
Отчет о научно-исследовательской работе
| Вид материала | Отчет |
- Реферат отчет о научно-исследовательской работе состоит, 61.67kb.
- Отчёт о научно-исследовательской работе за 2011 год, 1208.93kb.
- Отчёт о научно-исследовательской работе за 2009 год, 851.3kb.
- Отчёт онаучно-исследовательской работе гу нии но ур за 2010 год, 997.69kb.
- Отчет о научно-исследовательской работе профессорско-преподавательского состава, 617.56kb.
- Отчет о научно-исследовательской работе; пояснительная записка к опытно-конструкторской, 14.47kb.
- Отчет о научно-исследовательской работе (итоговый), 2484.06kb.
- Отчет о научно-исследовательской работе, 2473.27kb.
- Отчет о научно-исследовательской работе, 392.92kb.
- Задачи секции: широкое привлечение учеников к участию в научно исследовательской работе;, 67.94kb.
На основании проведённых ранее исследований были обнаружены определённые корреляционные зависимости между концентрациями и соотношениями препаратов глицин, коламин и янтарная кислота при прединкубационной обработке яиц в цехах инкубации и выращивания птичного племенного завода «Птичное», Наро-Фоминский р-н, пос. Птичное, а также в лабораториях кафедр зоогигиены им. профессора А.К. Даниловой, органической и биологической химии им. профессора С.И. Афонского и Г.А. Малахова ФГОУ ВПО МГАВМиБ.
Для более подробного изучения влияния этих препаратов на куриные эмбрионы были созданы опытная и контрольная партии из яиц, соответствующих установленным нормативам для инкубационных яиц, взятых от одного родительного стада при соблюдении равенства массы, времени снесения и срока хранения.
Таблица 22 - Показатели качества яиц.
| Масса, г | Индекс | Содержание в желтке | Индекс формы | Толщина скорлупы, мм | ||
| Белка % | Желтка, % | |||||
| Каратиноиды, мкг/г | Витамин А, мкг/г | |||||
| 65,37 | 11,6 | 0,49 | 13,0 | 6,14 | 78,3 | 0,39 |
Яйца хранили при температуре 12ºС, в течение 3 дней. Опытные партии аэрозольно из пульверизатора обрабатывали по схеме, представленной в таблице 23.
Таблица 23 - Схема опыта.
| Возраст родит. стада, дн. | Партии | Колич. залож. яиц, шт. | Обработка яиц до инкубации. концентрации препаратов коламин, глицин, янтарная кислота |
| 315 | Контрольная | 267 | ________ |
| Опытная 1 | 256 | 0,01%+0,01%+0,01% | |
| Опытная 2 | 256 | 0,05%+0,05%+0,05% | |
| Опытная 3 | 264 | 0,1%+0,1%+0,1% |
Учитывая раннее обнаруженные корреляции, были подобраны концентрации и соотношения препаратов, способствующих повышению вывода и выводимоcти эмбрионов, вследствие, снижения отходов инкубации. Действие препаратов в первую очередь по трем основным комплексам показателей: клинико-биохимическим, анатомо-морфологическим и биоконтроля инкубации.
Зооветеринарные показатели инкубации.
Таблица 24 - Показатели биоконтроля инкубации,% .
| Партия | Неоплод | Кровяные кольца | Замерзшие | Задохлики | Слабые | Вывод | ±Δ | Выводимость | ±Δ |
| Конт. | 11,55 ±1,96 | 2,62 ±0,98 | 3,06 ±1,05 | 5,62 ±1,41 | 1,87 ±0,83 | 75,28 ±2,64 | ----- | 87,40 ±2,03 | ----- |
| 1 группа | 9,77 ±1,86 | 1,96 ±0,87 | 2,73 ±1,02 | 3,13 ±1,09 | 3,9 ±1,21 | 78,9 ±2,55 | +3,62 | 87,45 ±2,07 | +0,05 |
| 2 группа | 7,03 ±1,60 | 1,95 ±0,86 | 0 | 1,95 ±0,86 | 1,95 ±0,86 | 87,89 ±2,04* | +12,61 | 94,54 ±1,42* | +7,14 |
| 3 группа | 3,79 ±1,18 | 1,90 ±0,84 | 1,14 ±0,65 | 2,27 ±0,92 | 1,89 ±0,84 | 89,02 ±1,92* | +13,74 | 92,52 ±1,62* | +5,12 |
Примечание здесь и далее: *- р<0,05; **- р<0,01; ***- р<0,001
Показатели отходов инкубации в 1-ой, 2-ой, 3-ей опытных партиях снизились соответственно по сравнению с контролем: «неоплод» на 1,78%, 4,52%, 7,76%; «кровяные кольца» на 0,06%, 0,67%, 0,72%; «замерзшие» на 0,33%, 3,06%, 1,92%; «задохлики» на 2,49%, 3,67%, 3,35%.
