
И. И. Ш м альгаузен избранные труды организм как целое в и ндивидуальном и и сторическом издательство «наука» москва 1982 развитии
| Вид материала | Книга |
- Сергей Лычагин «Макиавелли Н. Избранные произведения.», 922.62kb.
- И. И. Веселовског о издательство "наука" Москва 1967 Эта книга, 1700kb.
- Избранные научные труды. М.: Наука, 1988, с. 722-734, 363.13kb.
- А. Н. Леонтьев Избранные психологические произведения, 6448.08kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 206.83kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 238.78kb.
- Электронная библиотека студента Православного Гуманитарного Университета, 208.81kb.
- Д. Н. Мамине-Сибиряке Книга, 262.07kb.
- Литература Особенности типологического подхода и метода исследования личности, 2747.84kb.
- А. Н. Леонтьев Избранные психологические произведения, 6931kb.
342 Регуляция формообразования в индивидуальном развитии
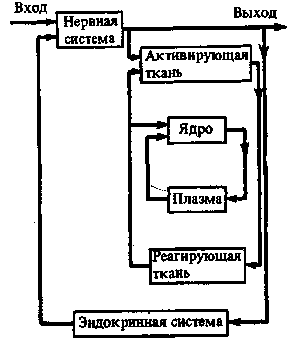
Рис. 37. Схема сильно упрощенной трехконтурной системы взаимозависимостей в развивающемся организме
Обратные связи во всех контурах изображены слева
даваемых прямой и обратной информации. Это, в свою очередь определяется строением системы. Однако, кроме того, надежность информации определяется и ее повторностью (при которой теряются «ошибки»). В организме элементарная обратная информация поступает от клетки. Число однородных клеток в любой ткани какого-либо органа буквально неисчислимо. Это означает столь же неисчислимую повторность клеточной информации. Ее надежность становится безупречной. Маломощная и ненадежная клеточная информация преобразуется, следовательно, во вполне достоверную тканевую информацию данного органа. Таким образом, высокая регу-ляторная способность высших организмов покоится, между прочим, на их высокой многоклеточности. Тканевая информация и информация, поступающая от органов, представляют собой интегральный эффект от множества клеточных информации в данной ситуации органа. Регуляция всего организма в целом обеспечивается последовательной интеграцией клеточных взаимодействий во всех тканях и органах. Организмы развиваются по унаследованной программе, реализация которой происходит в системе ядро — плазма в зависимости от ситуации каждой клетки и синтезируется в тканевых системах и органах соответственно положению и связям данных частей. Все частные взаимозависимости охватываются общей системой регуляции всего организма в целом. Последняя определяется в значительной мере положением организма во внешней среде.
Высокая регуляторная способность организма не исключает, конечно, возможности нарушений формообразования в индивидуальном развитии. Эти нарушения могут быть результатом изменений в унаследованной программе, т. е. системе ядро — цитоплазма. Особое значение имеют изменения в ядерных структурах. Они реализуются в виде наследственных мутаций. Возможны нарушения и во внутренних связях — они также приводят к изменениям формообразования и к тем или иным уродствам (как это видно в опытах экспериментального вмешательства в соотношения частей). Возможно также искусственная изоляция. частей, например в эксплантате. Это приводит обычно к более или менее значительному недоразвитию, а затем и к остановке в раз-
Рлава X. Регуляция формообразования и устойчиводтъ Ьрганизмов 343
витии. Нормальные связи являются необходимыми факторами формообразования.
Если, однако, обеспечить достаточное питание изолированной части, то возможен, очевидно, ее неограниченный рост, как это видно на тканевых культурах. По-видимому, это возможно и в организме при нарушениях формативных связей. Так, по данным А. Н. Студитского, при реимплантации разрушенной мышечной ткани на нормальное место из нее развивалась соответствующая мышца. Когда, однако, такая разрушенная мышца изолировалась целлофаном от ближайшего окружения при сохранении нервных и механических связей, оформленная мышца не развивалась, а мышечная ткань перерождалась в опухоль. В этом случае были нарушены только формативные связи с соседними тканями при изоляции целлофаном. В организмах, конечно, этого не бывает, однако, вполне возможна биохимическая изоляция известной части в результате коренной перестройки ее метаболизма, приводящая к ее бесконтрольному, анархическому росту, т. е. к возникновению опухоли.
