
Морфометрические особенности постнатального развития околоушной слюнной железы крыс в условиях питания диспергированной пищей 03. 03. 04 клеточная биология, цитология, гистология
| Вид материала | Автореферат диссертации |
- Особенности постнатального морфогенеза мышечной оболочки фундального отдела желудка, 394.65kb.
- Структурно-функциональная характеристика нонапептидергической гипоталамо-гипофизарной, 343.48kb.
- Формирование конечного мозга крыс после нарушения эмбрионального развития, вызванного, 370.65kb.
- Морфофункциональная характеристика слизистой оболочки кишечника потомства самок крыс, 303.36kb.
- Цитологические особенности вторичных миелодисплазий при лимфомах 03. 00. 25 гистология,, 526.98kb.
- Лимфоидные органы и миокард в системе мать-плод при вибрации, воздействии кадмием, 640.36kb.
- Онтогенетические особенности морфофизиологического состояния продуктивных животных, 585.31kb.
- Ультраструктурная и цитохимическая характеристика макрофагов, инфицированных рнк-содержащими, 636.4kb.
- Закономерности дегенерации и адаптации сетчатки глаз при экспериментальных ретинопатиях,, 745.16kb.
- Влияние биологически активных клеточных компонентов растений на структурные изменения, 324.85kb.
Особенности постнатального морфогенеза ацинусов околоушной
слюнной железы белых крыс при питании диспергированной
пищей
Препубертатный период характеризуется уменьшением площади сечения и диаметра ацинусов животных I опытной группы, что обусловлено статистически значимым снижением площади ядер и цитоплазмы сероцитов.
Таблица 2
Морфометрические показатели ацинусов околоушной слюнной железы контрольных и опытных животных в разные периоды постнатального развития
| Показатель Возраст (сутки), группа | Площадь ядер сероцитов (мкм2) | Диаметр ядер сероцитов (мкм) | Площадь сечения цитоплазмы сероцитов (мкм2) | ЯЦО (%) сероцитов | Количество сероцитов на срезе ацинуса | Площадь сечения ацинусов (мкм2) | Диаметр ацинусов (мкм) | |
| 21 | 9,06±0,06 | 3,39±0,01 | 56,81±0,73 | 17,53±0,18 | 4,01±0,04 | 263,78±3,42 | 16,92±0,12 | |
| 60 | Контроль | 9,77±0,05X | 3,52±0,01X | 51,24±0,55X | 19,49±0,23X | 4,19±0,05 | 254,88±3,72 | 16,41±0,13X |
| Опыт I | 9,14±0,06* | 3,41±0,01* | 53,72±0,78X | 17,09±0,22* | 3,96±0,05* | 243,94±3,43X | 16,24±0,12X | |
| 120 | Контроль | 8,53±0,05X | 3,29±0,01X | 59,00±0,73X | 14,55±0,17X | 4,31±0,04 | 294,61±4,46X | 17,97±0,14X |
| Опыт I | 9,08±0,06* | 3,40±0,01* | 60,78±0,86X | 15,23±0,17X* | 4,05±0,04* | 279,15±3,36X | 17,52±0,11X | |
| 180 | Контроль | 9,07±0,05X | 3,40±0,01X | 65,08±0,88X | 14,51±0,19 | 4,38±0,05 | 326,34±4,15X | 18,93±0,12X |
| Опыт I | 8,88±0,05 | 3,36±0,01 | 56,77±0,81X* | 16,27±0,23X* | 4,54±0,05X | 307,08±4,62X* | 18,35±0,15X* | |
| Опыт II | 9,63±0,06*V | 5,01±0,01*V | 72,15±1,05*V | 14,57±0,21V | 4,35±0,05 | 352,94±4,95*V | 19,73±0,14*V | |
| 240 | Контроль | 10,76±0,05X | 3,67±0,01X | 77,27±0,95X | 14,28±0,17 | 4,82±0,05X | 421,40±4,88X | 21,59±0,13X |
| Опыт I | 8,94±0,05* | 3,38±0,01* | 77,59±0,84X | 11,65±0,16X* | 4,64±0,05 | 419,94±5,76X | 21,71±0,15X | |
| Опыт II | 9,92±0,05X*V | 3,56±0,01X*V | 74,15±0,71V | 13,62±0,14X*V | 4,77±0,05X | 409,73±5,44X | 21,15±0,13X | |
Примечание.
