
И. И. Ш м альгаузен избранные труды организм как целое в и ндивидуальном и и сторическом издательство «наука» москва 1982 развитии
| Вид материала | Книга |
- Сергей Лычагин «Макиавелли Н. Избранные произведения.», 922.62kb.
- И. И. Веселовског о издательство "наука" Москва 1967 Эта книга, 1700kb.
- Избранные научные труды. М.: Наука, 1988, с. 722-734, 363.13kb.
- А. Н. Леонтьев Избранные психологические произведения, 6448.08kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 206.83kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 238.78kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 208.81kb.
- Д. Н. Мамине-Сибиряке Книга, 262.07kb.
- Литература Особенности типологического подхода и метода исследования личности, 2747.84kb.
- А. Н. Леонтьев Избранные психологические произведения, 6931kb.
Изучение эмбриональной изменчивости показывает, что изменчивы все стадии развития и что особенно часто встречаются сдвиги во времени закладки органа, во времени его морфологической и гистологической дифференцировки. Если бы все эти сдвиги накапливались независимо друг от друга в виде бесконечной цепи гетерохронии, определяемых эволюцией взрослого организма (путем, например, естественного отбора), то мы получили бы приблизительно ту картину онтогенеза, какую нам нарисовал Э. Менерт,— ряд независимых, протекающих бок о бок морфогенетических процессов, скорость которых пропорциональна «филетическим энергиям», измеряемым уровнем филогенетического развития органа. Рекапитуляция, по Менерту, происходит лишь в пределах отдельных органогенезов, и закономерности последних определяются «филетическими энергиями». В результате Э. Менерт приходит к чисто механо-ламаркистским выводам.
Мы должны отметить, что рекапитуляция касается иногда и целого организма (вначале развивается гаструла с первичной кишкой, позднее — вторичная полость тела и кровеносная система; очень рано — закладка нервной системы, позднее — закладка дыхательной и выделительной), чаще она проявляется в известной системе органов (порядок развития в скелете позвоночных: хорда, осевой скелет, позднее — скелет конечностей, притом сначала соединительнотканный, затем хрящевой и, наконец, костный скелет) и еще чаще, правда, рекапитуляция выражается в частных особенностях развития отдельных органов. Однако с последовательностью в развитии отдельных частей внутри определенного органа дело тоже обстоит не вполне гладко — далеко не так, как этого требует биогенетический закон. Последовательность в закладке отдельных элементов скелета пятипалой конечности ни в какой мере не соответствует последовательности филогенетического появления этих частей. Мы должны прямо сказать, что сдвиги во времени закладки как органов, так и их частей так часты, что на последовательности в развитии органов или их частей нельзя было бы построить никакой теории рекапитуляции. Последняя и построена не на этих фактах, а на фактах повторения (рекапитуляции) известных соотношений, зависимостей. Поэтому рекапитуляции особенно часты в системах, аппаратах, органах сложного состава, части которых связаны много-
74 Организм нак целое в индивидуальном и историческом развитии
численными взаимозависимостями (голова в целом, в частности висцеральный аппарат в целом, конечности в целом, кровеносная и нервная система в их соотношениях с другими органами и т. д.). Всякая теория рекапитуляции должна учитывать эти факты. Рекапитуляция связана с взаимозависимостями, с целостностью развивающегося организма. Это подтверждается также тем, что рекапитуляция выражается гораздо более полно при регуляционном развитии, характеризующемся сложными взаимозависимостями частей (позвоночные), чем при мозаичном с его относительно автономными органогенезами.
Так как и системы градиентов внутри еще недифференцированного яйца или любого недифференцированного позднейшего зачатка имеют регуляторный характер, то нарушения в их величине, в их физиологической активности и во времени обособления нередко выравниваются и в дальнейшем не проявляются в заметных нарушениях развития. Точно так же и индукционные системы допускают в известных пределах изменение физиологической активности и взаимное смещение взаимодействующих компонентов как в пространстве, так и во времени их «созревания». Поэтому многочисленные малые мутации, на которых в основном собственно и строится процесс эволюции, нередко не оказывают заметного влияния на течение важнейших морфоге-нетических процессов. Формообразование идет в течение ранних стадий эмбрионального развития нормально и только на более поздних стадиях постепенно начинает проявляться влияние данной мутации.