Перестройка метаболизма возможна под различными влияниями. Обычно она не выходит за пределы унаследованной нормы реакций. Особенно велики, однако, изменения, связанные с преобразованиями ядерных структур— унаследованной ДНК и построенной соответственно РНК. Это возможно, между прочим, в следующих случаях.
- Заражение группы клеток вирусом, который вносит свою дезоксирибонуклеиновую кислоту в зараженные клетки, или вирусом, РНК которого вытесняет рибонуклеиновую кислоту в цитоплазме тех же клеток. В обоих случаях метаболизм зараженных клеток меняется в корне — специфический метаболизм клеток данного организма замещается метаболизмом самого вируса. Это может нарушить воспроизведение собственной ДНК, т. е. вызвать мутацию. Продукты вирусного метаболизма не могут заменить продуктов клеточного и тканевого обмена данного организма. Если эти продукты были носителями специфической информации, передаваемой соседним тканям, то продукты вирусного обмена не могут играть подобной роли — как совершенно чуждые субстанции они не могут вызвать специфического ответа в нормальных тканях. Компетенция последних ограничена нормой реакций только на свои специфические нормальные раздражители. Таким образом,' обратная информация от зараженных клеток оказывается глубоко нарушенной, контроль формообразования со стороны окружающих частей невозможен.
- Возникновение соматической мутации (под влиянием вируса или спонтанно), связанной с глубокой перестройкой метаболизма, может в известных случаях привести к такому же результату. Искажение обратной информации может также достигнуть той степени, когда она становится недоступной («непонятной»)
344 Регуляция формообразования в индивидуальном развитии
для окружающих нормальных тканей, а, следовательно, не способной вызвать и ответного формативного воздействия.
Мутации возникают с большей частотой, и притом повторно, в сильно пролиферирующих тканях. При возникновении разных мутаций, отличающихся интенсивностью своего обмена и скоростью пролиферации, неизбежно вытеснение малоактивных клеточных элементов более активными, с более интенсивным ростом. В результате некоторые мутанты получают преобладание и дают начало особой ткани с быстрым ростом. Вследствие нарушения обратной информации и невозможности формативных влияний (чему способствует и утрата компетенции к специфическим реакциям) эта ткань не может дифференцироваться и ее рост становится неограниченным (при достаточном ее питании).
3. Возникновение соматической мутации, связанной с разрывами, перестройками и переносом целых хромосом. В этих случаях возникают многие летальные комбинации, которые сразу исключаются из ткани. Возможны и очень многие комбинации с пониженным обменом — они рано или поздно вытесняются более активными клетками. Встречаются, однако, и такие комбинации, которые не обеспечивают равнонаследственного деления ядра. Возникают также многополюсные митозы с совершенно неправильным распределением хромосом и т. п. В этих случаях образуется конгломерат клеток, которые все отличаются друг от друга по своим наследственным структурам (ДНК). Если говорить о клеточной информации, то она здесь не только в корне искажена (вследствие глубокого изменения метаболизма), но, кроме того, совершенно различна для разных клеток. Здесь нет той повторности, которая делает нормальную тканевую информацию столь надежной. Интеграция клеточной информации не возможна. Нет ни системной организации, ни повторности. Обратная информация от" получившегося клеточного конгломерата не только искажена, но и полностью дезорганизована. Без обратной информации, однако, невозможны ни контроль, ни регуляция. Такие клеточные конгломераты способны лишь к анархическому, совершенно бесконтрольному росту. Регуляция невозможна вследствие полного разрушения тех взаимозависимостей, на которых эта регуляция основывается.
Мы коснулись только двух наиболее распространенных теорий опухолевого роста — вирусной и мутационной. Я старался показать возможные источники физиологической изоляции группы клеток, которая при отсутствии обратной связи выходит из-под контроля со стороны основных частей организма. Возможны и другие гипотезы возникновения опухолевого роста. Все же основной причиной мне представляется нарушение нормальных формативных связей. Однако, таким нарушением можно объяснить лишь рост опухоли, который по возобновлении связей должен прекратиться, после чего эта ткань может дать начало различным
Литература 345
дифференцировкам. Злокачественное же перерождение клеток не может быть объяснено одной только физиологической изоляцией. Поэтому все гипотезы возникновения опухолей должны быть дополнены объяснением причин их малигнизации, которая, по-видимому, наступает при глубокой перестройке контрольного аппарата ядра в результате последовательного ряда мутаций.
