
Особенности эволюции морфологических и молекулярных признаков на примере близкородственных видов дрозофил 03. 02. 07 генетика
| Вид материала | Автореферат |
- Молекулярные механизмы формирования множественной лекарственной резистентности у Burkholderia, 787.44kb.
- Рабочая программа генетика Код дисциплины по учебному плану опд ф. 6 для студентов, 274.5kb.
- Наследственность и ее роль в патологии медицинская генетика и ее задачи Наследственность, 327kb.
- Эволюция и Генетика, 167.01kb.
- Рабочая программа генетика и селекция Код дисциплины по учебному плану опд ф 1 для, 292.62kb.
- Практическая работа «Генетика», 116.83kb.
- Л. П. Татаринов необратимость эволюции и ее направленность, 194.79kb.
- Генетика Генетика, 18.18kb.
- Л. И. Параллелизмы в молекулярной организации генома и проблемы эволюции. В кн.: Молекулярные, 251.18kb.
- Рабочая программа дисциплины «молекулярная генетика» Код дисциплины по учебному плану, 112.43kb.
На правах рукописи
Куликов Алексей Михайлович
ОСОБЕННОСТИ ЭВОЛЮЦИИ МОРФОЛОГИЧЕСКИХ И МОЛЕКУЛЯРНЫХ ПРИЗНАКОВ НА ПРИМЕРЕ
БЛИЗКОРОДСТВЕННЫХ ВИДОВ ДРОЗОФИЛ
03.02.07 – генетика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени доктора
биологических наук
МОСКВА 2012
Работа выполнена в лаборатории генетики Учреждения Российской академии наук Института биологии развития им. Н.К. Кольцова РАН
Научный консультант – доктор биологических наук, профессор
Митрофанов В.Г.
Официальные оппоненты: член-корреспондент РАН,
доктор биологических наук, профессор
Илья Артемьевич Захаров-Гезехус (ИОГен РАН)
доктор биологических наук, профессор
Елена Алексеевна Ляпунова (ИБР РАН)
доктор биологических наук, доцент
Гриценко Вячеслав Владимирович (ТСХА)
Ведущая организация: Учреждение Российской академии наук Институт молекулярной генетики РАН
Защита состоится 18 января 2012 г. на заседании диссертационного совета Д002.238.01, созданного при Учреждении Российской академии наук Институте биологии развития им. Н.К.Кольцова РАН по адресу: 119334, Москва, ул. Вавилова, 26.
Сайт: mcor.ru; E-mail: idbras@bk.ru
С диссертацией можно ознакомиться в библиотеке Учреждения Российской академии наук Института биологии развития им. Н.К. Кольцова РАН по адресу: 119334, Москва, ул. Вавилова, 26, и на сайте mcor.ru
Автореферат разослан________________2011 г.
Ученый секретарь диссертационного совета,
кандидат биологических наук Е.Б.Абрамова
E
 -mail: ele0806@yandex.ru
-mail: ele0806@yandex.ruАктуальность проблемы
Неравномерность темпов эволюции впервые была отмечена Дарвином и позже нашла отражение в сформулированных представителями школы эволюционной палеонтологии правилах и законах. Они предполагают связь гиперболического роста биоразнообразия и объема биоты с ростом устойчивости живых систем, зависимость периодов бурного видообразования и интенсивного вымирания видов от глобальных климатических изменений, наличие периодов быстрого и медленного накопления эволюционно-значимой изменчивости в родословных видов на длительных интервалах геологического времени. Изменения темпов эволюции прямо или косвенно связаны с действием отбора под влиянием факторов внешней среды, и заметны на масштабных отрезках геохронологической шкалы.
Все больше данных свидетельствует, что неравномерность темпов накопления эволюционно-значимой изменчивости характерна даже для относительно коротких временных промежутков, сопоставимых со средним временем эволюции вида, порядка нескольких млн. лет. Предполагается, что статусные, или эволюционно-значимые видовые признаки могут эволюционировать быстрее других, но остается неясным, насколько постоянной является скорость накопления видоспецифических различий по этим признакам и что определяет темпы накопления выявляемой изменчивости на морфологическом и молекулярном уровнях.