X - статистически значимые отличия от предыдущего возраста (р<0,01);
* - статистически значимые отличия от контрольных значений (р<0,01);
V - статистически значимые отличия от значений животных I опытной группы (р<0,01).
Пубертатный период, наоборот, характеризуется увеличением площади сечения ацинусов и повышением функциональной активности сероцитов, на которую указывает снижение их ЯЦО (табл. 2). В репродуктивный период происходит увеличение площади сечения ацинусов, в основе которого лежит существенное увеличение количества сероцитов на единицу площади сечения ацинусов. При этом площадь цитоплазмы сероцитов и их ЯЦО уменьшаются. Указанные показатели у 180-суточных животных составляют 56,77±0,81 мкм2 и 16,27±0,23 %. В период возмужания отмечается рост площади сечения ацинусов до 419,94±5,76 мкм2 за счет увеличения площади цитоплазмы сероцитов. При этом, судя по уменьшению значений ЯЦО, функциональная активность сероцитов повышается.
В период с 21-х по 120-е сутки отмечается уменьшение количества сероцитов на площади сечения ацинусов у животных I опытной группы по отношению к контрольным животным. Судя по ЯЦО, функциональная активность сероцитов у животных I опытной группы в препубертатный период выше, чем у животных контрольной группы. Наблюдающееся снижение размеров ацинусов 180-суточных животных I опытной группы обусловливается соответствующими изменениями площади цитоплазмы сероцитов
(рис. 1). Значения ЯЦО сероцитов 120- и 180-суточных животных I опытной группы превышают аналогичные показатели контрольных животных соответствующего возраста. Уменьшение размеров ацинусов околоушной слюнной железы обусловлено питанием предварительно измельченной пищей и, как следствие, сокращением продолжительности акта жевания. Раздражение