
Особенности эволюции морфологических и молекулярных признаков на примере близкородственных видов дрозофил 03. 02. 07 генетика
| Вид материала | Автореферат |
- Молекулярные механизмы формирования множественной лекарственной резистентности у Burkholderia, 787.44kb.
- Рабочая программа генетика Код дисциплины по учебному плану опд ф. 6 для студентов, 274.5kb.
- Наследственность и ее роль в патологии медицинская генетика и ее задачи Наследственность, 327kb.
- Эволюция и Генетика, 167.01kb.
- Рабочая программа генетика и селекция Код дисциплины по учебному плану опд ф 1 для, 292.62kb.
- Практическая работа «Генетика», 116.83kb.
- Л. П. Татаринов необратимость эволюции и ее направленность, 194.79kb.
- Генетика Генетика, 18.18kb.
- Л. И. Параллелизмы в молекулярной организации генома и проблемы эволюции. В кн.: Молекулярные, 251.18kb.
- Рабочая программа дисциплины «молекулярная генетика» Код дисциплины по учебному плану, 112.43kb.
Principial axis factoring (I), Iterated communalities (MINRES) (II), Communalities=multiple R-square (III) и Maximum likelihood factors (IV).
В таблицу внесены ИМП, нагрузки которых на выявленные факторы превышают значения |0,5|. Жирным шрифтом выделены значения > |0,7|. ИМП, вошедшие с отрицательными нагрузками в данный фактор, занесены со знаком “минус”.
Анализ итоговых нагрузок ИМП на факторные оси позволил выделить следующие группы корреляций: 1-й фактор связан с сопряженной изменчивостью показателей высоты различных участков эдеагуса, положения “вершины” эдеагуса относительно точки 1 и длины парамеров; 2-й – ИМП 2, определяющего кривизну передней части эдеагуса над шипом, показателей высоты центральной и задней части эдеагуса и ИМП 15, характеризующего кривизну нижней передней части эдеагуса; 3-й определяет скоррелированную изменчивость характеристик аподемы; 4-й – независимую изменчивость ИМП, определяющих высоту передней половины эдеагуса; 5-й фактор определяет связь показателя кривизны нижней передней части проекции эдеагуса (ИМП 15), ширины парамеров у основания (ИМП 19) и показателей высоты парамеров (ИМП 21 и ИМП 25); наконец, 6-й фактор контролирует изменчивость показателей высоты среднего участка тела эдеагуса (ИМП 7-9).
В общей сложности 28 индексов проявили высокие значения корреляции с выявленными факторами, войдя со значительным весом в факторные нагрузки. ИМП 3, 16, 18, 20, 26, 27 и 36 не входят ни в одну из групп корреляций. При этом по данным дискриминантного анализа индексы 3, 18 и 36 вошли в состав хороших дискриминаторов, полученных с помощью методов последовательного отбора переменных – статистик F-включения или F-удаления. Среди признаков, вошедших в состав факторных структур, ИМП 2, 5, 11, 14, 33 и 34 выделяются как хорошие дискриминаторы. Кроме того, признаки 6, 7, 13 и 23 всегда отбираются в случае использования статистики F-удаления, и признаки 15 и 24 – в случае использования статистики F-включения. С учетом результатов post-hoc сравнений, следует отметить, что все ИМП, составляющие 1-ю факторную ось, определяют основные отличия D. virilis от остальных видов, а ИМП аподемы, входящие в состав 3-го фактора, определены как сходные в подавляющем большинстве сравниваемых пар. Сходство по ИМП, входящим в состав 2-го и 4-го факторов, встречается в среднем в 4-6 раз чаще при сравнении видов внутри филад virilis и montana, чем между этими филадами.