Количество «слабых» незначительно увеличилось во всех группах на 2,03%, 0,08%, 0,02%, что может свидетельствовать, о неустойчивости эффекта действия препаратов, при однократной обработке.
Снижение большинства показателей отходов инкубации указывает на повышение жизнеспособности эмбрионов, и подтверждается возросшими показателями вывода и выводимости. Показатели вывода в 1-ой опытной группе увеличились на 3,62% по сравнению с контролем, во 2-ой группе на 12,61%, в 3-ей на 13,74% . Показатель выводимости так же возрос во всех опытных группах: в 1-ой на 0,05%, во 2-ой на 7,14%, в 3-ей на 5,15%) соответственно по сравнению с контролем.
Из данных таблицы 3 видно, что наиболее выраженные положительное влияние на организм эмбрионов оказал комплекс препаратов в концентрациях 0,05%+0,05%+0,05% и 0,1%+0,1%+0,1%. В 1-ой опытной группе с наименьшей концентрацией 0,01%+0,01%+0,01% влияние препаратов проявилось незначительно.
Анатомо-морфологические показатели цыплят.
Спустя сутки после вывода был проведен контрольный убой птенцов, для определения степени развития внутренних органов (табл. 25).
Таблица 25 - Интерьерные показатели суточных цыплят, г.
| Показатель | Контрольная | 1 опытная | 2 опытная | 3 опытная |
| Масса цыпленка | 41,933±1,17 | 42,374±1,34 | 44,677±1,92 | 43,710±1,36 |
| Желточный мешок, с остаточным желтком | 3,745±0,14 | 3,690±0,22 | 2,730±0,24 | 2,690±0,021 |
| Печень | 1,1073±0,05 | 1,200±0,06 | 1,244±0,11 | 1,275±0,06 |
| Сердце | 0,306±0,07 | 0,315±0,08 | 0,354±0,13 | 0,335±0,11 |
| Селезенка | 0,032±0,01 | 0,031±0,02 | 0,033±0,02 | 0,033±0,02 |
| Желудок | 2,572±0,12 | 2,617±0,14 | 2,703±0,15 | 2,715±0,12 |
| Фабрициева сумка | 0,074±0,002 | 0,070±0,002 | 0,076±0,003 | 0,076±0,003 |
Из таблицы следует, что в 2-ой – 3-ей опытных группах присутствует тенденция к лучшему развитию органов по сравнению с контролем. В частности, достоверно снизилось количество остаточного желтка во 2-й и 3-й на 27,1% и 28,1%соответственно по сравнению с контролем.
Биохимические исследования.
Биохимические исследования показали, что обработка яиц растворами препаратов при определенных концентрациях оказала положительное действие на обменные процессы (табл. 26).
Таблица 26 - Клинико-биохимические показатели крови и сыворотки крови суточных цыплят.
