изменения толщины циркулярного и продольного слоев МО в целом.
Интенсивное утолщение последних происходит в период с 21-х по 180-е
сутки, по завершению которого наступает относительная стабилизация их степени развития.
2. Особенности постнатального морфогенеза мышечной оболочки
желудка при питании диспергированной пищей
Интенсивное развитие МО ФО желудка животных, питающихся
диспергированной пищей, продолжается до 180-х суток (табл. 1). В этот
период происходит утолщение как циркулярного, так и продольного слоев (р<0,05). В последующем (180-360 сутки) толщина как МО в целом, так и каждого из ее слоев относительно стабилизируется (табл. 2, 3). Такой
динамике толщины МО соответствуют изменения средней скорости
прироста МО (табл. 1), достигающей максимума (р<0,05) в период с 21-х по 45-е сутки (1,42 мкм/сутки) и с 45-х по 60-е сутки (1,71 мкм/сутки).
Минимальные значения средней скорости прироста МО (р<0,05)
характеризуют период со 180-х по 270-е сутки (0,03 мкм/сутки) и с 270-х по 360-е сутки (0,02 мкм/сутки). Примечателен тот факт, что в раннем
постнатальном онтогенезе (21-180 сутки) средняя скорость прироста
циркулярного слоя в 6-13 раз превышает (р<0,05) таковую продольного слоя (рис. 7, 8). В последующем (180-360 сутки) эти различия исчезают (р>0,05). Как следствие этого циркулярный слой претерпевает в исследованный
период (21-360 сутки) почти пятикратное утолщение (рис. 2), в то время как толщина продольного слоя возрастает в 2,5 раза (рис. 3).
Ядерно-цитоплазматическое отношение гладких миоцитов
циркулярного слоя МО характеризуется на большем отрезке исследованного периода постнатального онтогенеза относительной стабильностью (рис. 13). Возрастание его значений (р<0,05) наблюдается с 21-х по 45-е сутки и со
120-х по 180-е сутки (табл. 2). В период со 180-х по 270-е сутки ядерно-цитоплазматическое отношение существенно снижается (р<0,05),
стабилизируясь в последующем (рис. 6). Такая динамика ядерно-цитоплазматического отношения обусловлена, с одной стороны,
значительным увеличением площади сечения (а, следовательно, и объема) ядер гладких миоцитов (р<0,05), а с другой стороны – постепенным
уменьшением объема цитоплазмы, подтверждаемого уменьшением
плотности расположения ядер гладких миоцитов (р<0,05) на стандартной площади среза (табл. 2).
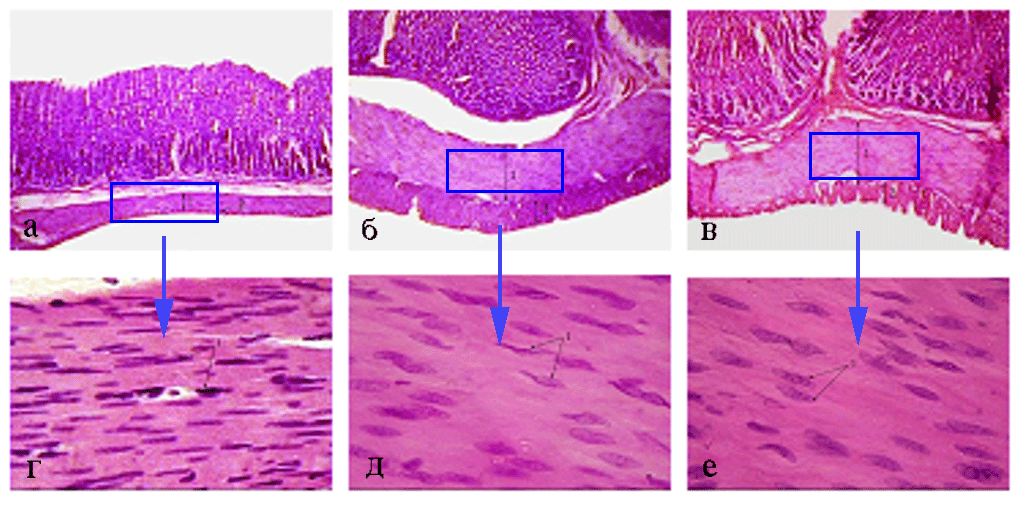
Рис. 6. Мышечная оболочка фундального отдела желудка (поперечный срез) 45- (а - 100; г - 1000), 180- (б - 100; д - 1000) и 360-суточных
(в - 100; е - 1000) животных I опытной группы. Окраска гематоксилином и эозином
Более сложные четырехэтапные изменения ядерно-цитоплазматичекого отношения отличают продольный слой МО (табл. 3). Два периода