До настоящего времени остается нерешенным ряд проблем эволюционной генетики, связанных с оценкой темпов и механизмов накопления эволюционной изменчивости. Среди них: применимость концепции «прерывистого равновесия» Гулда и Элдриджа, т.е. чередования периодов стазиса и быстрой эволюции, к эволюции конкретных видов, признаков и геномов; возможность распространения концепции Сингха «эволюции по быстрому пути» («Evolution in fast lane»), рассматривающей эволюцию последовательностей генов, экспрессия которых ограничена полом, на эволюцию более широкого круга морфологических признаков (например, быстро эволюционирующих признаков копулятивного аппарата самцов); влияние отбора на эволюцию признаков, непосредственно не связанных с адаптациями; существование специфических генетических механизмов, определяющих скорость и направление эволюции признака. Известно, что нарушение темпов эволюции молекулярных признаков, или «хода молекулярных часов», характерно как для разных последовательностей одного генома, так и для гомологичных последовательностей удаленных видов, дивергировавших на протяжении десятков млн. лет. Остаются открытыми вопросы о минимально необходимом времени для формирования таких нарушений, и о том, насколько сопоставимы механизмы, нарушающие равномерную скорость эволюции молекулярных и морфологических признаков.
Экспериментальное исследование этих вопросов имеет принципиальное значение для проверки множества теоретических концепций эволюционной биологии. В отношении молекулярной филогенетики и биоинформатики полученные оценки предоставят возможность для более точной калибровки «молекулярных часов», и, соответственно, построения более достоверных реконструкций эволюционной истории различных групп организмов. Кроме того, ответы на поставленные вопросы могут иметь практическое значение, т.к. позволят более точно прогнозировать влияние различных факторов, в том числе антропогенного воздействия, на микроэволюционные процессы в популяциях.
Цель исследования:
Оценка темпов накопления генетической изменчивости, выявляемой при анализе морфологических и молекулярных признаков, на модели близнецовых видов дрозофил группы virilis, а так же определение механизмов, влияющих на темпы накопления изменчивости данных признаков.
Задачи:
- Оценить скорость и характер накопления межвидовой изменчивости морфологических признаков за время дивергенции близнецовых видов на примере эволюционно-значимых количественных признаков формы копулятивного аппарата и крыловой пластины дрозофил группы virilis, выявить роль генетической изменчивости в дивергенции видов по эволюционно-значимым признакам.
- Оценить скорость накопления видоспецифической изменчивости молекулярных признаков по данным анализа RAPD-маркеров и SNP-маркеров у видов дрозофил группы virilis.
- Сопоставить темпы накопления изменчивости молекулярных и морфологических признаков.
- На основе полученных данных провести анализ и верификацию различных концепций генетического контроля темпов эволюции на геномном и организменном уровнях.
Научная новизна.
Впервые определены количественные характеристики межвидовой изменчивости по форме копулятивного аппарата у дрозофил группы virilis, установлена морфофункциональная взаимосвязь между структурой быстро эволюционирующего копулятивного органа и половым отбором у дрозофил.
Показано, что характер дивергенции дрозофил группы virilis по форме копулятивного аппарата и форме крыла существенно различается. Представлены подтверждения концепции «эволюции по быстрому пути» признаков формы копулятивного аппарата дрозофил по сравнению с другими количественными признаками, в том числе признаками формы крыловой пластины. Эволюция признаков копулятивного аппарата самцов имеет направленный характер, т.е. вектор изменчивости по большинству количественных признаков формы органа сохраняется в ряду монофилетических видов. Эволюция признаков формы крыла не направлена и связана с адаптациями к активному полету.
Получено экспериментальное подтверждение концепции Добжанского об эволюционной роли хромосомных перестроек для накопления генетической изменчивости по эволюционно-значимым признакам. Впервые в эксперименте получено подтверждение концепции эволюции доминантности Фишера, как составной части эволюции генетической архитектуры, реализующейся на эволюционных отрезках в несколько млн. лет и более. Феномен эволюции доминантности объясняет направленный характер и неравномерный темп эволюции признаков формы копулятивного аппарата самцов дрозофил и в более общем виде – направленность и неравномерный темп эволюции количественных признаков.