При анализе факторных структур внутривидовой изменчивости у видов D. virilis, D. lummei, D. montana и D. borealis были выделены общие для этих видов характеристики выявленных коррелирующих групп признаков. 1. Признаки объединены по принципу топологической близости и количество групп скоррелированной изменчивости больше, чем в случае анализа межвидовой изменчивости. 2. Индексы аподемы всегда входят в 1-й или 2-й факторы практически в полном составе. 3. ИМП, определяющие характеристики высоты проекции эдеагуса, могут быть организованы в 2-3 группы корреляции и часто объединены с ИМП 2, определяющим кривизну передней части эдеагуса над шипом. 4. Характеристики парамеров (ИМП 21-24) всегда представлены в факторной структуре в виде одной – двух корреляционных плеяд, не ассоциированных с ИМП эдеагуса и аподемы.
Сопоставляя меж- и внутривидовые факторные структуры, можно отметить, что прослеживается «модульная» организация эволюционирующего органа – эволюционно-значимая изменчивость независимо накапливается в разных, пространственно разобщенных частях органа, по-разному в разных эволюционных линиях. С этим же связано и «укрупнение» групп корреляции при переходе к межвидовой изменчивости – отбор подхватывает лишь часть изменчивости, как правило, ограниченной тем или иным компартментом. Изменчивость, связанная с признаками эдеагуса и парамеров, определяет преимущественно межвидовые различия. Изменчивость, связанная с признаками аподемы, определяет в основном внутривидовую изменчивость. Факторы 2, 4 и 6 включают в себя как внутри - так и межвидовую дисперсию. Ряд индексов, входящих в состав факторных структур внутривидовой изменчивости (3, 16, 18, 36), не скоррелированы с другими индексами в составе факторных структур межвидовой изменчивости, что свидетельствует об их качественном изменении на видовом уровне.
П
Рис.6 б