рецепторов ротовой полости во время жевания является главным механизмом в регулировании синтеза белков слюны (Anderson D.J., Hector M.P., 1987; Ikawa M. et al., 1991; Johnson D.A., Cardenas H.L., 1993). Время нахождения в ротовой полости предварительно измельченной пищи сокращается, что ведет к угнетению физиологического рефлекторного процесса синтеза секрета околоушной слюнной железой (Mackie D.A., Pangborn R.M., 1990; C.-K. Yeh et al., 1990). Как следствие, наблюдается снижение объема ацинусов, которое обусловлено уменьшением объема сероцитов (Wilborn W.H., Schneyer C.A., 1970; Sreebny L.M. et al., 1971; Johnson D.A., Cardenas H.L., 1993; Scott J., Gunn D.L., 1994; Leal S.C. et al., 2003).
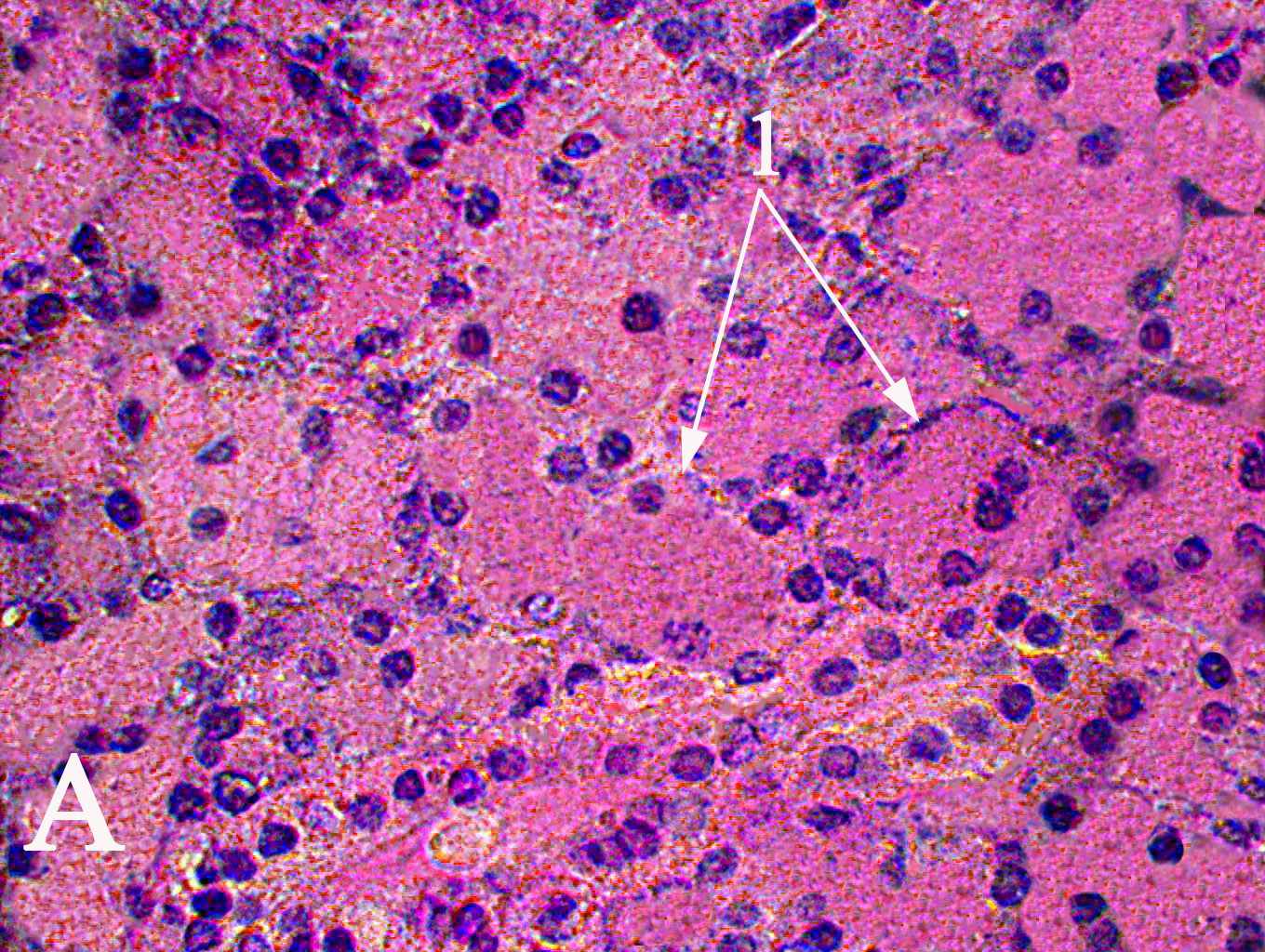
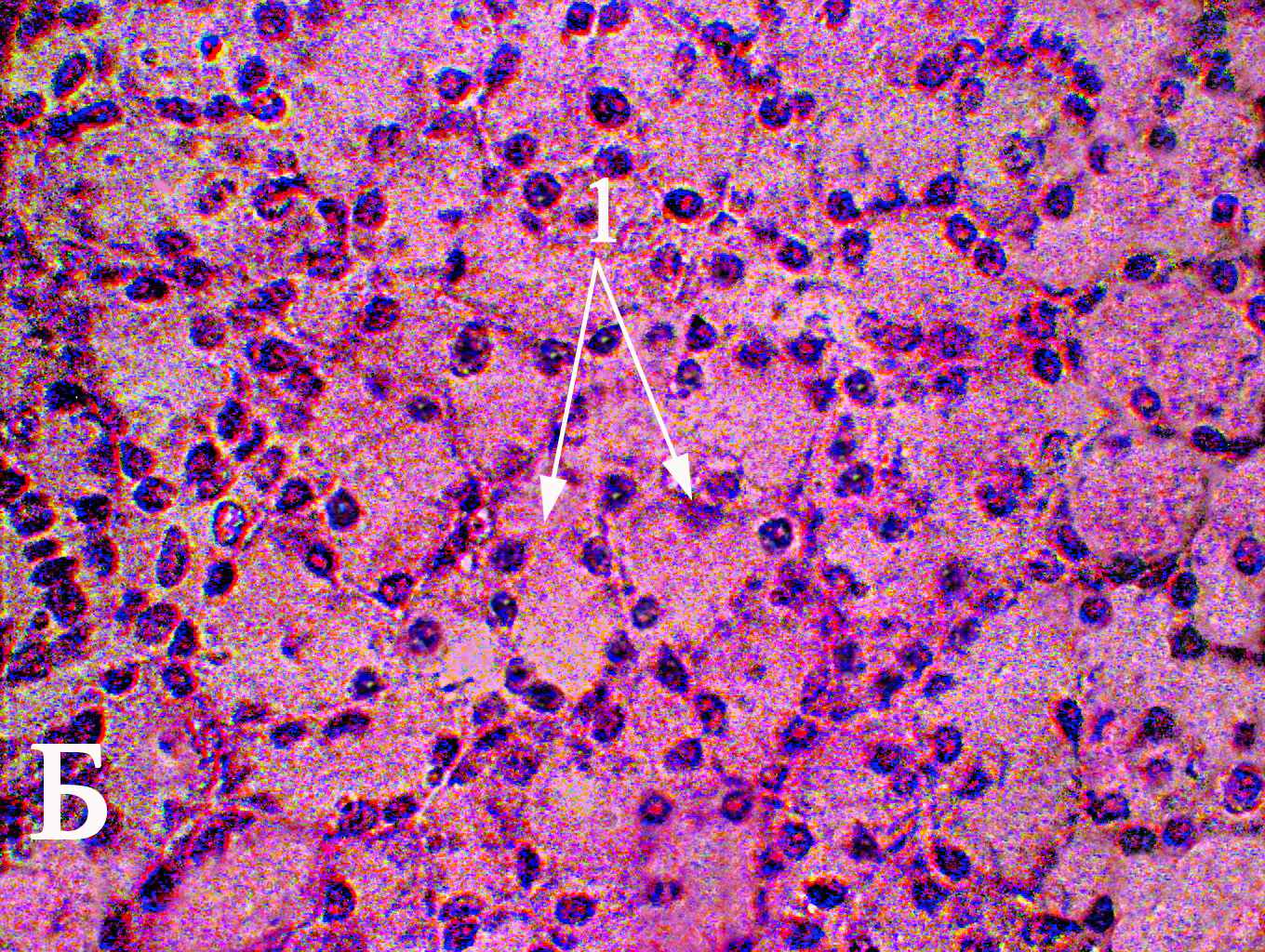
Рис. 1. Ацинусы (1) околоушной слюнной 180-суточных животных контрольной группы (А) и I опытной группы (Б). Окраска гематоксилин-эозином, увеличение ×1000.
К 240-м суткам площадь сечения ацинусов и цитоплазмы сероцитов, а также их количество на площади сечения ацинусов животных I опытной группы достигают значений контрольных животных. При этом у 240-суточных животных I опытной группы ЯЦО сероцитов оказывается ниже, чем у 240-суточных контрольных животных. Увеличение площади сечения ацинусов 240-суточных животных I опытной группы, вероятно, связано с потреблением большего количества пищи. В ходе исследования установлено, что к 240-м суткам животные I опытной группы потребляли пищи в 2,3 раза больше, чем животные контрольной группы (p<0,001). Последнее мы склонны связывать с более поздним наступлением преабсорбтивного (сенсорного) насыщения вследствие более позднего наступления рефлекторных раздражений от механорецепторов слизистой оболочки полости рта, обусловленных слабым раздражающим действием на них диспергированной пищи.