Впервые показан неравномерный темп накопления молекулярной изменчивости в родословных близкородственных видов. Подтверждена взаимосвязь двух важнейших концепций молекулярной эволюции – неравномерности хода молекулярных часов и мозаичности геномов. Для проверки этого вывода нами впервые разработан метод прямого учета индекса дисперсии как меры равномерного хода молекулярных часов, учитывающий одновременно и поправку Гилеспи на популяционно-генетические факторы, нарушающие ход молекулярных часов, и поправку, связанную с недоучетом множественных и обратных замен в сравниваемых последовательностях (индекс инфляции Балмера).
Впервые экспериментально показано, что неравномерный темп эволюции морфологических и молекулярных признаков опосредован различными механизмами – действием отбора и механизмов эволюции доминантности в первом случае и действием генетико-автоматических процессов во втором.
Научно-практическая значимость работы
Работа носит фундаментальный характер и посвящена анализу механизмов нарушения равномерного хода эволюционной дивергенции видов, действующих на молекулярном, морфологическом и поведенческом уровнях. Представления о генетических механизмах, определяющих темп и направленность эволюционного процесса, являются необходимыми для оценки биоразнообразия и устойчивости животных и растительных сообществ, создания прогностических селекционных и эволюционных моделей.
Впервые проведена работа по сравнению темпов эволюционных изменений молекулярных и морфологических признаков на комплексе близнецовых видов со сравнительно короткой эволюционной историей. Выявленные в ходе работы закономерности накопления изменчивости по количественным признакам определяют систему общих требований к сравниваемым видам, используемым в качестве модели для изучения темпов эволюции. В частности, удобной моделью являются монофилетические виды, имеющие разное время существования в качестве независимого таксона. Предложен новый метод оценки скорости накопления молекулярной изменчивости, обладающий высокой точностью и применимый для близкородственных видов. Метод может быть использован для широкого спектра молекулярно-филогенетических исследований.
Основные положения, выносимые на защиту
1. Форма копулятивного аппарата самцов дрозофил группы virilis эволюционирует гораздо быстрее, чем признаки крыла. Действие отбора на этот признак осуществляется через брачное поведение. Изменчивость признаков формы крыловой пластины дрозофил группы virilis отражает адаптацию дрозофил к полету и не связана с дивергенцией данной группы.
2. Накопление видоспецифической изменчивости связано в наибольшей степени с видоспецифическими инверсиями.
3. Степень доминантности не постоянна, она меняется под действием отбора, связана с эпистатическими межлокусными взаимодействиями и прямо зависит от продолжительности эволюции данного вида.
4. Темп накопления изменчивости морфологических и молекулярных признаков на относительно коротких временных отрезках неравномерен.
5. Причины неравномерности темпов накопления изменчивости для обеих групп признаков различны: в первом случае они связаны с отбором, во втором – с действием генетико-автоматических процессов.
Внедрение результатов исследования в практику
Работы автора представлены на сайте «Проблемы эволюции» (iol.ru/) и используются студентами Государственных Университетов, а также аспирантами академических институтов, при подготовке по курсу «Теория эволюции» в качестве дополнительной литературы.
Степень личного вклада автора
Автор непосредственно участвовал в проведении всех экспериментов, на всех этапах: планирование и проведение эксперимента, оценка полученных результатов, статистическая обработка, создание новых оценочных алгоритмов.
Апробация работы. Результаты исследований и основные положения работы доложены и обсуждены на конференциях: 16th European Drosophila Res. Conf., Sept.29 - Oct. 2, 1999, Zurich. Switzerland; III Съезд ВОГиС ″Генетика в XXI веке: современное состояние и перспективы развития″, 6-12 июня 2004, Москва, 2004; 45th Annual Drosophila Research Conference, Washington, DC. 2004; 46th Annual Drosophila Research Conference, San-Diego, California March 30 – April 3, 2005; 47th Annual Drosophila Research Conference, Houston Texas 29.03-02.04,2006; 48th Annual Drosophila Research Conference Philadelphia, Pennsylvania March 7-11, 2007; 49th Annual Drosophila Research Conference San Diego, California, April 2–6, 2008; Genetics - understanding living systems. XX International congress of genetics. Berlin, Germany, July 12-17, 2008; 50th Annual Drosophila Research Conference, Chicago, Illinois, March 4-8, 2009; V съезд Вавиловского общества генетиков и селекционеров. Москва, 21-28 июня 2009 г., 51st Annual Drosophila Research Conference, April 7-11, 2010, Washington.