| Группа Показатель | Контрольн. | 1 опытная | 2 опытная | 3 опытная |
| 1 | 2 | 3 | 4 | 5 |
| Общий белок, g/l | 32,3±1,42 | 33,2±1,62 | 37,2±1,16* | 39,04±1,81* |
| Альбумин, g/dl | 3,99±0,08 | 4,01±0,12 | 4,12±0,07 | 4,24±0,06 |
| Мочевая к-та, umol/l | 0,31±0,008 | 0,29±0,01 | 0,33±0,02 | 0,33±0,02 |
| 1 | 2 | 3 | 4 | 5 |
| АлАт, u/l | 7,48±0,12 | 7,66±0,35 | 8,29±0,23* | 8,19±1,17 |
| АсАт, u/l | 146,8±5,23 | 172,8±3,06* | 192,8±7,6** | 195,1±3,01* |
| α-Амилаза, u/l | 1080±92,7 | 1104±47,7 | 1330±57,57 | 1447±39,4* |
| Глюкоза, Mmol/l | 9,36±0,08 | 9,66±0,06* | 10,2±0,18* | 10,7±0,34* |
| ЛДГ, U/l | 997,6±37,2 | 1054,1±71,46 | 1344±79,03* | 1369±39,92*** |
| Общие липиды, g/l | 1,45±0,014 | 1,468±0,006 | 1,492±0,012 | 1,48±0,011 |
| Креатинин, mkmol/l | 17,7±0,76 | 18,2±0,70** | 19,4±0,75 | 19,4±0,74 |
| Са, mmol/ l | 3,18±0,06 | 3,18±0,17* | 3,55±0,05** | 3,54±0,06* |
| Р, mmol/ l | 2,03±0,07 | 2,12±0,06 | 2,33±0,03* | 2,20±0,07 |
| Щелочная фосфатаза, U/l | 1227±15,16 | 1514±18,70 | 1660±11,92* | 1622±19,03 |
| Лизоцим, mkg/ml | 35,7±0,4* | 35,4±0,8 | 39±0,5*** | 37,7±0,6 |
Во 2-й и 3-й группах отмечается интенсификация белкового и углеводно-энергитического обменов, в частности в печени, о чем свидетельствует достоверное повышение содержания ферментов переаминирования - АлАт и АсАт, а так же увеличение показателей общего белка на 15,17% и 20,87%, альбумина на 3,26% и 6,27%. Содержание мочевой кислоты выросло на 6,45%. Уровень глюкозы вырос на 8,97%, 14,3%, активность α-амилазы на 23,14%, 34%, соответственно по сравнению с контролем.
Повышение содержания общих липидов во 2-й группе на 2,9% напрямую свидетельствует о стимуляции биосинтеза липидов в организме, что связано с действием коламина.
Нужно отметить, что все вышеуказанные биохимические показатели не превышают нормативные данные.
Из данных таблицы так же видно, что лучшие и наиболее достоверные данные по показателям кальций-фосфорного обмена были зарегистрированы во 2-ой опытной группе, где содержание Са в сыворотке крови превзошло контроль на 11,6%, Р на 14,8%, активность щелочной фосфатазы возросла в 1,35 раза. Это свидетельствует о положительном влиянии препаратов на кальций-фосфорный обмен в организме цыплят, что, очевидно, способствует стимуляции формирования костяка птицы. Кальций так же принимает активное участие в нервно-мышечной возбудимости, мышечном сокращении, свертывании крови, а так же влияет на проницаемость клеточных мембран.
Из таблицы следует, что на фоне оптимизации уровня метаболизма повысилась и неспецифическая резистентность организма цыплят, за счет увеличения лизоцимной активности в сыворотке 2-ой и 3-ей опытной группе на 9,24% и 5,6%, в сравнении с контролем.
Таким образом, по данным исследований, проведённых на моделях – куриных эмбрионах – можно отметить благоприятное влияние препаратов на оптимизацию и стимуляцию белково-ферментативного, углеводно-энергетического, липидного и минерального обменов, что может быть использовано в дальнейшем при работе с разными видами животных.
- Изучение состава и свойств биологических жидкостей коров под воздействием внешних факторов.
Методами межфазной тензиометрии и биохимического анализа были изучены сыворотка крови коров разного возраста, молоко и сыворотка молока.
Исследование сыворотки крови. В онтогенезе организм животного претерпевает весьма сложные изменения. В первую очередь это касается биохимического состава и метаболического характера, протекающих в организме обменных процессов. С возрастом существенно изменяется молекулярная структура, соотношение в тканях и клетках организма различных видов биологически-активных соединений (белков, пептидов, нуклеиновых кислот, липидов и др.). Кроме того, коренные перемены наблюдаются в концентрации и структуре многих ферментативных систем и количественном содержании макро- и микроэлементов. На разных этапах онтогенеза организм характеризуется качественно иным строением. В процессе возрастного развития механизмы адаптации переводят его жизнедеятельность на новый уровень функционирования, а гомеостатические механизмы - стабилизируют достигнутое состояние. В организме животных существует целый ряд механизмов поддержания гомеостаза, которые находятся между собой в весьма сложных взаимоотношениях. В результате их взаимной обусловленности обеспечивается системный (интегральный) характер любого приспособительного акта, результатом которых являются гомеостатические константы. Причем, в процессе адаптации перестройка систем гомеостаза может осуществляться двумя путями: как изменением структуры включенных механизмов систем регулирования, так и изменением допустимого диапазона регулируемых констант.
Кровь, являясь внутренней средой организма и зоной свободного перемещения веществ, концентрация которых находится под контролем различных физиолого-биохимических механизмов, обеспечивает стабильность функционирования органов и регуляцию их функциональной активности. При этом, обладая определенным набором констант, кровь наиболее полно отражает физиолого-биохимические процессы, протекающие в организме, и в процессе онтогенеза претерпевает количественные и качественные изменения.