о признакам формы копулятивного аппарата были рассчитаны расстояния Махаланобиса, позволяющие количественно оценить расхождение между видами и линиями. Кластерный анализ был проведен по полученным расстояниям методами кластеризации UPGMA и Варда (рис. 3а). Виды группы montana кластеризуются отдельно от видов D. lummei, D. americana и D. novamexicana. Вид D. virilis кластеризуется отдельно от всех остальных видов, а D. kanekoi – близок к видам D. montana, D. flavomontana, D. lacicola, D. borealis и D. littoralis, но кластеризуется отдельно от них. D. ezoana близок к D. kanekoi по положению на дендрограмме, но в зависимости от способа кластеризации может быть равноудаленным от подгрупп montana и lummei, располагаясь при этом ближе к ним, чем к D. virilis.
Кластерный анализ, проведенный по всей совокупности используемых линий, в основном подтвердил отмеченную для видов степень сходства по форме копулятивного аппарата (рис. 3б). Отметим, что две линии D. littoralis имеют существенные различия, и независимо от способа кластеризации располагаются на дендрограмме на значительном удалении друг от друга.

б
а
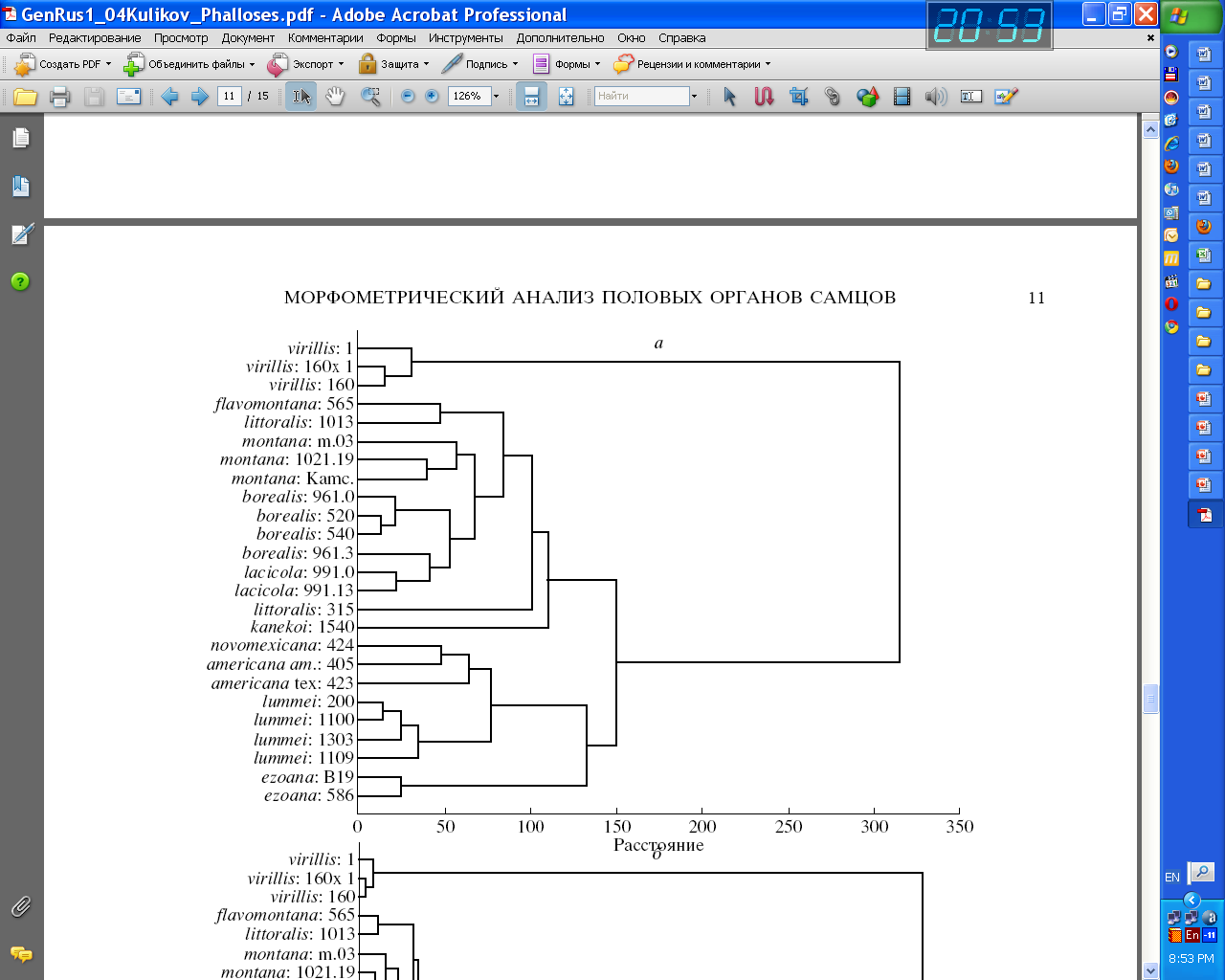
Рис.6 а
Рис. 3 Дендрограммы UPGMA, полученные для видов (а) и линий (б) дрозофил группы virilis по признакам формы копулятивного аппарата.
Сравнение полученной для видов группы virilis дендрограммы c филогенетическим деревом, построенным на основании анализа пяти последовательностей ядерных и митохондриальных генов (см. раздел «Оценки степени родства видов по молекулярным последовательностям») показывает топологическое сходство полученных деревьев. Отдельные эволюционные ветви деревьев различаются по длине от 11 до 43%, что приводит к единственному топологическому отличию: на представленной дендрограмме вид D. virilis находится за пределами своей собственной филады как внешний по отношению ко всем видам.
В целом, приведенные результаты факторного, дискриминантного и кластерного анализов хорошо подтвердили выводы, сделанные по результатам дисперсионного анализа ИМП. На протяжении всего времени дивергенции видов группы virilis форма копулятивного органа самцов была эволюционно значима и менялась направленно: происходило закономерное, но независимое в эволюционных линиях видов субфилад lummei, kanekoi и montana укорочение парамеров и смещение наиболее высокой точки дорсальной части эдеагуса в латеральном направлении.
1.4. Эволюция доминирования признаков формы копулятивного органа самцов дрозофил группы virilis.