Эти соображения заставляют нас отвергнуть точку зрения Т. Моргана. Все гены участвуют в процессах клеточного обмена веществ и, следовательно, активны с самого начала развития яйца. Любая мутация связана с изменениями в биохимических процессах, протекающих в тканях самых молодых эмбрионов. Однако подавляющее большинство тех небольших мутаций, которые действительно имеют значение в процессе прогрессивной эволюции, вызывают сдвиги в реакциях, не выходящие за пределы пороговых уровней нормального течения процессов раннего морфогенеза, т. е. на эмбриональных стадиях развития ничем себя не проявляют. Их действие сказывается лишь на более автономных процессах, в особенности в конце морфогенеза, когда реакции, связанные с развитием менее существенных признаков, не столь зависимы от других морфогенетических процессов и не имеют столь выраженного регуляторного характера, как основные процессы формообразования на ранних стадиях эмбриогенеза.
Это объясняет нам более легкую изменяемость поздних стадий онтогенеза и относительную консервативность ранних стадий, что и лежит в основе зародышевого сходства не только близких, но и более отдаленных представителей животных, входящих в такие крупные их подразделения, как разные классы, типы,
Глава II. Эмбриогенез 75
а отчасти и между представителями разных типов (сходство в процессах гаструляции и обособления зародышевых листков).
Однако само собой разумеется, что и соображения Ч. Дарвина о крупных нарушениях, вносимых в процессы индивидуального развития при изменении ранних его стадий, полностью сохраняют свое значение. Если сдвиги в корреляционных системах выводят взаимодействующие компоненты за известные пределы, контакт между ними уже не осуществляется в полной мере. При значительных сдвигах во времени их созревания нормальная реакция также не может осуществиться. Либо концентрация морфогенных веществ во время контакта еще не достигла нижнего порога чувствительности реактора или, наоборот, превысила верхний порог его нормального реагирования. С другой стороны, реактивная способность может не достигнуть нормального уровня чувствительности или, наоборот, эта способность может, ко времени наступления контакта, уже оказаться утраченной. Во всех этих случаях произойдут значительные нарушения нормального развития, в особенности, если эти сдвиги за пределы пороговых уровней проявятся на ранних и ответственных морфо-генетических процессах.
[«Консервативность» ранних стадий выражается только в морфогенетических системах, определяющих развитие нормальной организации, и, конечно, совершенно не исключает свободной адаптации эмбриона или личинки к их условиям существования. Это лишь подтверждает правильность нашей точки зрения на значение корреляций и регуляторных систем, «защищающих» морфогенез].
Мы уже отмечали, что при наличии сложной системы морфогенетических корреляций большие сдвиги в этой системе (связанные с более значительными мутациями), т. е. изменения в скорости развития взаимодействующих частей или значительные их смещения, очень легко приводят к недоразвитию частей и, следовательно, к дефектам в организации (см. дальше рис. 11 Для редукции органов). Соответственно большинство рано проявляющихся мутаций характеризуется более или менее глубокими нарушениями строения организма. Наибольшие нарушения получаются при значительных сдвигах в основных, детерминирующих системах (полное выпадение) или при сдвигах в индукционных системах, определяющих ранние стадии дифференцировки зачатка. Если учесть еще последовательность установления взаимозависимостей между отдельными органогенезами, то станет ясно, что крупные сдвиги на ранних стадиях развития эмбриона приводят к глубочайшим нарушениям всего эмбриогенеза, к невозможности дальнейшего развития (остановка и гибель; обычная картипа летальных мутантов, например, гомозиготных короткохвостых мышей) или, иногда, к развитию уродов, рано или поздно все же погибающих. Мы видели, что все это обычные яв-
76 Организм как целое в индивидуальном и историческом развитии
ления. Однако они, как указал еще Ч. Дарвин, не имеют никакого значения в эволюционном процессе именно потому, что все эти уклонения гибнут, не оставляя потомства. Мутации, выражающиеся в заметных сдвигах на ранних стадиях развития, могут приобрести значение в эволюционном процессе лишь в том случае, если они касаются очень рано автономизирующейся системы органов, органа или части (первичная кишка, эпидермис позвоночных). В таком случае возможно ценогенетическое развитие новых образований и на ранних стадиях (амнион, сероза, аллантоис). В таких автономных частях мы тогда не найдем и явления рекапитуляции анцестральных состояний.