Таким образом, длительное питание диспергированной пищей вызывает изменения ацинусов околоушной слюнной железы беспородных белых крыс, которые в целом можно охарактеризовать как гипо- и атрофические. К концу периоду возмужания происходит увеличение морфометрических показателей ацинусов животных I опытной группы до значений контрольных животных.
Особенности постнатального морфогенеза внутридольковых
протоков околоушной слюнной железы белых крыс при питании диспергированной пищей
Период с 21-х по 180-е сутки характеризуется стабильными значениями площади сечения стенок и просветов вставочных протоков. При этом ЯЦО эпителиоцитов протоков, составляя у 21-суточных животных 31,84±0,61 %, существенно не изменяется у 60-суточных животных. К 120-м суткам ЯЦО уменьшается, затем этот показатель стабилизируется до 240-х суток. Период возмужания характеризуется увеличением площади сечения стенок и просветов вставочных протоков, что происходит за счет увеличения количества эпителиоцитов в стенках протоков (табл. 3 и 4).
Препубертатный период не сопровождается изменениями площади сечения стенок и диаметра исчерченных протоков, в то время как площадь сечения цитоплазмы эпителиоцитов уменьшается с 40,18±1,07 мкм2 у 21-суточных животных до 33,64±0,79 мкм2 у 60-суточных животных (р<0,001). При этом отмечается также увеличение ЯЦО эпителиоцитов с 29,12±0,62 % до 39,48±0,90 %. В пубертатный период площадь сечения и диаметр исчерченных протоков уменьшаются, однако площадь цитоплазмы эпителиоцитов и их количество на площади поперечного сечения исчерченных протоков сохраняют стабильные значения. ЯЦО эпителиоцитов уменьшается до
27,46± 0,57 %, не изменяясь до конца исследованного периода онтогенеза.