Работа была проведена по 36 количественным признакам формы полового аппарата самцов: D. lummei 200, 1100 и 1109, D. novamexicana, D. virilis 160, самцов F1 из скрещивания ♂D. lummei200,1100,1109 × ♀D. virilis, самцов F1 из скрещивания ♂D. novamexicana × ♀D. virilis, самцов Fb с генотипом XVi/YVi, AVi/ALu200, самцов Fb с генотипом XVi/YLu200, AVi/ALu200. Статус признака определяли по результатам post-hoc сравнений в Главной линейной модели дисперсионного анализа. Основные результаты представлены в табл.2.
Табл.2 Доля признаков с различной степенью доминирования в скрещиваниях.
| Статус признака | Скрещивание, дизайн половых хромосом | |||||
| No x Vi XViYNo | Lu200 x Vi XViYLu | Lu1109 x Vi XViYLu | Lu1100 x Vi XViYLu | Lu200 x Vi XLuYVi | Lu200 x Vi XViYVi | |
| DVi+sDVi | 1712+5* | 7 | 8 | 12 | 7 | 8 |
| DLu+sDLu | 51+4* | 9 | 7 | 5 | 11 | 10 |
| N | 3 | 3 | 1 | 2 | 4 | 1 |
| ID | 8 | 11 | 13 | 11 | 10 | 8 |
| Ns | 5 | 8 | 9 | 8 | 6 | 11 |
| Σ | 38 | 38 | 38 | 38 | 38 | 38 |
DVi+sDVi – доминирование и сверхдоминирование признаков D. virilis;
DLu+sDLu – доминирование и сверхдоминирование признаков D. lummei или D. novamexicana; N – новый статус признака, ID – промежуточное доминирование;
*сумма в надстрочнике – количество признаков, проявляющих доминирование по данному фенотипу, в том числе: доминировавших в скрещиваниях No × Vi и Lu × Vi, уникальных только для скрещивания No × Vi.
Выявлены следующие закономерности:
1. Доля признаков у потомства от прямого и обратного скрещиваний D. lummei × D. virilis, доминирующих или даже сверхдоминирующих по фенотипу D. virilis, примерно соответствует доле рецессивных признаков;
2. У гибридов от скрещивания D. virilis × D. novamexicana существенно возрастает доля признаков, имеющих доминирование родительского фенотипа D. virilis, по сравнению с гибридами D. virilis/D. lummei. Гомологичные признаки, имеющие у гибридов D. lummei/D. virilis доминирование D. virilis, сохраняют этот статус и в скрещиваниях ♂D. novamexicana × ♀D. virilis. Признаки, имеющие доминирование второго родительского фенотипа, D. lummei или D. novamexicana, в основном не сохраняют преемственности своего статуса в разных скрещиваниях. То есть, признаки, потерявшие на данном этапе дивергенции доминантный статус, оставались рецессивными и на более поздних этапах дивергенции. Наоборот, доля вновь возникающих доминантных признаков у молодых видов снижалась одновременно с уменьшением времени независимой эволюции молодого вида;
3. В среднем, 35% признаков проявляют сходную степень доминирования в скрещиваниях ♂D. lummei x ♀D. virilis и ♂D .novamexicana x ♀D. virilis, тогда как при сравнении скрещиваний различных линий D.lummei в направлении ♂D. lummei x ♀D. virilis сходством обладают уже 53% признаков. При этом степень доминирования индивидуальных признаков в скрещиваниях D. virilis с линиями D. lummei, имеющими происхождение из разных частей ареала обитания, может существенно различаться;
4. Потомство F1 от скрещивания ♂D. lummei200 × ♀D. virilis имеет генотип (XViYLu АViАLu), идентичный полностью гетерозиготному по аутосомам потомству Fb от возвратного скрещивания самцов F1 с самками D. virilis. Тем не менее, 17 из 38 признаков показывают достоверную разницу по доминированию для самцов F1 и Fb, что позволяет предположить значимое влияние на экспрессию этих признаков отцовского генотипа;
5. Состав половых хромосом у гибридных самцов от скрещиваний D. lummei200 × D. virilis влияет на степень доминирования признаков. Замена Х-хромосомы D. lummei на Х-хромосому D. virilis в возвратном скрещивании самцов F1 из потомства скрещивания ♂D. virilis x ♀D. lummei 200, на самку D. virilis, приводит к изменению статуса доминирования половины из всех проанализированных признаков. Тем не менее, соотношение признаков, проявляющих доминирование родительского генотипа D. virilis к признакам, проявляющим доминирование генотипа D. lummei, практически не меняется: 7/11 для самцов F1 от скрещивания ♂D. virilis x ♀D. lummei 200, и 8/10 для самцов F2 от возвратного скрещивания.