Наоборот, в конечных стадиях отдельных органогенезов вполне возможны и значительные сдвиги без глубоких нарушений развития всего организма, так как чем дальше идет развитие органа, тем более оно становится автономным. Впрочем, и такие изменения не остаются изолированными благодаря существованию известных зависимостей, главным образом эргон-тических, определяющих окончательную отшлифовку органов. Однако эти изменения могут оказаться без заметного влияния на жизнеспособность организма. Если при этом основное изменение благоприятно для организма в его соотношениях с внешней средой, то оно может приобрести положительное значение в борьбе за существование, а не совсем благоприятные побочные явления могут сгладиться в процессе дальнейшей эволюции (в результате подбора модификаторов). Сказанное дает дополнительное объяснение «консервативности» ранних стадий развития и «закону терминального изменения» [A. Naef, 1917]. Изменение ранних стадий приводит обычно к недоразвитию и гибели, изменение поздних стадий может привести и к благоприятному для организма результату. Изменение конечных стадий развития — это северцовские надставки, или «анаболии». Мы знаем, что именно они ведут к рекапитуляции анцестральных состояний. И мы должны отметить, что такие анаболии только и возможны в сложно связанных органогенезах. Соответственно мы действительно наблюдаем наиболее полные рекапитуляции главным образом в сложных аппаратах и органах, тесно связанных с другими частями организма, как нами на это обращено внимание (см. рис. 7).
Так как в филогенезе, таким образом, фактически изменяются главным образом поздние стадии развития эмбриона, а ранние стадии испытывают лишь незначительные в общем изменения (и то в рано автономизирующихся частях), то ранние стадии развития эмбрионов всех позвоночных в общем довольно сходны. Отмеченное К. Бэром «типичное» сходство ранних стадий развития эмбрионов даже далеких представителей одного типа находит свое объяснение в сложности системы эмбриональных корреляций регуляторного характера, обеспечивающей нормальное
Глава II. Эмбриогенез 77
(формообразование при небольших мутациях и в постоянной элиминации более значительных мутаций, коренным образом нарушающих весь процесс эмбриогенеза.
В индивидуальном развитии организма наиболее тесно связаны первые процессы, определяющие детерминацию основных органов. Эти зависимости наиболее ответственны и в наименьшей степени допускают какие-либо нарушения. В дальнейшем онтогенез вступает в «мозаичную» фазу, характеризующуюся «самодифференцированием» уже детерминированных органов. Отдельные органогеыезы теперь автономизированы. Теперь уже возможны и некоторые более заметные сдвиги и нарушения без того, чтобы серьезно пострадало развитие всего организма. Возможны «девиации». Возможны даже и «архаллаксисы», т. е. сдвиги во времени формирования уже детерминированных зачатков, и тем более возможны, чем более данная самодифференцирующаяся система независима от других.