увеличения ядерно-цитоплазматического отношения (с 21-х по 45-е сутки и с 60-х по 180-е сутки) последовательно сменяются двумя периодами снижения рассматриваемого показателя: с 45-х по 60-е сутки и со 180-х по 270-е сутки (рис. 14). Эти четырехэтапные изменения ядерно-цитоплазматического
отношения гладких миоцитов продольного слоя обусловлены более сложной динамикой изменения объема их ядер и цитоплазмы, о которых судили на основании площади сечения ядер и их количества на стандартной площади среза (табл. 3). Так, увеличение площади сечения ядер гладких миоцитов с 21-х по 60-е сутки сменяется периодом относительной стабилизации
показателя (60-120 сутки), после которого наступают периоды его роста (120-180 сутки) и относительной стабилизации (180-360 сутки). Динамике количества ядер гладких миоцитов на стандартной площади среза
продольного слоя свойственны этапы снижения показателя (21-60 сутки), относительной стабилизации (60-180 сутки), повторного снижения
(180-270 сутки) и повторной относительной стабилизации (270-360 сутки).
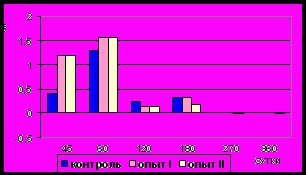
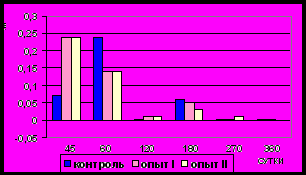
Рис. 7. Средняя скорость прироста (мкм/сутки) циркулярного слоя
мышечной оболочки фундального
отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
Рис. 8. Средняя скорость прироста (мкм/сутки) продольного слоя
мышечной оболочки фундального
отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
3. Сравнительная характеристика постнатального морфогенеза
мышечной оболочки желудка при питании диспергированной и
недиспергированной пищей
На всем исследованном отрезке онтогенеза толщина МО животных,
питающихся диспергированной пищей, превышает (р<0,05) таковую
животных, питающихся недиспергированной пищей. Следовательно,
питание диспергированной пищей обусловливает устойчивую гипертрофию МО ФО желудка белых крыс (рис. 9, 10). Уже в первом исследованном
периоде (21-45 сутки) МО опытных животных утолщается более чем в 2 раза и достигает 66,60±0,52 мкм (р<0,05), в то время как ее толщина у
45-суточных контрольных животных равна 43,83±1,05 мкм (табл. 1).
Значительное утолщение за сравнительно короткий период времени МО претерпевает с 45-х по 60-е сутки, увеличиваясь с 66,60±0,52 мкм до 92,23±1,37 мкм (р<0,05).
Различия средней скорости прироста МО у опытных и контрольных животных (табл. 1) обеспечивают появление максимальных различий в
толщине МО между опытными и контрольными животными в возрасте 45-и и 60-и суток (р<0,05). У 120-, 180-, 270- и 360-суточных животных они
снижаются до разницы в толщине равной примерно 20 мкм (р<0,05).
Показатели средней скорости прироста МО в эти периоды обретают
сходство у животных опытной и контрольной групп (табл. 1).
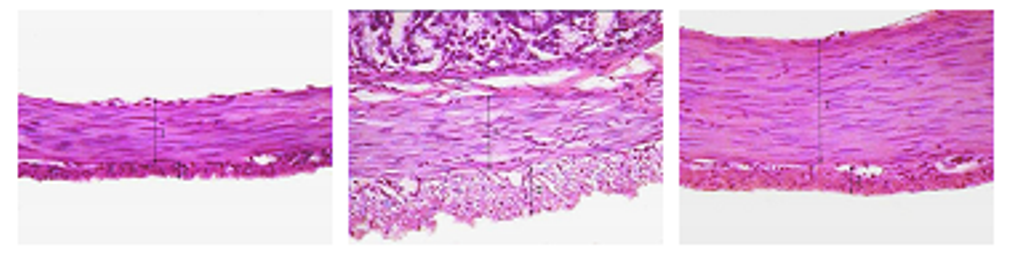
Рис. 9. Мышечная оболочка фундального отдела желудка (поперечный срез) 45- (а), 180- (б) и 360-суточных (в) животных контрольной группы.