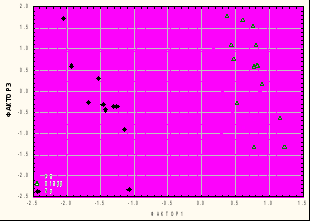
Результаты анализа ИМП хорошо подтверждаются многофакторными методами анализа, учитывающими скоррелированную изменчивость, что существенно повышает надежность выводов. Для примера приведены распределения вторичных признаков гибридных и родительских форм в пространстве гомологичных факторных структур (Ф1 - большая часть признаков полового аппарата, кроме аподемы; Ф2(3) – признаки аподемы), для скрещиваний Vi × No (рис.4 а) и Vi × Lu200 (рис.4 б).
Хорошо видно доминирование фенотипа D. virilis в первом случае, и D. lummei во втором, в пространстве значений 1-го, наиболее весомого фактора. Полученные результаты свидетельствуют, что в филогенетической ветви D. lummei → D. novamexicana происходило накопление рецессивной изменчивости, причем преимущественно из-за потери доминирования гомологичных признаков у эволюционно более молодого вида.
а
б
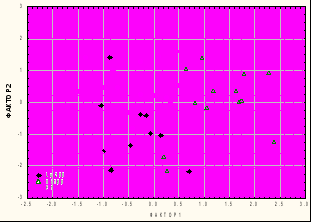
Рис. 4 Распределение родительских и гибридных F1 генотипов в плоскости факторов 1 и 2: а – скрещивание ♂No × ♀Vi; б – скрещивание ♂Lu200 × ♀Vi.
Несмотря на статусную роль признаков формы копулятивного аппарата, в популяциях D. lummei сохраняется значительный полиморфизм по степени доминирования данных признаков, что дополняет сделанное выше наблюдение о сохраняющемся полиморфизме этих признаков в географически удаленных популяциях разных видов дрозофил. Наконец, состав половых хромосом у гибридных самцов влияет на степень доминирования признаков. Факторный анализ максимизирует влияние половых хромосом: в реципрокных скрещиваниях F1 замена ХVi на ХLu и Y-хромосомы в противоположном направлении приводит к двукратному снижению доли объясненной изменчивости, проявляющей доминирование фенотипа D. virilis (с 9.6 до 4.5%), и двукратному увеличению доли изменчивости, проявляющей доминирование фенотипа D. lummei (с 27 до 48%). Наименьшей долей изменчивости, приходящейся на признаки, проявляющие доминирование фенотипа D. lummei, и наибольшей – фенотипа D. virilis, обладают самцы FbXViYVi, несущие обе половые хромосомы D. virilis.
Эти результаты хорошо подтверждают гипотезу Фишера о механизмах эволюции доминирования за счет эпистатической изменчивости (подбор модификационных, т.е. регуляторных систем, повышающих степень доминирования эволюционно-значимых признаков).
1.5. Хромосомная локализация признаков формы копулятивного аппарата самцов дрозофил группы virilis
Используемые для оценки формы признаки могут иметь сходную изменчивость, определяемую одним и тем же набором локусов. Поэтому для выявления таких групп признаков мы провели факторный анализ изменчивости, характеризующий все используемые генотипы.