На приведенных выше примерах мы подошли к нашей основной проблеме — регуляции формообразования — так сказать, с обратной стороны, рассмотрев возможные результаты полного нарушения аппарата регуляции. Подведем краткие итоги нашему рассмотрению проблемы регуляции в целом.
Нормальное формообразование есть процесс, строго регулируемый на всех уровнях организации — клеточном, тканевом и органотипическом. На первых стадиях развития формообразование определяется организацией зиготы, унаследованной от обоих родителей, и в особенности организаций яйцеклетки. Все позднейшие процессы определяются как перераспределением субстанций зиготы, так и установлением новых взаимодействий с внешней средой, в особенности последовательными взаимодействиями частей самого зародыша. Такие взаимодействия меняют характер метаболизма. Продукты последнего активируют или подавляют синтетическую деятельность известных участков наследственного кода хромосом. Активные участки хромосом взаимодействуют с цитоплазмой, и это вновь ведет к дальнейшей перестройке метаболизма.
Во всех взаимодействиях одна часть оказывается более активной и может быть условно названа индуктором, или активатором, другая — менее активной и может быть названа реактором. Однако результаты взаимодействия определяются не только индуцирующим влиянием, но и компетенцией реагирующей части; нередко обнаруживается и обратное действие реактора на индуктор. Формообразовательные системы имеют характер замкнутых циклов. Обратные связи, к сожалению, еще очень мало изучены, и их выявление составляет в настоящее время наиболее актуальную задачу эмбриологии. Все регулирующие циклы связаны с внешней средой посредством входных и выходных каналов. Они образуют все вместе сложную взаимосвязанную и иерархически соподчиненную систему авторегуляции формообразования всего организма в целом.
ЛИТЕРАТУРА
Астауров Б. Л. Значение опытов по мерогонии и андрогенезу для теории развития и наследственности.— Усп. совр. биол., 1948, т. 25, с. 49—88.
Астауров Б. Л., Острякова-Варшавер В. П. Получение полного гетеросперм-ного андрогенеза у межвидовых гибридов шелковичного червя.— Изв. АН СССР, серия биол., 1957, № 2, с. 154—175.
Балинский Б. И. Развитие зародыша. Проблема детерминации в эмбриональном развитии. М.; Л.: Биомедгиз, 1936,
346 Регуляция формообразования в индивидуальном развитии
Враше Ж. Биохимическая эмбриология. М.: ИЛ, 1961.
Вильсон Э. Клетка и ее роль в развитии и наследственности, т. I. M.; Л.: Биомедгиз, 1936.
Вильсон Э. Клетка и ее роль в развитии и наследственности, т. II. М.; Л 1940.
Гексли Дж. С, де Вер Г. Р. Основы экспериментальной эмбриологии. М.; Л.: Биомедгиз, 1936.
Геммерлинг И. Морфогенетические и генетические основы формообразования у зонтичной водоросли Acetabularia.— Усп. совр. биол., 1935, т. 4 с. 229—239.
Детлаф Т. А. Нейруляция у бесхвостых амфибий как комплексный формообразовательный процесс— Труды Ин-та морфогенеза, 1938, т. 6, с. 187— 200.
Драгомиров Н. И. Корреляция в развитии эктодермальных зачатков глаза.— Изв. АН СССР, серия биол., 1939, № 5, с. 741—768.
Завадовский М. М. Пол и развитие его признаков. К анализу формообразования у животных. М.; Госиздат, 1922.
Завадовский М. М. Противоречивое взаимодействие между органами в теле развивающегося животного. М.: МГУ, 1941.
Лопашов Г. В. Механизмы развития зачатков глаз в эмбриогенезе позвоночных. М.: Изд-во АН СССР, 1960.
Машковцев А. А. Саянский дикий реликтовый северный олень.— Докл. АН СССР, 1940, т. 27, с. 78-80.
Морган Т. Г. Экспериментальная зоология. М.: Типография торг. дома Мош-кина и Ге, 1909.