Понятия «самодифференцирования» и «мозаичной» фазы развития имеют, однако, лишь относительное значение. Ни один органогенез не является вполне автономным (как мы это уже рассматривали выше) и соответственно далеко не всякие изменения зачатков и уклонения в их развитии проходят без особых нарушений в развитии других частей. Поэтому «архаллаксисы» все же относительно редки, особенно в более тесно связанных системах, чаще наблюдаются «девиации», а еще чаще «анаболии». Анаболии представляют собой надстройки, связанные с установлением новых корреляций, которые прямо не могут быть включены в уже существующую сложную систему взаимозависимостей. Новые корреляции могут быть введены лишь на тех стадиях развития, когда основные морфогенетические связи уже утратили свое значение. Поэтому анаболии наименее нарушают строение других частей или органов, связанных с изменяемыми органами посредством морфогенетических, или — на этих стадиях уже чаще — эргонтических корреляций. Эволюция посредством анаболии связана, однако, как показали Фр. Мюллер и А. Н. Северцов, с рекапитуляцией анцестральных состояний. Так как система морфогенетических зависимостей имеет не во всех частях и органах одинаковое значение, то и эволюция этих частей и органов может идти по-разному — в более автономных частях путем ар-халлаксиса, в частях со сложными взаимозависимостями — путем анаболии. Следовательно, в более автономных системах река-пйтуляция наблюдается реже, а в системах зависимых рекапи->ляция оказывается тем более полной, чем сложнее эти зависимости (рис: 6).
Эпидермис и его производные автономизируются у позвоноч-
1х очень рано. Поэтому в этой системе возможны довольно сво-
Дные и ранние изменения. Возможны изменения самих зачат-
' т- е. архаллаксисы. Это, конечно, не означает их полной не-
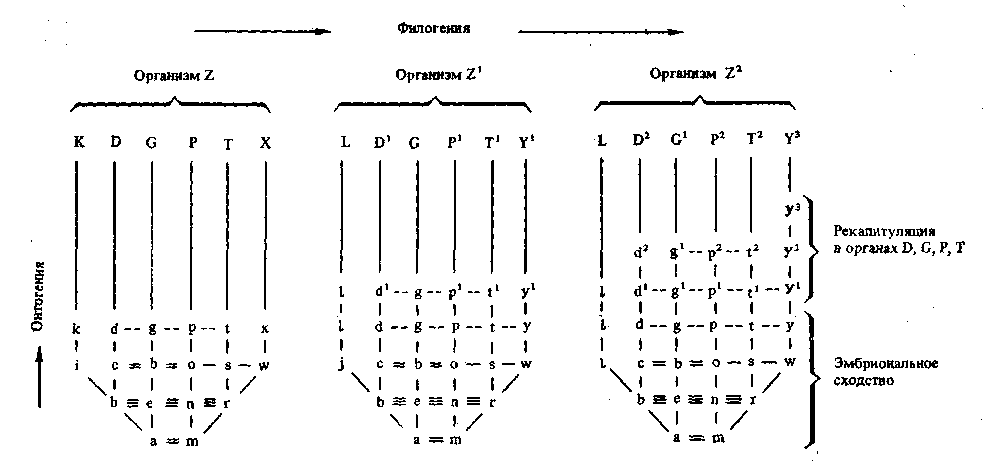
Рис. 6. Схема филогенетических изменений онтогенеза при наличии взаимозависимостей между органогене-зами
Морфогенетические корреляции показаны горизонталь- В рано автономизирующемся органогенезе, ведущем
ными сплошными линиями, эргонтические — прерыви- к образованию органа х эволюция идет путем девиации
отыми. Большие буквы — отдельные органы взрослого и приводит к образованию органа Y3 развивающегося при
животного. Малые буквы — стадии онтогенетического явлениях частичной рекапитуляции (фаза X утеряна),
развития этих органов. На схеме показано автономное В зависимых органогенезах, ведущих к образованию ор-
развитие органа k и его изменение в L путем архаллак- ганов D, G, Р, Т, эволюция идет путем анаболии и свя-
сиса (зачаток г изменился в j). В этом случае у потомка зана с ясной рекапитуляцией анцестральных состояний
Z2 нет рекапитуляции анцестральных состояний г, h. {d—d1—сР, g—£', р—р1—р2, t—li—t2)
Глава II. Эмбриогенез 79
зависимости. Наоборот, на примере развития оперения ног и развития хохла на голове у кур мы могли убедиться в противном, но все же здесь зависимости не столь сложны, чтобы служить препятствием для изменений даже иа ранних стадиях развития, g этом случае, однако, как мы знаем, нет предпосылок для повторения исторических фаз развития. И действительно, в системе эпидермиса ясные картины рекапитуляции анцестральных состояний наблюдаются редко. Вспомним, что до сих пор морфология не вскрыла еще вопроса о происхождении такого молодого образования, как волосы млекопитающих, и это именно потому, что эмбриональное развитие волоса не дает ясной опоры для суждения о его происхождении, Поэтому существующие теории происхождения волоса и построены на принятии эмбриональных изменений самих зачатков.