Публикации Опубликовано 19 работ по теме диссертации, в том числе 16 в журналах, рекомендованных Перечнем ВАК.
Объем и структура диссертации Диссертационная работа состоит из введения, обзора литературы, материалов и методов, результатов, обсуждения и списка цитированной литературы. Общий объем диссертации составляет 315 страниц, основной текст изложен на 262 страницах, включая 37 таблиц и 33 рисунка. Список цитированной литературы составляет 924 работы.
Материалы и методы исследования
Систематическое положение видов изучаемой группы. 11 видов группы virilis относятся к подроду Drosophila рода Drosophila, разделенных на две филады: virilis (D. virilis (Vi), D. lummei (Lu), D. americana (Am), D. novamexicana (No)) и montana (D. flavomontana (Fl), D. montana (Mo), D. lacicola (La), D. borealis (Bo), D. kanekoi (Ka), D. ezoana (Ez), D. littoralis (Li)). За исключением синантропного вида D. virilis, остальные виды группы обитают в зонах умеренного и субарктического климата Евразии и Северной Америки. Природные популяции D. virilis обитают в субтропических лесах Южного Китая, в зоне обитания предкового вида группы.
В работе использовано 34 изосамковые линии 11 видов дрозофил группы D. virilis.
Оценка внутри- и межвидовой изменчивости признаков формы копулятивного аппарата самцов была проведена на 25 линиях дрозофил группы virilis 11-ти близнецовых видов дикого типа, 15-30 образцов на линию, по 36 морфометрическим признакам (МП, рис. 1). Всего было проанализировано 278 препаратов. Морфометрический анализ проводили по фотографиям препаратов, сделанным в режиме сканирования на электронном микроскопе Jen-100C. Во избежание «эффекта шкалы» мы использовали индексы морфометрических показателей (в дальнейшем ИМП, или индексы), определяя их как отношение данного МП к МП 1.
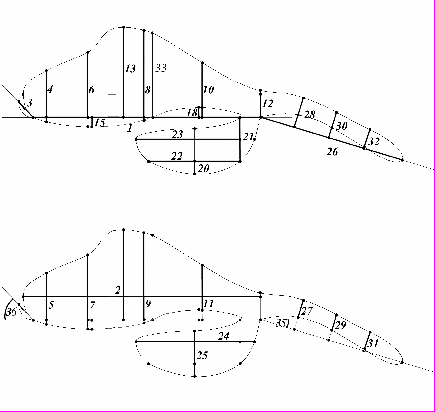


Рис. 1 Схема промеров копулятивного аппарата самца дрозофилы
Для анализа изменчивости признаков формы копулятивного органа самцов у межвидовых гибридов от скрещиваний трех линий D. lummei и линии D. novamexicana, с одной стороны, и линии D. virilis с рецессивными маркерами на аутосомах, с другой, были изготовлены и проанализированы 206 препаратов, от 15-и до 28-и препаратов на линию и на каждый из вариантов межвидового скрещивания. Гибридные самцы F1 от скрещиваний D. lummei × D. virilis были использованы для получения потомства из возвратных скрещиваний от самок D. virilis 160 и оценки участия хромосом в формировании видоспецифических признаков копулятивного аппарата самцов. В ходе этой работы было проанализировано 440 препаратов: 25 от самцов F1 из прямого скрещивания ♂ D. lummei x ♀D. virilis, 321 от самцов Fb – потомков F1 от прямого скрещивания, и 94 от самцов Fb – потомков F1 от обратного скрещивания.
Анализ внутри- и межвидовой изменчивости по признаку наличия микрохет на эдеагусе самцов дрозофил, а также участия хромосом в формировании данного признака, был проведен на 694 препаратах половых органов, полученных от 25 линий 11 близнецовых видов группы virilis, от гибридных самцов F1 из скрещиваний D. lummei 200 × D. virilis 160 и самцов Fb из возвратных скрещиваний самцов F1 на самок D. virilis 160.