Исследования были проведены на физиологически здоровых животных, коровах черно-пестрой породы животноводческого комплекса ОАО Племзавод «Петровское» Люберецкого района Московской области.
Все животные были разделены на 6 групп по принципу аналогов:
1) телочки в возрасте 6 месяцев, весом 150-165 кг;
2) телочки в возрасте 12 месяцев, весом 260-280 кг;
3) телочки в возрасте 17-18 месяцев, весом 370-390 кг;
4) нетели в возрасте 22-23 месяцев, находящиеся на 5 месяце стельности,
5) лактирующие коровы (3-4 месяц лактации) в возрасте 3 лет, находящиеся на 1-2 месяце стельности,
6) лактирующие коровы (6-7 месяц лактации) в возрасте 3,5 лет, находящиеся на 5-6 месяце стельности.
В 5 и 6 группу были включены коровы массой от 530 до 580 кг. с удоем за предыдущею лактацию 5300-5400 кг с жирностью молока 3,7-3,8%.
Исследование ДПН сыворотки крови коров, находящихся на разных фазах постнатального онтогенеза позволило выявить определенные закономерности, связанные с возрастом и периодом репродуктивного цикла животных.

Рисунок 30 - Тензиограммы сыворотки крови коров.
Тензиограммы сыворотки крови коров всех исследуемых групп характеризуются относительно небольшим понижением ПН при малых временах «существования» поверхности и последующим быстрым снижением ПН при t > 0,6 c (рис. 30). Наиболее высокие значения ПН при средних и больших временах «существования» поверхности наблюдаются для телок 12-месячного возраста, а наибольшие отличия в форме самих тензиограмм - для сыворотки крови 6-месячных телочек и лактирующих коров, находящихся на 1-2 месяце стельности.
На основании полученных тензиограмм были рассчитаны значения ДПН при разных временах существования поверхности и углы наклона кривых.
Таблица 27 - Параметры ДПН сыворотки крови коров.
| группа пара- метры | тёлочки, 6 мес | тёлки, 12 мес | тёлки, 17-18 мес | нетели, (5 мес стель- ности) | коровы (1-2 мес стель- ности) | коровы (5-6 мес стель- ности) |
| σ0, мН/м | 72,07±0,17 | 73,94±0,49* | 73,34±0,35 | 73,22±0,31 | 74,39±0,41 | 73,18±0,31 |
| σ1, мН/м | 71,91±0,43 | 73,52±0,14 | 73,08±0,48 | 72,86±0,23 | 73,09±0,51 | 72,45±0,39 |
| σ2, мН/м | 66,80±0,30 | 70,11±0,42 | 69,86±0,76 | 66,10±0,48 | 64,24±0,43 | 65,62±0,37 |
| σ3, мН/м | 61,19±0,66 | 64,35±0,64 | 62,22±0,55 | 60,45±0,59 | 56,80±0,59 | 58,97±0,69 |
| λ0, мН∙м-1с-1/2 | 5,96±0,15 | 3,16±0,39 | 4,59±0,39 | 6,07±0,28 | 5,47±0,36 | 7,33±0,36 |
| λ1, мН∙м-1с1/2 | 6,12±0,23 | 5,70±0,49 | 7,34±0,38 | 6,79±0,29 | 9,14±0,35 | 7,98±0,60 |
Для всех значений Р>0,99 (p<0,01)
У 6-месячных телочек параметры ДПН сыворотки крови имеют особенности, которые с возрастом претерпевают существенные изменения. Наступление половой зрелости и достижение телочками живой массы 260-280 кг. приводит к адаптивным изменениям параметров ДПН их сыворотки крови, так, σ0, σ1, σ2 и σ3 к 12 месяцам увеличиваются на 2,6%, 2,3%, 5% и 5,2% соответственно, а значения коэффициента наклона кривой в области малых (λ0) времен существования поверхности, напротив, снижаются на 47%. Это свидетельствует о более низком содержании ПАВ в сыворотки крови этой группы животных по сравнению с телочками 6-месячного возраста. К моменту наступления физиологической зрелости (17-18 мес.) телки достигают живой массы 370-390 кг, завершается формирование их организма, что приводит к значительным изменениям значений параметров ДПН их сыворотки крови: σ3 снижается на 3,3%, а λ0 и λ1 повышаются на 48% и 28% соответственно.