Нейфах А. А. Проблема взаимоотношений ядра и цитоплазмы в развитии. М.: Изд-во АН СССР, 1962.
Полежаев Л. В. Основы механики развития позвоночных. М.; Л.: Изд-во АН СССР, 1945.
Полетаев И. А. Сигнал. О некоторых понятиях кибернетики. М.: Сов. радио, 1958.
Строева О. Г., Никитина А. А. Пересадка ядер у амфибий и ее значение для исследования проблемы дифференцировки.— Журн. общ. биол., 1960, т. 21, с. 336-346.
Студитский А. Н. Некоторые вопросы биологической теории рака.— Журн. общ. биол., 1962, т. 23, с. 176—192.
Тойвонен С. Первичная эмбриональная индукция у амфибий.— Успехи совр. биол., 1963, т. 55, № 1, с. 87.
Филатов Д. П. Значение фактора объема в ускорении некоторых морфогенезов.— Журн. экспер. биол., 1931, т. 7, с. 137—162.
Хеммонд П. Теория обратной связи и ее применения. М., 1961.
Шмалъгаузен И. И. Организм как целое в индивидуальном и историческом развитии. М.; Л.: Изд-во АН СССР, 1938, 1942.
Шмалъгаузен И. И. Проблема устойчивости органических форм (онтогене-зов) в их историческом развитии.— Журн. общ. биол., 1945, т. 6, с. 3—25.
Шмалъгаузен И. И. Факторы эволюции. М.; Л.: Изд-во АН СССР, 1946.
Шмалъгаузен И. И. Интеграция биологических систем и их саморегуляция.— Бюлл. МОИП, отд. биол., 1961, т. 66, с. 104—134.
Child С. М. Patterns and problems of development. Chicago, 1941.
Dalcq A. M. Germinal organization and induction phenomena.— In: Fundamental aspects of normal and malignant growth/Ed. W. W. Nowinski. Amsterdam, 1960.
Dobzhansky Th. Genetics and the origin of species. N. Y., 1951.
Dobzhansky Th. Evolution, genetics and man. N. Y.; London, 1955.
Dragomirow N. [Драгомиров Н.\ Uber die Faktoren der embryonalen Entwick-lung der Linse bei Amphibien.— Roux' Arch. Entwicklungsmech. Organis-men, 1929, Bd. 116, S. 633-668.
Durken B. Lehrbuch der Experimentalzoologie. Berlin, 1928.
1)цгкец В. Grundriss der Entwicklungsmechanik. Berlin, 1929,
Литература 347
Qoldschmidt R. Physiological genetics. N. Y.; L., 1938.
jtiihn A. Vorlesungen iiber Entwicklungsphysiologie. Berlin, Gottingen, Heidelberg, 1955.
Mangold 0. Das Determinationsproblem I. Das Nervensystem und die Sinnesor-gane der Seitenlinie unter spezieller Beriicksichtigung der Amphibien.— Ergebn. Biol., 1928, Bd. 3, S. 152—227.
Mangold 0. Das Determinationsproblem. II. Die paarigen Extremitaten der Wirbeltiere in der Entwicklung.— Ergebn. Biol., 1929, Bd. 5, S. 290—404.
Mangold 0. Das Determinationsproblem. III. Das Wirbeltierauge in der Entwicklung und Regeneration.—Ergebn. Biol., 1931, Bd. 7, S. 193—403.
Morgan Т. Н. Experimental embryology. N. Y.: Columbia Univ. Press, 1927.
Saxen L., Toivonen S. Primary embryonic induction. London, 1962.
Spemann H. Experimented Beitrage zu einer Theorie der Entwicklung. В., 1936.
Waddington С. Н. Principles of embryology. London, 1956 (1957).
Waddington С. Н. The strategy of genes. London, 1957.
Weiss P. Entwicklungsphysiologie der Tiere. Dresden; Leipzig, 1930.