Эффективность брачных ухаживаний в кон- и гетероспецифичных скрещиваниях оценивали по доле успешных копуляций из числа пар, осуществляющих стадию «садки», и по длительности копуляции. Было оценено по 30 пар в скрещиваниях видов D. virilis, D. lummei и реципрокных межвидовых скрещиваниях.
Форму крыловой пластины анализировали с помощью метода геометрической морфометрии (TPS v.1.37) с использованием 14 опознавательных точек (лэндмарок), маркировавших места пересечения жилок крыла и окончания их на краю крыла. Всего для анализа было использовано 1020 правых крыльев из 17 линий 11 видов дрозофил группы virilis, по 30 крыльев на линию.
Статистическая обработка результатов проведена с использованием программы “Statistica 6.1”. QTL-картирование полученных признаков проводили с помощью программы “Map manager QTX 0.30”, при выявлении значимых ассоциаций хромосом во всех случаях использовали значения χ2-критерия LRS-статистики для общего эффекта ассоциации P < 10-5 и для собственно эффекта взаимодействия P < 10-2 и post-hoc сравнениями в Главной линейной модели дисперсионного анализа. Для сравнения полиномиальных распределений использовали критерий Кульбака, в том числе для сравнения разных линий одного вида.
Для поиска генов-кандидатов на микрохромосоме D. virilis использовали известные по литературным данным последовательности SOP-энхансера, последовательность скаффолда 13052 D. virilis, соответствующего микрохромосоме, и программу “Vector NTI Advance 9.1”.
Экстракция и амплификация ДНК. ДНК экстрагировали из гомогената самцов стандартным фенол-хлороформным методом. Для проведения ПЦР-реакции использовался термоциклер PCH-3 (Techne, Великобритания). Условия амплификации были оптимизированы для каждого варианта ПЦР. Всего использовали 14 RAPD-праймеров, давших наиболее воспроизводимые видоспецифические паттерны, и одну пару праймеров, специфичных для амплификации фрагментов 1-го и 2-го экзонов и 2-го интрона гена Dras1 дрозофилы. Продукты реакции анализировались методом электрофореза в 2% агарозном геле. Секвенирование фрагмента гена Dras1 проводили дважды, с использованием прямого и обратного праймеров, набора ABI PRISM и ДНК-секвенатора 375А (Applied Biosystems, США). Полученные последовательности размещены в банке данных под номерами EF212396-EF212406.
Для анализа RAPD-маркеров составляли бинарные матрицы представленности маркеров в образцах и определяли генетические расстояния между видами и линиями по Нею и Ли (1979). Дальнейший анализ осуществляли методами кластерного анализа (UPGMA и NJ) и главных компонент, с использованием программ "PHYLIP" и "NTSYS". Филогенетический и молекулярный анализы по последовательности ДНК были проведены с использованием программы “MEGA version 4.1”.
При анализе равномерности накопления замен в родословных изученных видов дрозофил использовали программы “PAUP 4.0” (Swofford, 1998), “MODELTEST 3.7” (Posada, Crandall, 1998), “LINTREE” Takezaki (Takezaki et al., 1995) и “MEGA 4.1”. Все расчеты индекса дисперсии и критерия значимости χ2 его отклонения от единицы проводились в электронных таблицах “MS Excel”.
РЕЗУЛЬТАТЫ
1. Признаки копулятивного аппарата дрозофил группы virilis.
1.1. Межвидовая изменчивость признаков формы.
Размер фаллической части структуры, без учета аподемы, колебался в среднем от 0,136 мм (D. novamexicana) до 0,194 мм (D. kanekoi). Заметна общая тенденция уменьшения размера органа у видов D. lummei, D. novamexicana, D. americana, по сравнению с D. virilis, и наоборот, увеличение у видов филады montana. Эти результаты коррелируют с данными по морфометрии торакса (Spicer, 1993; Pitnick et al., 1995), и не совпадают с изменчивостью длины сперматозоидов представителей данной группы видов (Pitnick et al., 1995).