Известно, что наступление беременности сопровождается многочисленными сложными изменениями в организме самок, что создает оптимальные условия для внутриутробного развития плода. Обмен веществ при беременности подвергается особенно значительным изменениям, что связано с перестройкой функций нервной системы и желез внутренней секреции. При беременности ряд содержащихся в крови биологически-активных веществ существенно изменяется по количеству и по активности, это приводит к изменениям значений ДПН сыворотки крови стельных животных. Так, к 5 месяцу стельности у нетелей и лактирующих коров значения ПН снижаются: при средних временах существования поверхности (σ2) на 5,4% и 6%, и при больших временах существования поверхности (σ3) на 2,8% и 5,8% соответственно, а значения коэффициента λ0 увеличиваются на 32% и 58% соответственно, по сравнению с телками 17-18-месячного возраста. У коров, находящихся на 1-2 месяце стельности, значения параметров σ2.…σ3 в среднем на 5%, и λ1 - на 24% ниже, а значения λ0 на 19% выше по сравнению с животными в 17-18-месяцев. Нетели и коровы на 5-6 месяце стельности имеют более высокие значения ПН (σ0 выше в среднем на 1,6%, σ2 - на 2,5%, σ3 - на 6 и 3,6% соответственно) и λ0 (выше на 10 и 25% соответственно), и более низкие значения λ1 (соответственно на 35 и 15% ниже) по сравнению с коровами, находящимися на 1-2 месяце стельности. Очевидно, это связано с более низким содержанием ПАВ в сыворотки крови животных во второй трети стельности, а у лактирующих коров с максимально высокими удоями на 3-4 месяце лактации.
В целом, максимальные значения параметров ДПН сыворотки крови коров всех исследуемых групп отмечаются в области очень малых времен «существования» поверхности (0) и, по-видимому, определяются солевым составом сыворотки крови, а минимальные - при больших временах «существования» поверхности, и определяются постепенной адсорбцией поверхностно-активных компонентов сыворотки крови на границу раздела фаз. Наиболее высокие значения параметров σ1 (73,52±0,14 мН/м), σ2 (70,11±0,42 мН/м) и σ3 (64,35±0,64 мН/м) отмечаются для телочек 12-месячного возраста, а σ0 (73,22±0,31 мН/м) и λ1 (6,79±0,29 мН∙м-1с1/2) для коров, находящихся на 1-2 месяце стельности. Для этих же групп животных отмечаются и наиболее низкие значения параметров λ0 (3,16±0,39 мН∙м-1с-1/2) и λ1 (5,70±0,49 мН∙м-1с1/2) - у телочек и σ2 (64,24±0,43 мН/м) и σ3(56,80±0,59 мН/м) - у лактирующих коров.
Следовательно, рост, развитие и физиологическая адаптация организма коров в период стельности и лактации сопровождается закономерными изменениями в системе крови, что определяет изменения значений параметров ее ДПН, связанными со степенью функциональной активности.
Биохимические показатели сыворотки крови для животных разных групп также имеют свои особенности. Так, для телочек 12-месячного возраста уровень общего белка и альбуминов сыворотке крови на 3,8% и 11,7% ниже по сравнению с телочками в возрасте 6 месяцев. В период физиологического созревания организма концентрация альбуминов в сыворотке крови телок черно-пестрой породы вновь снижается и к 17-18 месяцев достигает 28,18±0,93 г/л, что на 18,6% ниже по сравнению с телочками 6-месячного возраста, а уровень общего белка увеличивается до 70,45±0,77 г/л.
Переход от телок к нетелям и далее к лактирующим коровам сопровождается повышением концентрации общего белка и альбуминов на 4,2% 14,1%, и 13,7% и 28,5% соответственно. У коров, находящихся на 3-4 месяце лактации (1-2 месяц стельности) отмечается наиболее высокий уровень общего белка (80,37±1,18) и альбуминов (36,21±0,73), который на более поздних сроках стельности (на 7-8 месяце лактации) снижается на 2,4% и 5,4% соответственно.
Концентрация триглицеридов в сыворотке крови коров уменьшается до 17-18-месячного возраста, а затем вновь увеличивается, однако все эти изменения находится в пределах ошибки средней арифметической величины.