Относительно рано автономизируется у позвоночных, по-видимому, также энтодерма как замкнутая в себе система. В развитии большинства производных кишечника мы и в самом деле не находим ясной рекапитуляции (печень, поджелудочная железа). В развитии легких у рептилий, птиц и млекопитающих сразу появляются характерные черты класса, и последовательное почкование бронхов у млекопитающих совершенно отлично от филогенетического хода постепенного усложнения системы трабекул.
Исключением, подтверждающим правило, является развитие жаберных мешков, в котором мы имеем один из классических примеров рекапитуляции (рис. 7). Однако как раз здесь установилась весьма сложная зависимость между отдельными частями разного происхождения, именно: между энтодермальной закладкой жаберных мешков, эктодермальной закладкой жаберных щелей и жаберных лепестков, эктомезодермальной закладкой скелета, энтомезодермальной закладкой мускулатуры. Эти зависимости усложнены еще связью с развитием артериальных дуг, жаберных сосудов и висцеральных нервов. Одним словом, здесь как раз имеется та чрезвычайно сложная взаимозависимость частей, которая, как мы признали, является необходимой предпосылкой Для более полной рекапитуляции (рис. 7).
В мышечной, нервной и кровеносной системах наблюдаются вообще весьма постоянные взаимозависимости на всех стадиях развития. Большинство филогенетических изменений наступает здесь на последних стадиях морфогенеза, т. е. идет по способу знаболий. В результате в эмбриональном развитии нервной системы, мышечной и кровеносной мы имеем прекрасные примеры Рекапитуляции анцестральных состояний, известные всем как Классические иллюстрации биогенетического закона. Укажу на закладку кардинальных вен и кювьеровых протоков у всех позвоночных, на закладку артериальных дуг и их преобразований у ааземных позвоночных и т. д.
80 Организм как целое в индивидуальном и историческом развитии
Отмечу также, что основные теории происхождения головы и конечностей позвоночных построены главным образом на данных но эмбриональному развитию мускулатуры и нервной системы и в меньшей мере на основании развития скелета. Скелет, наиболее консервативная система взрослых животных, на которой в
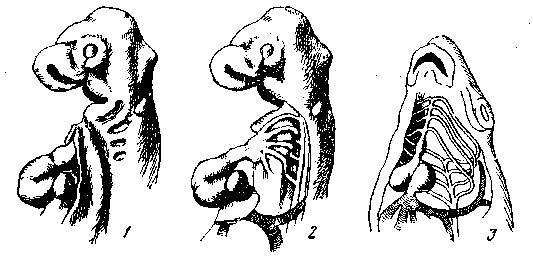
Рис. 7. Жаберные щели и жаберные дуги с артериями у куриного зародыша (1, 2) как рекапитуляция предкового состояния
Для сравнения показаны жаберные сосуды зародыша рыбы (5)
основном построена сравнительная анатомия и филогения позвоночных, дает нам часто гораздо менее ясные картины рекапитуляции.
Отчетливая рекапитуляция наблюдается и в развитии выделительной системы и в ее преобразованиях в связи с развитием половой системы. И здесь имеются далеко идущие зависимости с полостью тела, с кровеносной системой и обеих названных систем между собой.