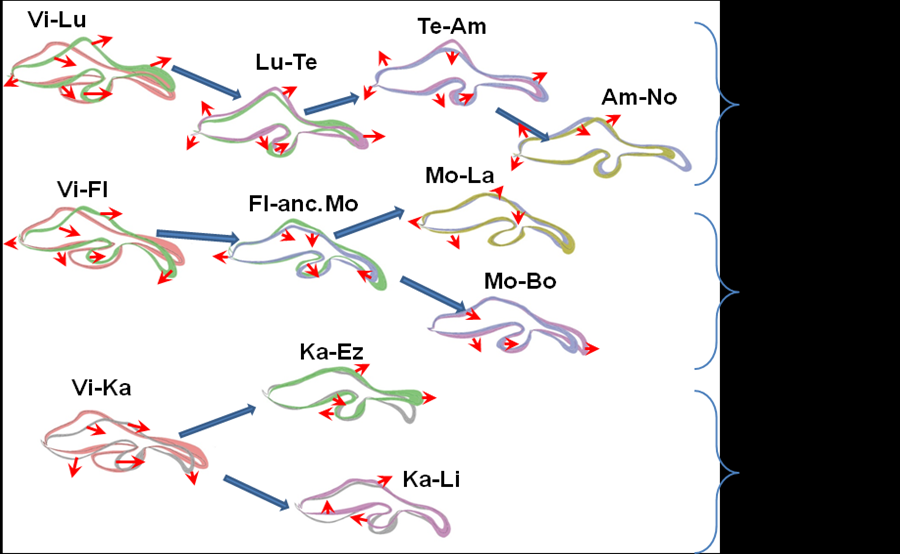
Рис.2 Направленная эволюция признаков формы копулятивного аппарата самцов в родословных видов дрозофил группы D. virilis.
По результатам анализа исследованных признаков в выборках из 11 видов и 2 х подвидов дрозофил построены схематические изображения проекций полового органа самцов для каждого вида. На рис. 2 приведены последовательные изменения признаков формы копулятивного аппарата в филогенетических линиях lummei, montana и kanekoi. Ширина каждого контура определяет 95%-ый доверительный интервал морфометрических показателей. Контуры предкового вида и вида-потомка на каждом этапе дивергенции наложены друг на друга. Стрелки показывают основные направления смещения морфологических структур. В каждой субфиладе происходили преимущественно однонаправленные смещения.
Оценка зависимости различий по исследованным признакам от факторов “принадлежность виду” и “принадлежность линии” была проведена с помощью дисперсионного анализа (MANOVA) по всей совокупности данных. В качестве гнездового использовали фактор “принадлежность виду”. Значение RRao, многомерного аналога F-критерия (RRao=17.94; df1=385; df2=2294; p<<0.001), показывает высокую значимость влияния переменной “принадлежность виду” на устойчивость различий по совокупности исследованных признаков.
При определении компонент дисперсии каждого индекса, факторы “принадлежность виду” и ”принадлежность линии” интерпретировались как случайные переменные. Компоненты дисперсии наиболее значимых ИМП (Fdf1,2 >50), определяемые переменной “принадлежность виду”, принимали на себя в среднем 60-80% общей дисперсии. Доля остальных индексов в общей дисперсии снижалась до 20%. Оценка их значимости подтвердила достоверное участие 32 ИМП из 35 в формировании видоспецифического паттерна. Индексы аподемы имеют слабую видовую специфичность.
Полученные результаты свидетельствуют, что в ходе эволюции близкородственных видов дрозофил группы virilis происходит преимущественно направленное накопление видоспецифической изменчивости по признакам формы копулятивного аппарата самцов.
Post-hoc сравнения в парах видов
С помощью post-hoc тестов Дункана (Duncan) и Шеффе (Scheffe) была оценена значимость различий видов по исследуемым признакам, и выделены группы видов со сходными групповыми средними по каждому признаку. Выявленная картина объединения видов по разным признакам оказалась неслучайной, и обладала следующими характерными чертами:
- по 11-ти ИМП вид D. virilis отличается от всех остальных. При этом 10 признаков имеют крайние значения по сравнению с остальными видами;
- еще по восьми ИМП этот вид, обладая крайними значениями признаков, тем не менее, оказался сходен с другими;
- у всех остальных видов независимость по групповым средним и их крайние характеристики встречаются значительно реже.