Таблица 28 - Биохимические показатели сыворотки крови коров.
| группа показа- тели | тёлочки, 6 мес | тёлки, 12 мес | тёлки, 17-18 мес | нетели, (5 мес стель- ности) | коровы (1-2 мес стель- ности) | коровы (5-6 мес стель- ности) |
| Общий белок, г/л | 70,58 ±0,82 | 67,90 ±0,96 | 70,45 ±0,77 | 73,42 ±0,65 | 80,37 ±1,18 | 78,46 ±0,86 |
| Альбумины, г/л | 34,66 ±0,54 | 30,59 ±0,58 | 28,18 ±0,93 | 32,03 ±0,80 | 36,21 ±0,73 | 34,26 ±0,69 |
| Триглицериды, ммоль/л | 0,31 ±0,02 | 0,28 ±0,02 | 0,26 ±0,02 | 0,27 ±0,02 | 0,30 ±0,02 | 0,29 ±0,02 |
| Общий холестерол, ммоль/л | 1,92 ±0,12 | 2,18 ±0,12 | 2,43 ±0,19 | 2,60 ±0,26 | 3,46 ±0,26 | 3,12 ±0,29 |
| Глюкоза, ммоль/л | 2,94 ±0,08 | 3,26 ±0,33 | 3,03 ±0,28 | 3,14 ±0,32 | 2,99 ±0,33 | 3,08 ±0,55 |
| Мочевина, ммоль/л | 2,82 ±0,28 | 2,96 ±0,21 | 2,99 ±0,24 | 2,74 ±0,36 | 3,37 ±0,41 | 3,20 ±0,28 |
| Общий кальций, ммоль/л | 2,56 ±0,09 | 2,72 ±0,13 | 2,60 ±0,08 | 2,67 ±0,20 | 2,32 ±0,22 | 2,48 ±0,14 |
| Фосфор неорганический, ммоль/л | 1,69 ±0,04 | 1,77 ±0,07 | 1,87 ±0,07* | 1,70 ±0,09 | 1,96 ±0,10* | 1,72 ±0,05 |
| Калий, ммоль/л | 5,44 ±0,07 | 5,81 ±0,15 | 4,98 ±0,19 | 4,51 ±0,07 | 4,65 ±0,41 | 4,36 ±0,12 |
| Натрий, ммоль/л | 132,18 ±3,15 | 146,06 ±3,61 | 151,02 ±6,45 | 148,00 ±4,23 | 142,56 ±2,69 | 138,18 ±4,80 |
| Хлорид-ионы, ммоль/л | 97,44 ±3,08 | 97,88 ±3,67 | 105,26 ±3,28 | 102,00 ±2,07 | 101,47 ±1,89 | 101,85 ±2,68 |
Для всех значений Р>0,99 (p<0,01)
Уровень общего холестерола в сыворотке крови коров с возрастом повышается, так, у телок в 17-18 месяцев он на 27% выше по сравнению с телочками 6-месячного возраста, а у нетелей на 35% и 19% выше по сравнению с телочками в возрасте 6 и 12 месяцев. Период стельности и лактации животных характеризуется наиболее высокими концентрациями общего холестерола в сыворотке их крови. Так, максимальные значения уровня общего холестерола отмечаются на 3-4 месяце лактации, что выше по сравнению с телочками в возрасте 6, 12, 17-18 месяцев и нетелями на 79%, 58%, 42% и 33% соответственно. По-видимому, это объясняется высоким уровнем жирового обмена в период стельности и в первый период лактации, когда идет мобилизация жировых ресурсов и у коров отмечаются максимальные удои.
Уровень глюкозы в сыворотке крови у коров в разные фазы онтогенеза довольно стабилен и удерживается в пределах 2,94 - 3,26 ммоль/л, наиболее высокие значения отмечаются у телочек 12-месячного возраста.
В динамике содержания уровня мочевины отмечается плавное повышение ее концентрации в крови от телочек 6-месячного возраста к телкам 17-18 месяцев (на 6,0%). У нетелей отмечаются наиболее низкий уровень мочевины (2,74±0,36), что может быть обусловлено накоплением азота, запас которого необходим развивающемуся плоду. В период лактации содержание мочевины у стельных коров увеличивается, что в определенной мере, согласуется с наиболее высокими показателями общего белка у животных этой группы. Так, в период наивысших удоев (3-4 месяц лактации) уровень мочевины в крови коров повышается на 23,0% по сравнению с нетелями и на 19,5% по сравнению с телочками 6-месячного возраста. Но, при сопоставлении концентраций остальных, исследуемых нами, показателей азотистого обмена, таких, как общий белок и альбумины, становится очевидным, что это повышение не столь значительное. Это свидетельствует о более интенсивных процессах белкового синтеза у лактирующих животных и достаточном использовании азота корма при больших затратах белка в период наивысших удоев.