Окраска гематоксилином и эозином, ×400
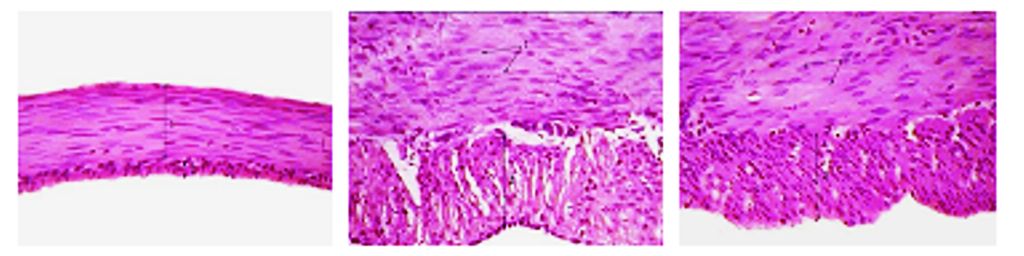
Рис. 10. Мышечная оболочка фундального отдела желудка
(поперечный срез) 45- (а), 180- (б) и 360-суточных (в) животных I опытной группы. Окраска гематоксилином и эозином, ×400
Динамика различий толщины циркулярного слоя между животными
I опытной и контрольной групп соответствует таковой общей толщины МО (табл. 1, 2). Максимальные различия (р<0,05) толщины циркулярного слоя МО между животными, питающимися диспергированной и
недиспергированной пищей, наблюдаются в возрасте 45-и и, особенно, 60-и суток. В последующем (120-360 сутки) они несколько уменьшаются,
сохраняя, однако, достоверный характер (рис. 4).
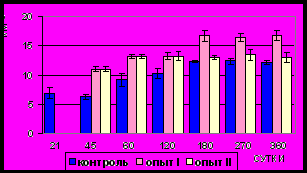
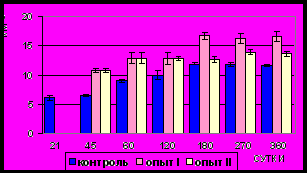
Рис. 11. Площадь сечения ядер
гладких миоцитов (мкм2) циркулярного слоя мышечной оболочки фундального отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
Рис. 12. Площадь сечения ядер
гладких миоцитов (мкм2) продольного слоя мышечной оболочки фундального отдела желудка в норме, при
потреблении диспергированной пищи и в период адаптации к питанию
недиспергированной пищей
Питание диспергированной пищей обусловливает также гипертрофию продольного слоя МО. Наименьшие различия в толщине продольного слоя свойственны 60- и 120-суточным животным (рис. 5). Максимальными
(с учетом абсолютных значений толщины продольного слоя) они
оказываются у 45-суточных животных (табл. 3).
Таким образом, гипертрофия МО ФО желудка белых крыс, питающихся диспергированной пищей, обеспечивается разрастанием гладкой мышечной ткани как в циркулярном, так и в продольном слоях. Однако с учетом
соотношения толщины продольного и циркулярного слоев следует отметить, что общая гипертрофия МО животных, питающихся диспергированной
пищей, обеспечивается в основном утолщением ее циркулярного слоя
(рис. 7, 8).
Степень дифференцированности гладких миоцитов циркулярного слоя МО обнаруживает сходство у 45-, 270- и 360-суточных животных I опытной и контрольной групп (табл. 2, 3). У 120- и 180-суточных животных,
питающихся диспергированной пищей она резко снижается по отношению к контрольным животным соответствующих возрастных групп (р<0,05).
Небольшое повышение (р<0,05) степени дифференцированности гладких миоцитов циркулярного слоя по отношению к контрольным животным
наблюдается у 60-суточных животных I опытной группы (рис. 11, 13). Эти изменения рассматриваемого показателя циркулярного слоя позволяют
констатировать, что в период с 60-х по 180-е сутки у животных, питающихся диспергированной пищей, в циркулярном слое МО активизируются
процессы биосинтеза и роста. Подтверждением этому может
рассматриваться значительное увеличение (р<0,05) площади сечения
(объема) ядер гладких миоцитов у животных I опытной группы в сравнении с соответствующим показателем для животных контрольной группы
(табл. 2). Отдельного рассмотрения заслуживают результаты сравнения двух показателей площади сечения ядер гладких миоцитов и количества ядер гладких миоцитов циркулярного слоя на стандартной площади среза у 270- и 360-суточных животных I опытной и контрольной групп: если первый
показатель у опытных животных возрастает по отношению к контрольным животным (р<0,05), то второй достоверно снижается (табл. 2). Это
свидетельствует о сохранении гладкими миоцитами циркулярного слоя
гипертрофированного состояния по завершении, в основном, процессов
роста и дифференцировки гладкой мышечной ткани.