Таким образом, становится довольно ясным, что явления эмбрионального сходства ранних стадий, а также явления рекапитуляции объясняются регуляторным характером корреляций, в особенности на ранних стадиях развития, с одной стороны, и трудностью перестройки системы взаимозависимостей между морфо-генетическими процессами без коренного нарушения самого эмбриогенеза с его летальными или полулетальными последствиями,— с другой.
Все сказанное об основах зародышевого сходства и явлений рекапитуляции можно пояснить следующими схемами, которые покажут нам также несколько яснее существование принципиального различия между обеими категориями явлений (рис. 8).
1. Явления зародышевого сходства основаны на большей изменяемости поздних стадий развития по сравнению с ранними. «Консервативность» ранних стадий объясняется сложностью системы морфогенетических корреляций регуляторного характера. Благодаря этому малые мутации на ранних стадиях развития не
Рла'ва И. Эмбри'огеНёЗ 81
получают видимого выражения, а более значительные мутации, выходящие за пределы пороговых уровней, приводят на ранних стадиях к слишком большим нарушениям развития и рано или поздно к элиминации таких особей. Поэтому поздние стадии онтогенеза изменяются в процессе эволюции быстрее, чем ранние.
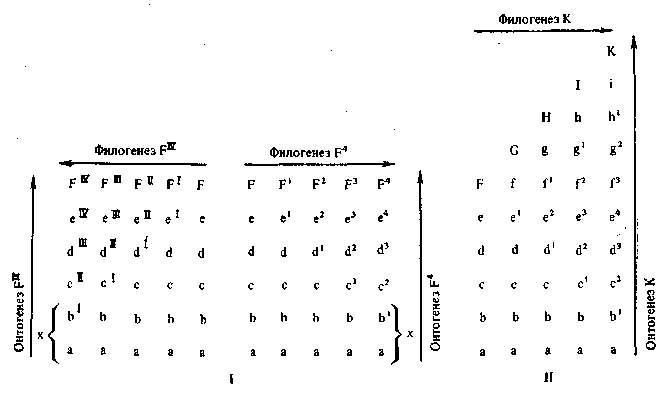
Рис. 8. Схемы явлений зародышевого сходства (I) и рекапитуляции (II)
2. Явления рекапитуляции предковых стадий основаны на преобладающем значении надставок (анаболии) в процессе эволюции, сопровождающемся установлением новых дифференцировок. Преобладающее значение надставок объясняется также сложностью системы морфогенетических корреляций, не пускающей ее нарушения внесением в нее чуждых, еще новых, не достаточно «освоенных» элементов. Поэтому новые признаки, а следовательно,, и новые взаимозависимости (как внутренние факторы их развития), вносятся, как правило, главным образом после уже законченного морфогенеза. Онтогенез усложняется прибавлением новых стадий.
Дефинитивные фазы развития предков: F, G, Н, /, К повторяются в онтогенезе потомка /3, gz, hl, i, k. Повторение это означает именно историческую последовательность в фазах развития. Ранние стадии развития оказываются при этом более измененными по сравнению с последними (степень уклонения обозначена на диаграмме цифровыми показателями). Сходство онтогенетических стадий (малые буквы) с дефинитивными стадиями предков (большие буквы), конечно, имеет лишь весьма общий характер. [При этом мы должны еще пояснить, что под дефини-
82 Организм пак целбе в индивидуальном и историческом развитии
тивными фазами мы понимаем конечные стадии морфогенеза, а вовсе не взрослые состояния предков, так как при этом в онтогенезе потомков обнаруживается сходство лишь с онтогенезом предков, а не со взрослыми формами, то некоторые современные авторы предпочитают употреблять вместо выражения рекапитуляция термин репетиции (de Beer) ].