Изучение минерального состава сыворотки крови коров показало, что с возрастом уровень натрия сначала повышается, достигая максимума в 17-18 месяцев (достоверно на 14,3% выше по сравнению с телочками 6-месячного возраста), а затем с переходом от телок к нетелям и далее к лактирующим коровам плавно уменьшается на 2,0% и 5,6% соответственно.
Общая активизация обмена веществ в период полового и физиологического созревания телок ведет к достоверному увеличению концентрации неорганического фосфора в сыворотки их крови, которая к 17-18-месячному возрасту достигает 1,87±0,07 ммоль/л, что достоверно на 10,7% выше по сравнения с телочками 6 месяцев. Максимальные концентрации фосфора у коров отмечаются на 3-4 месяце лактации, что достоверно на 16,0% выше по сравнения с телочками в 6-месячного возраста. У нетелей и коров во второй трети стельности содержания фосфора вновь снижается, в среднем на 12,5% по сравнению с коровами, находящимися на 1-2 месяце стельности. Это можно объяснить растущими потребностями плода и матери в фосфорных соединениях и положительным балансом фосфора во второй половине стельности. Содержание в крови коров общего кальция и хлорид-ионов мало зависит от физиологического состояния организма и колеблется в пределах: общего кальция - 2,32±0,22 - 2,72±0,13 ммоль/л; хлорид-ионов - 97,44±3,08 - 105,26±3,28 ммоль/л. Относительная стабильность концентрации этих элементов в сыворотке крови коров связана, по-видимому, с их меньшей метаболической активностью по сравнению с фосфором и так называемой «буферной» ролью кальция в организме. В целом, минимальные значения общего кальция отмечаются у коров на 1-2 месяцев стельности, а хлорид-ионов в сыворотке крови нетелей.
Из одновалентных электролитов наибольшую лабильность проявляет калий. В сыворотке крови 12-месячных телок он достоверно на 6,8% выше по сравнению с телочками 6 месяцев. Затем, к возрасту 17-18 месяцев уровень калия вновь снижается (достоверно ниже на 8,5% и на 14,3% по сравнению с телочками 6 и 12 месяцев соответственно). При переходе от телочек к нетелям концентрация калия вновь снижается достоверно на 9,4%, а у коров на 1-2 месяце стельности вновь увеличивается на 3,1%; при дальнейшем развитии плода во второй трети стельности уровень калия вновь снижается и на 5-6 месяце его концентрация в сыворотке крови коров составляет 4,36±0,12 ммоль/л, что достоверно ниже по сравнению с телками 6, 12 и 17-18 месяцев на 19,9%, 25,0% и 12,4 % соответственно.
Таким образом, наблюдаемые изменения всех исследуемых биохимических показателей в сыворотке крови коров с возрастом находятся в пределах физиологической нормы и отражают физиолого-биохимический статус конкретной группы животных.
Уровень метаболизма организма животных определяется генетически-заложенным потенциалом, обуславливающим интенсивность и скорость протекающих в нем всех физиолого-биохимических процессов. В разные фазы постнатального онтогенеза происходит приспособление целого организма, его органов и клеток к меняющимся условиям окружающей среды. В процессе онтогенеза помимо морфологических систем и функций отдельных клеток, тканей и органов изменяется и характер обмена веществ. Рост, развитие и продуктивность животных, а также биохимические, гуморальные и другие показатели резистентности, такие, как формирование защитных сил организма и его иммунобиологической реактивности, главным образом, зависят от процессов ассимиляции и диссимиляции, в том числе и белковых структур. В свою очередь, белки крови являются важным показателем, который характеризует уровень метаболизма в организме животных в разные сроки постнатального онтогенеза. Они являются строительным материалом для клеток и тканей самого организма, а также активно участвуют в различных видах продукции.
В связи с этим изучение анализа динамики концентрации белков в сыворотке крови коров в зависимости от возраста и физиологического состояния является весьма важным в комплексной оценке их физиолого-биохимического статуса.
В значительной мере на содержание общего белка и его фракций в сыворотке крови коров оказывает структура рациона. В данной работе, животные, у которых проводилось взятие проб крови, получали рационы, представленные в табл. П1-П3. Все рационы сбалансированы по содержанию энергии и питательных веществ и соответствуют нормам кормления для крупного рогатого скота.