Различия в динамике ядерно-цитоплазматического отношения гладких миоцитов продольного слоя между животными I опытной и контрольной групп соответствуют таковым циркулярного слоя МО с тем отличием, что у 45-суточных опытных животных показатель ядерно-цитоплазматического отношения оказывается выше (р<0,05), чем у контрольных животных этого возраста (рис. 14). Последнее обусловлено существенной гипертрофией гладких миоцитов у 45-суточных животных, питающихся диспергированной пищей, подтверждаемой как значительным увеличением (р<0,05) у
45-суточных животных I опытной группы площади сечения ядер, так уменьшением (р<0,05) плотности расположения ядер гладких миоцитов,
указывающим на увеличение объема цитоплазмы миоцитов продольного слоя (табл. 3).
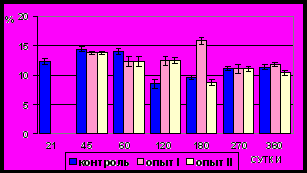
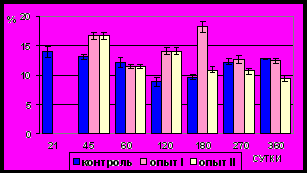
Рис. 13. Ядерно-цитоплазматическое отношение гладких миоцитов (%)
циркулярного слоя мышечной оболочки фундального отдела желудка в норме, при потреблении диспергированной пищи и в период адаптации к питанию недиспергированной пищей
Рис. 14. Ядерно-цитоплазматическое отношение гладких миоцитов (%)
продольного слоя мышечной оболочки фундального отдела желудка в норме, при потреблении диспергированной пищи и в период адаптации к питанию недиспергированной пищей
Подводя итог рассмотрению результатов сравнительной оценки
особенностей постнатального морфогенеза МО ФО желудка белых крыс можно заключить, что питание диспергированной пищей обуславливает
устойчивую гипертрофию МО, проявляющуюся уже на 21-е сутки после
начала эксперимента и сохраняющуюся в течение всего исследованного
периода постнатального онтогенеза (45-360 сутки). Гипертрофия МО
желудка белых крыс, питающихся диспергированной пищей
обусловливается интенсивным разрастанием гладкой мышечной ткани как в циркулярном, так и в продольном слоях. В основе гипертрофии
циркулярного и продольного слоев МО лежит гипертрофия их гладких
миоцитов, подтверждаемая увеличением (р<0,05) площади сечения
(а, следовательно, и объема) ядер гладких миоцитов и уменьшением (р<0,05) плотности расположения ядер на стандартной площади среза,
свидетельствующим об увеличении объема цитоплазмы гладких миоцитов (табл. 2, 3). Характер изменений значений ядерно-цитоплазматического
отношения гладких миоцитов как циркулярного, так и продольного слоев (рис. 13, 14), возрастание плотности расположения клеточных ядер в толще среза гладкой мышечной ткани у 120- и 180-суточных животных,
питающихся диспергированной пищей, дает основания предположить
возможность пролиферации гладких миоцитов в обоих слоях МО (табл. 2, 3). Характер изменений ядерно-цитоплазматического отношения гладких
миоцитов указывает на специфику морфологической реакции каждого из слоев МО. В циркулярном слое показатель ядерно-цитоплазматического
отношения возрастает (р<0,05) в течение одного периода (60-180 сутки). В продольном слое его рост наблюдается в течение двух периодов: с 21-х по 45-е сутки и с 60-х по 180-е сутки (р<0,05).
Таким образом, если первый биологический механизм гипертрофии МО ФО желудка белых крыс при питании диспергированной пищей –
гипертрофия гладких миоцитов - проявляется на протяжении всего
исследованного периода онтогенеза (21-360 сутки), то реализация второго механизма - пролиферации гладких миоцитов - ограничивается в
циркулярном слое одним (60-180 сутки), а в продольном слое – двумя
(21-45 сутки и 60-180 сутки) временными отрезками.