В репродуктивный период площадь сечения стенок и диаметр исчерченных протоков не изменяются на фоне увеличения количества эпителиоцитов в стенках исчерченных протоков. Площадь сечения и диаметр просветов, сохраняющие стабильные значения до 120-х суток, в репродуктивный период уменьшаются. Период возмужания характеризуется увеличением площади сечения стенок и просветов исчерченных протоков, что обусловлено увеличением площади цитоплазмы эпителиоцитов.
Период с 21-х по 180-е сутки характеризуется отсутствием различий в площади сечения стенок и диаметра внутридольковых протоков между животными контрольной и I опытной группы. Однако внутридольковые протоки 60-суточных животных I опытной группы имеют меньшую площадь цитоплазмы эпителиоцитов по сравнению с таковой 60-суточных контрольных животных (р<0,01). Репродуктивный период отличается большими значениями площади сечения и диаметра просветов внутридольковых протоков контрольных животных, чем соответствующие показатели животных I опытной группы. В период возмужания площадь сечения и диаметр просветов внутридольковых протоков животных, питающихся диспергированной пищей, увеличиваются, достигая значений соответствующих показателей животных контрольной группы, в то время как площадь сечения стенок внутридольковых протоков контрольных животных оказывается большей, чем у животных I опытной группы (р<0,01). Площадь сечения стенок внутридольковых протоков 240-суточных животных I опытной группы соответствует значениям, свойственным 21-суточным животным. В период с 21-х по 180-е сутки ЯЦО эпителиоцитов вставочных протоков животных контрольной и I опытной группы имеет сходные значения. Однако ЯЦО эпителиоцитов исчерченных протоков 60-суточных животных I опытной группы превышает таковое животных контрольной группы (р<0,01). В период возмужания ЯЦО эпителиоцитов внутридольковых протоков контрольных животных оказываются ниже, чем у животных I опытной группы (табл. 3).
Таблица 3
Морфометрические показатели внутридольковых протоков околоушной слюнной железы контрольных и опытных животных в разные периоды постнатального развития
| Показатель | 21 | Контроль-60 | Опыт I-60 | Контроль -120 | Опыт I-120 | Контроль -180 | Опыт I-180 | Опыт II-180 | Контроль -240 | Опыт I-240 | Опыт II-240 | |
| S ПР | ВП | 13,70±0,51 | 13,01±0,50 | 12,43±0,80 | 15,40±0,47X | 14,02±0,79 | 21,16±0,96X | 14,48±0,84* | 23,82±1,15 | 26,29±0,89X | 28,24±1,40X | 21,79±0,73*V |
| ИП | 34,69±1,46 | 35,96±1,63 | 39,53±1,84 | 43,34±1,63X | 43,36±1,70 | 49,14±1,51X | 35,87±1,23X* | 46,88±1,23 | 56,82±1,57X | 60,49±1,94X | 63,76±2,13X | |
| D ПР | ВП | 4,16±0,08 | 4,01±0,08 | 3,91±0,12 | 4,38±0,07X | 4,13±0,12 | 5,10±0,12X | 4,20±0,12* | 5,37±0,14V | 5,72±0,10X | 5,83±0,14X | 5,14±0,08*V |
| ИП | 6,75±0,15 | 6,62±0,14 | 6,98±0,16 | 7,19±0,12X | 7,29±0,14 | 8,09±0,15X | 6,76±0,12X* | 8,45±0,15V | 9,23±0,19X | 9,28±0,19X | 8,82±0,14 | |
| КЭСП | ВП | 6,26±0,08 | 6,28±0,09 | 6,42±0,14 | 6,62±0,06X | 6,29±0,09* | 6,57±0,09 | 6,47±0,10 | 6,82±0,08V | 6,91±0,06X | 6,94±0,09X | 7,01±0,01X |
| ИП | 9,14±0,10 | 8,99±0,10 | 9,03±0,12 | 9,01±0,08 | 8,79±0,08 | 9,28±0,10 | 9,22±0,10Х | 9,38±0,09 | 9,83±0,10Х | 9,57±0,10 | 9,47±0,10 | |