СТАБИЛИЗИРУЮЩИЙ ОТБОР
И ЭВОЛЮЦИЯ
ИНДИВИДУАЛЬНОГО РАЗВИТИЯ
В понимании естественного отбора и его аналогии с искусственным в настоящее время распространены различные взгляды, находящиеся порой в резком противоречии со взглядами самого Ч. Дарвина. Одним из наиболее ясных выражений такого расхождения является обычное отрицание творческого значения естественного отбора. Последний действует, по Дарвину, через накопление в известных направлениях множества малых наследственных изменений. Мутационная теория перенесла внимание исследователей на отдельные более значительные наследственные изменения, в которых стали видеть подлинные ступени по пути эволюции органических форм. Естественному отбору отводится лишь роль решета, отбрасывающего негодные формы и сохраняющего годные. Эти последние создаются будто бы не в процессе отбора более сложных комбинаций, а уже самим процессом мутирования. Я не ставлю своей задачей в этой статье критику указанных взглядов. Упомяну только, что очень большую роль в их преодолении сыграли блестящие работы Р. Фишера (R. A. Fisher) Дж. Холдена (J. В. S. Haldane), Г. Меллера (Н. Muller) и С. Харланда (S. E. Harland).
Здесь я хочу обратить внимание на другое: 1) на общий механизм действия естественного отбора, 2) на его поддерживающее влияние в условиях установившихся уже соотношений между определенным организмом и нормальной для него средой и возможную роль в «фиксировании» результатов индивидуального приспособления, 3) на его значение в перестройке самого механизма индивидуального развития. В этой связи прежде всего коснемся вопроса о селекционном значении «малых» и «безразличных» наследственных изменений, а также и возможной роли ненаследственных изменений.
Так как естественный отбор, в отличие от современного искусственного отбора, покоится не на выборе положительных уклонений, а на элиминации отрицательных, то этим самым при учете процессов повторного мутирования первый вопрос, служивший долгое время предметом дискуссии, получает сразу положительное решение. В особенности при этом следует помнить, что естественный отбор имеет всегда дело с целостными индивидуальностями сложного происхождения, а не с отдельными мутациями, которые имеют до известной степени реальное существование
Стабилизирующий отбор и эволюция индивидуального развития 340
лишь в наших экспериментальных культурах. Этим самым получает положительное разрешение и второй вопрос — о роли ненаследственных изменений. В природных условиях каждая особь отличается от любой другой особи многими наследственными и ненаследственными особенностями, покоящимися на различных нормах реакций этих особей. Вопрос о жизни и смерти особи решается каждый раз конкретными взаимоотношениями между данной особью, на всех стадиях ее индивидуального развития, и той средой, в которой развертывалась вся ее жизнь. В борьбе за существование гибнут или сохраняются и оставляют потомство особи, различающиеся по всему своему индивидуальному развитию, по всей своей морфофизиологической организации, по своим реакциям и своему индивидуальному поведению (а не особи, отличающиеся друг от друга лишь отдельными наследственными признаками). Естественно, что особи, «переживающие» и сохраняемые в борьбе за жизнь, отличаются от элиминируемых, т. е. от гибнущих или устраняемых от размножения, всей своей организацией в целом. По своей наследственной характеристике это будут всегда сложные комбинации весьма многих и в особенности «малых» «морфологических» и «физиологических» мутаций. Такие особи могут обладать не только положительными и «безразличными», но даже и незначительными отрицательными «признаками». Конечно, это возможно лишь в том случае, если вред, вносимый отрицательным (например, морфологическим) признаком, вполне перекрывается другими, положительными (например, физиологическими) особенностями данной особи.
Отметим также как прочно установленный факт, что огромное большинство даже незначительных вновь возникающих мутаций оказываются неблагоприятными для их обладателей (что вполне понятно при учете сложности и согласованности всей организации любого животного или растения и ее приспособленности к весьма разнообразным факторам нормальной внешней среды). Если мы примем во внимание сказанное, то указания Робсона и Ричардса [Robson, Richards, 1936, с. 279] на широкую распространенность бесполезных структурных различий среди признаков, характеризующих виды и роды, в значительной мере теряют свою убедительность. Ведь требование, чтобы более 50% таких признаков имело приспособительный характер, основывается на неверной предпосылке, что в исходном материале положительные и отрицательные уклонения возникают с одинаковой частотой, и на неверном представлении о механизме действия естественного отбора, мыслимого по аналогии с сознательным искусственным отбором одних лишь положительных уклонений.
В процессе мутирования происходит совершенно свободное накопление положительных, безразличных и даже малых отрицательных мутаций, которые через постоянное скрещивание переживающих особей вступают во все новые комбинации. В этих ком-