Мы здесь не даем исчерпывающего разбора проблемы соотношений между онтогенезом и филогенезом. Нашей задачей было лишь показать значение целостности эмбрионального организма для процесса эволюции самого эмбриона. Однако наше изложение было бы все же несколько односторонним, если бы мы здесь не упомянули о возможности некоторой рекапитуляции и при мозаичном развитии. Эта рекапитуляция возможна, во-первых, конечно, потому, что мозаичность развития никогда не бывает абсолютной и что элементы регуляции как выражения существования системы взаимозависимостей всегда имеются. Процессы детерминации протекают лишь с большей скоростью и на более ранних стадиях.
Однако, кроме того, рекапитуляция возможна и в отдельных автономных органогенезах в тех случаях, когда имеется тесная зависимость между предыдущими и последующими фазами развития, т. е. когда последующая фаза может быть осуществлена только на базе предыдущей. Так, например, «рекапитуляция» соединительнотканной, хрящевой и костной стадий развития скелета позвоночных есть выражение такой последовательной зависимости: хрящ может развиться только из мезенхимы, за ее счет, а кость внутреннего скелета («первичная») формируется на базе хрящевого скелета. Последняя связь, правда, иногда разрывается (первичная кость иногда, например, у некоторых костистых рыб, развивается без хрящевой базы), но вообще это одна из обычных и ясных рекапитуляции в развитии позвоночных. Я на этих явлениях не останавливаюсь более подробно, так как на существование таких рекапитуляции уже неоднократно обращалось внимание (особенно О. Гертвиг [Hertwig, 1906], который таким образом объясняет существование рекапитуляции вообще; мы считаем для большинства случаев такое объяснение недостаточным) .
Основные закономерности филогенетических преобразований онтогенеза определяются целостностью организма на всех стадиях развития и в особенности большим значением системы мор-фогенетических связей (корреляций) на ранних стадиях эмбриогенеза. Нарушение этой целостности в известной части организма всегда приводит только к одному результату — к недоразвитию. В большинстве случаев наследственные нарушения развития (мутации) являются простыми сдвигами во времени наступления известных реакций. Явления эмбрионального сходства и «консерватизм» ранних стадий развития объясняются регуляторным ха-
Глава II. Эмбриогенез 83
рактером и трудностью перестройки системы взаимозависимостей между основными морфогенетическими процессами без коренного нарушения самого эмбриогенеза с его летальными или полулетальными последствиями. Тем же самым объясняются и явления рекапитуляции (рис. 9). Так как изменения конечных стадий любого органогенеза вносят наименьшие нарушения во все другие
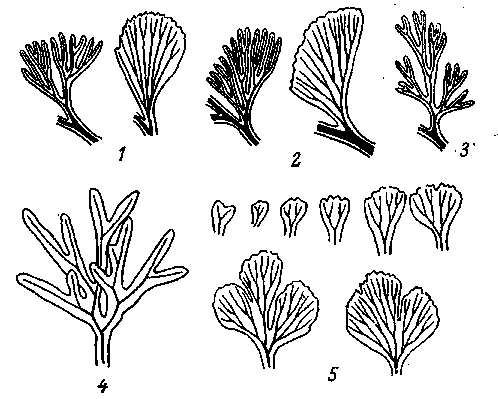
Рис. 9. Рекапитуляция у растений [Циммерманн, 1938]
Верхний ряд — листья палеозойских папоротников с дихотомическим ветвлением нервов и листовой пластинки; внизу — первичные листья современных папоротников: 1 — Archeoopteris; 2 — Rhacopteris; 3 — Sphenopteridium; 4 — Todea superba; 5 — Aneimia adiantifolia
связанные с ним морфологические процессы, то наибольшее значение в филогенетических преобразованиях эмбриона приобретают анаболии. Результатом же эволюции путем анаболии является рекапитуляция анцестральных состояний. Эти рекапитуляции будут наиболее полными у тех животных и в тех системах органов, в которых морфогенетические зависимости достигают наибольшей сложности. В автономных органогеиезах при «мозаичном» типе развития рекапитуляции никогда не бывают такими яркими, как при зависимых оргаыогенезах «регуляционного» типа, хотя и в этом случае они иногда возможны.