| S ЯЭ | ВП | 11,71±0,11 | 9,02±0,09Х | 8,77±0,14X | 8,15±0,07X | 8,99±0,11* | 9,43±0,08X | 9,15±0,12 | 10,10±0,11*V | 10,21±0,08X | 9,70±0,10X* | 9,84±0,07* |
| ИП | 11,13±0,09 | 9,55±0,08X | 9,47±0,10X | 8,25±0,05X | 8,92±0,07X* | 8,87±0,06X | 9,58±0,07X* | 9,85±0,05*V | 10,16±0,06X | 9,96±0,06X | 9,81±0,05* | |
| D ЯЭ | ВП | 3,85±0,02 | 3,38±0,02Х | 3,32±0,03Х | 3,21±0,01Х | 3,37±0,02* | 3,49±0,02 Х | 3,41±0,02* | 3,58±0,02*V | 3,61±0,01Х | 3,51±0,02Х* | 3,54±0,01* |
| ИП | 3,76±0,02 | 3,48±0,01Х | 3,47±0,02Х | 3,24±0,01Х | 3,34±0,01Х* | 3,35±0,01Х | 3,36±0,01 | 3,52±0,01*V | 3,62±0,01Х | 3,62±0,01Х | 3,64±0,01Х | |
| S ЦЭ | ВП | 36,42±0,81 | 37,83±0,88 | 33,64±0,78* | 31,04±0,62X | 33,68±0,87 | 34,24±1,02 | 32,85±0,95 | 34,10±0,82 | 44,32±1,02X | 36,28±0,95* | 37,67±0,86* |
| ИП | 40,18±1,07 | 37,82±0,69 | 33,64±0,79X* | 31,08±0,56X | 31,82±0,50 | 33,12±0,62 | 33,21±0,58 | 35,19±0,50*V | 45,88±0,87X | 38,65±0,64X* | 38,79±0,75X* | |
| ЯЦО Э | ВП | 31,84±0,61 | 30,32±0,57 | 33,15±1,18 | 27,24±0,58Х | 27,31±0,60Х | 28,16±0,54 | 27,93±0,74 | 30,11±0,74 | 23,83±0,22Х | 27,98±0,60* | 27,82±0,54* |
| ИП | 29,12±0,62 | 36,05±0,59Х | 39,48±0,90Х* | 27,62±0,46Х | 27,46±0,57Х | 28,45±0,49 | 29,35±0,59 | 29,27±0,52 | 23,43±0,39Х | 27,94±0,45* | 25,99±0,38Х*V | |
Приведенные результаты в целом позволяют констатировать наличие отклонений в морфогенезе ацинусов околоушной слюнной железы белых крыс в период длительного питания диспергированной пищей, которые к концу исследуемого периода онтогенеза частично восстанавливаются. Значения же площади сечения стенок внутридольковых выводных протоков животных, питающихся диспергированной пищей, к концу исследованного периода онтогенеза достигают лишь значений 21-суточных животных (табл. 4).
Таблица 4
Морфометрические показатели внутридольковых протоков
околоушной слюнной железы белых крыс животных контрольной и опытных групп в разные периоды развития
| Возраст (сутки) | Показатель Группа | Площадь сечения (мкм2) стенок внутридольковых протоков | Диаметр (мкм) внутридольковых протоков | ||
| ВП | ИП | ВП | ИП | ||
| 21 | Контроль | 310,13±7,32 | 467,58±12,40 | 17,19±0,23 | 21,47±0,31 |
| 60 | Контроль | 306,05±7,05 | 470,38±9,72 | 17,21±0,21 | 20,90±0,22 |
| Опыт I | 288,34±11,1 | 457,59±14,43 | 16,52±0,37 | 20,33±0,34 | |
| 120 | Контроль | 261,19±4,37Х | 361,29±7,25Х | 16,06±0,14Х | 18,92±0,19Х |
| Опыт I | 270,03±7,93 | 369,09±7,30Х | 16,18±0,23 | 19,12±0,19Х | |
| 180 | Контроль | 284,75±4,63Х | 378,50±6,22 | 16,79±0,21Х | 19,60±0,19Х |
| Опыт I | 277,00±7,53 | 393,13±7,31 | 16,51±0,25 | 19,89±0,21 | |
| Опыт II | 306,94±7,17V | 407,02±6,04* | 17,28±0,22 | 20,37±0,18* | |
| 240 | Контроль | 376,40±7,87Х | 523,96±9,61Х | 19,61±0,21Х | 23,32±0,23Х |
| Опыт I | 311,45±7,04Х* | 469,14±8,63Х* | 17,26±0,18* | 21,51±0,20Х* | |
| Опыт II | 323,85±6,05* | 461,37±8,59Х* | 17,92±0,19* | 21,42±0,21Х* | |
Примечание. X - статистически значимые отличия от предыдущего возраста (р<0,01); * - статистически значимые отличия от контрольных значений (р<0,01); V - статистически значимые отличия от значений животных I опытной группы (р<0,01).