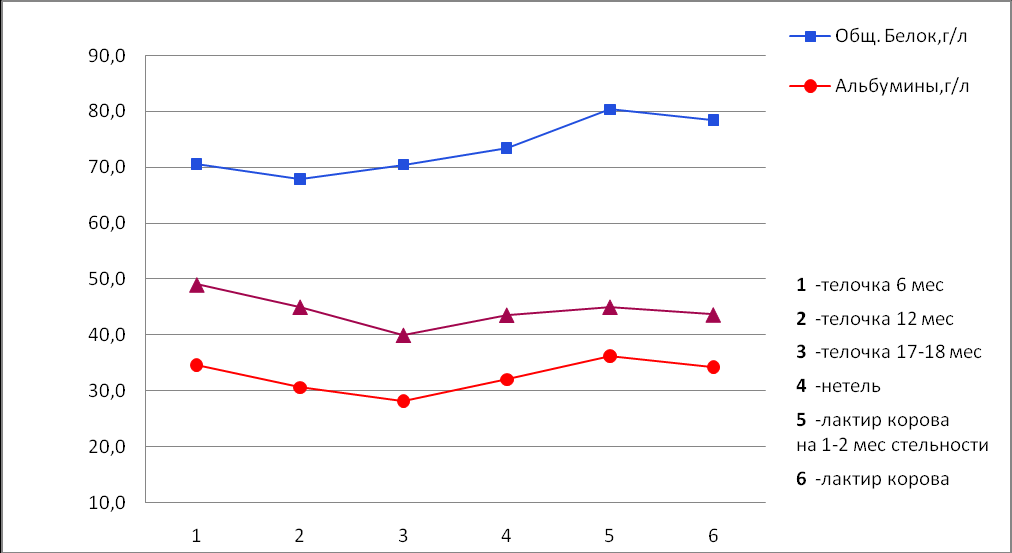
Рисунок 31 - Содержание белков в сыворотке крови коров.
В сыворотке телочек 6-месячного возраста отмечается высокое содержание альбуминов при относительно невысоком содержании общего белка. Как известно, основными белками крови являются альбумины и глобулины, следовательно, при определении концентрации общего белка и альбуминов с небольшой погрешностью мы можем говорить и об альбумин-глобулиновом соотношении (белковый коэффициент), которое у телочек в 6 месяцев равно 0,96. Это, по всей видимости, связано с тем, что в этом возрасте еще идет интенсивный процесс роста, и альбумины, в этом случае выполняют трофическую функцию и, являясь источником аминокислот, участвуют в синтезе тканевого белка. Увеличение отложения белка в тканях достигается за счёт его повышенного синтеза. Высокое значение белкового коэффициента у 6-месячных телочек свидетельствует о преобладании в их организме процессов ассимиляции, что находит своё отражение в их росте и увеличении живой массы. С наступлением физиологической зрелости количество альбуминов в сыворотке крови телок достоверно снижается (p<0,001) при одновременном снижении концентрации общего белка. Так, у 12-месячных телок белковый коэффициент составляет 0,82. После физиологического созревания в период плодотворного осеменения, процентное содержание альбумина в сыворотке крови телок снижается до 40,0% а уровень общего белка возрастает на 3,8%. С наступлением и развитием стельности концентрация альбуминов в их сыворотке крови телок достоверно повышается, одновременно увеличиваются и значения белкового коэффициента, который на 5-6 месяце стельности у нетелей равен 0,77. Интересно отметить, что эти же значения белкового коэффициента на тех сроках стельности отмечаются и у лактирующих коров в возрасте 3,5 лет, хотя концентрация в их крови общего белка и альбуминов достоверно выше, чем у нетелей. Увеличение уровня общего белка в сыворотке крови, по-видимому, связано с усилением белкового обмена веществ в организме коров во время лактации. В начальные сроки стельности в разгар лактации у коров отмечается наиболее высокий уровень общего белка (80,37±1,18), однако при этом значение белкового коэффициента (0,82) и процентного содержания альбуминов (45% от уровня общего белка) у них оказывается равным телочкам 12-месячного возраста. Относительно высокий процент альбуминовой фракции в сыворотке крови стельных животных объясняется тем, что альбумины, являясь наиболее подвижной мелкодисперсной фракцией, играют ведущую роль в синтетических процессах в организме в период беременности.
Таким образом, динамика белков сыворотки крови коров закономерно отражает физиологические процессы, происходящие в их организме в период полового и физиологического созревания и в период функциональной зрелости.