[Наконец, нельзя не отметить, что если в известных случаях конкретные адаптивные модификации приобретают постоянное значение и если на этой почве устанавливаются новые организационные признаки и дифференцировки, то это происходит всегда У молодого животного, ведущего уже самостоятельный образ
84 Организм как целое в индивидуальном и историческом развитии
жизни. При автономизации развития таких ддфференцировок они должны возникать заранее, т. е. на поздних стадиях морфогенеза. И в этом случае имеются все предпосылки для будущей рекапитуляции таких признаков в онтогенезе потомков].
Таким образом, мы рассматриваем явления рекапитуляции и известную консервативность ранних стадий развития как результат существования сложной корреляционной системы регулятор-ного характера, не допускающей заметных сдвигов и разрывов без нарушения жизнеспособности организма. Однако корреляционные механизмы связывают отдельные части организма в одно целое не только на ранних стадиях развития. В иной, может быть, форме, но они имеются и на поздних стадиях. Здесь их значение лишь не так резко бросается в глаза. Мы можем поэтому сказать, что наличие корреляционных систем вообще обусловливает известную стойкость организации. Это в корне меняет все наши представления. Еще недавно казалось, что стойкость организации «объясняется» стойкостью наследственной субстанции и в конечном счете прочностью (или даже «неизменяемостью») генов. Теперь даяе у генетиков звучат уже другие ноты. По работам Харланда [1933, 1937], наследственный механизм перестраивается легче, чем строение организма, и гомологичные — и притом весьма сходные — органы развиваются иногда на разной наследственной основе. При наличии сложного корреляционного механизма развития (регуляторного типа) организм в целом может оказаться более стойким, чем то, что принято называть его наследственной основой, т. е. чем генотип. Стойкость организма объясняется именно сложностью системы связей, объединяющих все его части в одно целое. Всякое значительное нарушение системы связей означает дезинтеграцию и имеет своим результатом, по меньшей мере, снижение жизнеспособности. В процессе борьбы за существование такие особи немедленно уничтожаются. Естественный отбор автоматически поддерживает существование сложных корреляционных механизмов, объединяющих развивающийся организм в одно относительно стойкое целое. В дальнейшем мы увидим, что естественный отбор не только поддерживает существование таких систем, но и создает их в процессе постепенной эволюции организмов.
Глава III. Регресс. Распад корреляционных систем в эволюции 85
Глава III
РЕГРЕСС.
РАСПАД КОРРЕЛЯЦИОННЫХ СИСТЕМ
В ЭВОЛЮЦИИ ЖИВОТНОГО ОРГАНИЗМА
Значение корреляционных систем в индивидуальном развитии изучается путем эксперимента, и в этом направлении механика, или динамика развития, имеет уже большие достижения. Нас, однако, интересует здесь другой вопрос: каково значение корреляционных систем в филогенетическом развитии, в эволюции? Может показаться странным, почему я говорю о значении корреляций в эволюции, а не о значении координации, которые ведь как раз выражают зависимости в филогенезе. Не отрицая большого значения координации в эволюции, этот вопрос мы разбираем в другой работе, специально посвященной закономерностям эволюционного процесса [Шмальгаузен, 1939—1940],—я все же считаю весьма важным обратить внимание на значение индивидуального развития и индивидуальных корреляций в эволюционном процессе. Этот вопрос до известной степени сходен с не менее актуальным вопросом о значении фенотипа в эволюции.
Проанализировать интегрирующее значение корреляционных систем можно, конечно, лишь путем их расчленения. Однако мы не можем экспериментировать с историческими процессами. И хотя мы можем сделать многие весьма вероятные выводы на основании изучения роли корреляций в индивидуальном развитии, все же эти выводы нуждаются в проверке, так как мы не можем быть уверены, что эта роль не меняется в процессах исторической перестройки организма. Наша задача облегчается, однако, тем, что мы знаем случаи дезинтеграции, т. е. распада корреляционных систем во время исторического развития организмов, и как будто довольно ясно видим даже основные факторы, ответственные за этот распад.