По результатам анализа методом максимального правдоподобия, наиболее значимая факторная структура представлена признаками длины парамеров (ИМП 24, 25 и 26) и соотношения длины и максимальной ширины парамеров. Структура 2-го фактора определена исключительно признаками аподемы. В состав 3-го фактора входят преимущественно показатели формы дорсальной поверхности центральной и задней части фаллоса – ИМП 8-16 и индексы 13/14, 15/16. Топологически близкие показатели формы дорсальной поверхности передней части фаллоса – ИМП 4-7 и ИМП 14 и 16 образуют факторную структуру 4-го фактора. В составе структуры 5-го фактора наибольшими весами обладают ИМП 17-20, определяющие форму вентральной поверхности фаллоса. Выявленные факторные структуры хорошо соответствуют показанным выше наиболее значимым факторам, определенным для внутривидовой изменчивости видов D. virilis и D. lummei.
Роль межвидовой изменчивости в составе факторных нагрузок иллюстрирует распределение генотипов Fb, представленных в анализируемой выборке, в пространстве значений факторов (рис. 5).
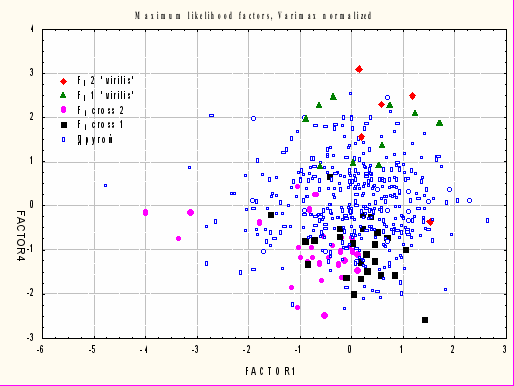
Самцы из возвратного скрещивания: Fb2 “virilis” – XVi/YVi, AVi/AVi; Fb2 “virilis” – XVi/YLu, AVi/ALu; Другой – частично гомозиготные по аутосомам D. virilis.
Самцы F1: F1 cross 2 – XLu/YVi, AVi/ALu; F1 cross 1 – XVi/YLu, AVi/ALu
Рис. 5. Распределение генотипов из возвратных скрещиваний ♀Vi × ♂Lu/Vi в плоскости факторов 1, 4.
Альтернативными генотипами в составе выборки являются полностью гетерозиготные самцы, обладающие в полной мере наборами аутомосом от обоих родительских видов, и гомозиготные по геному D. virilis самцы. Облака изменчивости переменных по 1-му и 4-му факторам, характеризующие полностью гетерозиготных и гомозиготных самцов, практически не пересекаются. Промежуточные варианты при этом плотно занимают все пространство между двумя крайними вариантами.
Применив регрессионный анализ значений признаков на гомо/гетерозиготный статус хромосом для модели возвратных скрещиваний, мы определили вклад отдельных хромосом в формирование видоспецифических признаков копулятивного аппарата.
Признаки формы парамеров (фактор 1). Признак 24: хромосома 6 – 5-8% от общей изменчивости, хромосомы 2 и 5 – 5-6%. Признаки 25, 26, 27: хромосома 5 – 13-24%, хромосомы 2 и 6 – 9-18% от общей изменчивости.
Признаки положения аподемы (фактор 2). Эти признаки являются слабыми дискриминаторами. Некоторый вклад аутосом в видоспецифическую изменчивость удалось получить для ИМП 28, 29 и β. Она составляет 13%, 4% и 9% соответственно. Для ИМП 28 и 29 наибольшее влияние оказывают хромосомы 5 (8-13%) и 4 (4-6%) соответственно. Признак β находится под контролем хромосом 2 – 21% и 6 – 3%, отмечено также влияние Y-хромосомы на экспрессию признака.
Признаки формы дорсальной поверхности центральной и задней части фаллоса (фактор 3). Признаки 8, 9: хромосома 2 до 7%, хромосома 3 – 4%, хромосома 6 – 2-6% от общей изменчивости. Признаки 10, 11, 14: хромосома 3 – 14-20%, хромосома 2 – 2-22%, хромосома 4 – 11-16%, хромосома 6 – 5-10% от общей изменчивости.