Сходство средних значений ИМП при парных сравнениях позволяет выделить наиболее близкие по признаку формы фаллоса пары видов: D. lacicola – D. borealis (нет различий по 24 ИМП), D. lacicola – D. littoralis (по 20 ИМП), D. americana americana – D. americana texana, D. lacicola – D. montana и D. borealis –D. montana (по 19 ИМП), D. lummei – D. novamexicana (по 18 ИМП). Эти пары в большинстве случаев представляют собой сочетание близкородственных видов дрозофил, согласно представлениям о филогенетических отношениях дрозофил группы virilis. Можно отметить также особое положение вида D. ezoana, расположенного по числу сходных ИМП между представителями филады virilis, за исключением собственно вида D. virilis, и филады montana. Эволюционно наиболее древние и наиболее молодые виды оказываются в диаметрально противоположных группах, выявляемых в ходе апостериорных сравнений, т.е. абсолютное большинство признаков эволюционируют направленно.
Приведенные данные подтверждают, что степень видоспецифических различий по признакам, описывающим форму копулятивного органа самцов, в целом соответствует представлениям о филогенетических отношениях видов данной группы. Накопление эволюционно-значимой изменчивости в разных эволюционных линиях происходило преимущественно в одном направлении.
1.2. Внутривидовая изменчивость признаков формы копулятивного аппарата дрозофил группы virilis
Для оценки межлинейной изменчивости признаков был проведен анализ влияния фактора “принадлежность линии” на внутривидовую изменчивость признака отдельно для каждого из видов: D. virilis, D. americana, D. lummei, D. borealis, D. montana и D. littoralis. Форма фаллоса имеет хорошо выраженную внутрилинейную специфику для каждого из исследованных видов. Количество ИМП, достоверно влияющих на фактор «Линия», менялось у разных видов от 11 до 21. Сходного для изученных видов набора признаков-маркеров межлинейной изменчивости среди ИМП парамеров и эдеагуса обнаружить не удалось. Так, у D. virilis основные межлинейные различия связаны с размерами и формой аподемы, у D. littoralis – эдеагуса, у остальных исследованных видов они распределены по всем частям органа. Значимость показателя межлинейной изменчивости λWilks определяли по наборам ИМП, включающим только достоверно различающиеся признаки. У видов D. littoralis (RRao, Form 2 =26,25; df1=13; df 2=5; p=0.001) и D. americana (RRao, Form 2 =38.96; df1=12; df 2=5; p<0.001) значения λWilks, трансформированные в показатель RRao, оказались высокими, значительно превысив общие характеристики межвидовых различий.
Эти данные подтверждают, что в популяциях исследованных видов сохраняется значительная изменчивость по различным признакам формы копулятивного аппарата, у разных видов эта изменчивость может зависеть от разных морфологических структур органа.
1.3. Соотношение межвидовой и внутривидовой изменчивости в группах скоррелированных признаков.
Для оценки латентной факторной структуры, определяющей сопряженную внутри- и межвидовую изменчивость индексов, мы предприняли разведочный факторный анализ данных. Критерии Каттела и Кайзера позволяют выделить от 4-х (первый) до 7 (второй) главных компонент и 6 главных факторов, полученных разными способами факторного анализа. Сходство нагрузок главных компонент и главных факторов свидетельствует об объективности выделенных групп признаков, принимающих на себя основную долю изменчивости. Результаты анализа показывают сходство вторичных факторных нагрузок, независимо от метода выделения главных факторов (табл. 1) и метода вращения факторных осей.
Табл. 1. Группы скоррелированных признаков формы копулятивного аппарата, выделенные с помощью ортогональных методов вращения.
| Способ выделения | Фактор 1 | Фактор 2 | Фактор 3 | Фактор 4 | Фактор 5 | Фактор 6 |
| I, II, III | 6, 7, 8, -10, -11, -12, 14, 17, 22, 23, 24, 34 | 2, 10, 11, 13, 33 | 28, 29, 30, 31, 32, 35 | 4, 5, 6 | 15, 19, 21, 25 | 7, 8, 9 |
| IV | 6, 7, -10, -11, 14, 17, 22, 23, 24, 34 | 28, 29, 30, 31, 32, 35 | 10, 11, 13, 33 | 8, 9 | 4, 5, 6 | 15, 21 |