Признаки формы дорсальной поверхности передней части фаллоса (фактор 4). Влияние хромосом для всех признаков данной группы является очень сходным. Признаки 4-7, 16: хромосома 5 – 13-23% от общей изменчивости и хромосома 3 – 10-17%.
Признаки формы вентральной поверхности фаллоса (фактор 5). Признак 17: ведущая роль хромосомы 5 (14%). Признак 20: ведущая роль хромосомы 3 (7-15%), второстепенная роль хромосом 6 (до 11%) и хромосомы 4 (3-8%). Для обоих признаков играет роль регуляторное влияние Y-хромосомы. Для ИМП 18 и 19 во всех вариантах анализа однозначно выявляется основная роль хромосомы 2 (21% и 14% от общей изменчивости соответственно), второстепенную роль играют хромосомы 3 (4%) и 6 (2-6%).
Признаки с высокой характерностью. Для ИМП 23, признака высоты парамеров у их основания, ведущую роль в формировании генетической составляющей общей дисперсии играет хромосома 5. Признаки шипа на конце эдеагуса, ИМП 3 и α, являются сильными дискриминаторами и принципиально различаются по вкладу аутосом в выявленную генетическую составляющую дисперсии. Ведущую роль в формировании дисперсии признака ИМП 3 у самцов Fb от прямого скрещивания играют хромосомы 3 и 2 (11% каждая), а для признака α – хромосомы 4 и 6 (7% каждая). Для самцов Fb от обратного скрещивания наиболее весомый вклад в контроль признака ИМП 3 имеют хромосомы 3 и 6 (22% и 12%, соответственно), признака α – хромосомы 2 и 4 (30% и 26%, соответственно). Учитывая наблюдаемые различия, оба признака зависят от эффектов взаимодействий Y-хромосомы с аутосомами.
Оценка неаддитивных взаимодействий хромосом в формировании видоспецифической изменчивости показала, что 26 из 38 признаков проявляет неаддитивную изменчивость, она может составлять до 97% от наблюдаемого эффекта влияния пары хромосом.
Из 13 признаков, для которых показан эффект эпистатических взаимодействий, 9 попадают в группу доминирования фенотипа D. lummei, причем это практически все признаки, доминирующие по эволюционно новому фенотипу. Очевидно, эпистатические взаимодействия межу локусами из разных групп сцепления необходимы для проявления эффекта доминирования. Этот результат подтверждает вывод о роли эпистатических взаимодействий в эволюции доминирования признаков с новым видовым статусом.
Как определить ожидаемый вклад хромосом в межвидовую изменчивость признаков формы копулятивного аппарата? Используем известную гомологию хромосом D. virilis и D. melanogaster по элементам Меллера и функциональную значимость и экспрессионную активность всех аннотированных генов D. melanogaster. В соответствии с данными FlyBase, из 302 SRR (sex and reproduced related) генов на 2-й – 6-й аутосомах расположено 34%, 17%, 24%, 24% и 1% этих генов соответственно. Сходным образом, из 60 известных генов, ограниченных экспрессией в тканях самцов, на 2-й – 6-й аутосомах расположено 35%, 27%, 17%, 20% и 2% от общего их числа соответственно. Оценим распределение по хромосомам объясненной изменчивости по видоспецифическим признакам формы копулятивного аппарата. Группы признаков, обладающих скоррелированной изменчивостью, имеют различный объем, и каждый признак, обладающий высокой характерностью, представляет собой самостоятельную группу. Поэтому следует последовательно взвесить показатели участия хромосом для каждого признака на размер группы скоррелированной изменчивости, включающей данный признак, и затем получить средние значения объясненной изменчивости по признакам для каждой хромосомы. Ожидаемые показатели определяются как доли от суммарной средней объясненной изменчивости, соответствующие двум наборам приведенных выше теоретических оценок. Результаты расчетов представлены в табл. 3.