
Луценко виктор Константинович-кандидат биологических наук, старший научный сотрудник нии общей патологии и патологической физиологии амн СССР. Занимается исследованием молекулярных механизмов патологии синапса
| Вид материала | Исследование |
- Программа по патологической физиологии кафедра общей патологии и патологической физиологии, 784.63kb.
- М. Г. Масевич доктор юридических наук, профессор Российского университета Дружбы народов,, 3719.62kb.
- Васенева Клавдия Георгиевна кандидат экономических наук, старший научный сотрудник,, 845.96kb.
- А. Ж. Макашева викторов И. С., главный научный сотрудник нии проблем укрепления закон, 169.47kb.
- О. К. Степанова понятие "интеллигенция": судьба в символическом пространстве и во времени, 245.26kb.
- П ii-12-77 строительные нормы и правила защита от шума, 4029.14kb.
- Номер: за 1994 год, 300.09kb.
- Хомик под редакцией О. Ю. Артемовой Художник, 5123.02kb.
- В. С. Лукашенко главный научный сотрудник, доктор с Х. наук А. Ш. Кавтарашвили ведущий, 487.31kb.
- Магнитоакустическая эмиссия магнетитовых и титаномагнетитовых руд железорудных месторождений, 290.93kb.
ЛУЦЕНКО Виктор Константинович—кандидат биологических наук, старший научный сотрудник НИИ общей патологии и патологической физиологии АМН СССР. Занимается исследованием молекулярных механизмов патологии синапса. Автор свыше 60 научных работ.
Рецензент: Ч е п у р н о в С, А, — кандидат биологических
наук,
Луценко В. К.
Л87 Пептиды и синапсы (Механизмы регуляции деятельности центральной нервной системы).—М.;
Знание, 1986.—64 с.—(Новое в жизни, науке, технике. Сер. «Медицина»; № 1).
11-к.
В брошюра рассказано о пептидах (новом классе химических сиг-валов нервной системы) и новых представлениях о природе синап-| тической передачи. Приведены данные о происхождении, классификация в функции пептндов, об участии пептидов в процессах памяти, восприятии боли, генетическом контроле поведения и роли в асимметрии функций мозга.
Брошюра рассчитана ва медицинских работников и студентов.
4118000000
ББК 56.1 617.9
Издательство «Знание», 1986 г.
ВВЕДЕНИЕ
«Если учесть, что нынешние концепций нейрофизиологии, нейрологии и нейропсихиатрии сложились без учета этих данных, то можно думать, что будущее развитие этой области исследования окажется революционизирующим». Эти слова лауреата Нобелевской премии Р. Гиллемина относятся к открытию нейропептидов— нового класса биорегуляторов нервной системы. Чем же объясняется столь высокая оценка роли пептидов в деятельности мозга?
Известно, что восприятие, хранение и воспроизведение информации основано на химических процессах. В частности, передача возбуждения от нейрона к нейрону осуществляется с помощью химического посредника (нейромедиатора), высвобождаемого нервным окончанием в месте контакта его с поверхностью другой клетки. Область контакта двух нейронов называется синапсом. Некогда еретическая идея химической сигнализации в наше время превратилась в стройную теорию си-наптических процессов, вносящую солидный вклад в понимание работы мозга. Стало ясно, что фундаментальные процессы деятельности мозга—возбуждение и торможение, важнейшее его свойство — память, многие нервные и психические болезни могут быть поняты на синаптическом уровне. Осознание этого обусловило быстрый прогресс в изучении нейромедиаторов. Тем не менее список известных нейромедиаторов пополнялся довольно медленно, пока на сцене не появились нейропеп-тиды, после чего ситуация изменилась коренным образом.
В течение нескольких лет количество известных «химических сигналов» возросло в несколько раз. Просматривая сейчас многочисленные публикации по они-оидным пептидам (эндорфинам), посвященным поведенческим, физиологическим, фармакологическим, ги-
стохимическим и просто химическим аспектам их изучения, с трудом веришь, что всего лишь 10 лет назад об эндорфинах ничего не было известно.
Дело, однако, не только в том, что увеличилось число веществ, участвующих в синаптической передаче. После открытия пептидов был сделан новый шаг в понимании принципов химической сигнализации в нервной системе. Названия «пептиды памяти, боли, сна, агрессивности, насыщения, жажды и настроения, пептиды шизофрении» отражали мнение ученых о причастности пептидов к осуществлению целостных поведенческих актов, к работе важнейших функциональных блоков мозга в норме и при патологии.
Сенсационные опыты Г. Унгара с переносом навыков от обученного животного к необученному, казалось, открыли фантастические перспективы перед наукой и практикой. В самом деле, если многочисленные наследственные задатки' могут быть переданы от одной клетки к другой с помощью нуклеиновых кислот, в которых информация закодирована в последовательности нук-леотидов, почему несравненно меньшую информацию о поведении нельзя перенести с помощью пептидов—химических соединений, представляющих собой цепочки аминокислот? Вместо формулы «один ген — один фермент» будет «одна форма поведения—один пептид». Только и всего.
Предполагалось, что «пептиды памяти» превращают неэффективные синапсы в действующие, поэтому их предложили назвать коннекторами то есть веществами, объединяющими нейроны. В соответствии с математическими расчетами для осуществления всех мыслимых форм поведения нужно всего 1000 коннекторов.
Отставной урядник Войска Донского из рассказа А. П. Чехова «Письмо к ученому соседу» оценивал новое в науке однозначно: «Этого не может быть потому, что этого не может быть никогда». Мы поступим иначе:
расскажем о том, как возникают пептиды, как работает синапс и что делают пептиды в синапсе. Ознакомившись с синаптической функцией пептидов, читатель сможет составить собственное мнение о гипотезе Г. Унгара.
Переходя от синаптического уровня к поведенческому, мы сталкиваемся с невообразимым возрастанием уровня сложности: на каждом нейроне имеется несколько тысяч синапсов, а число нейронов в мозге превы
шает 1011. Понятно, что уяснение места пептидов в деятельности сложных нейрональных систем и механизма их действия на эти системы является нелегкой задачей. Встречаясь со сложными проблемами, исследователь расчленяет их на более простые, поддающиеся решению. Сведение сложного к простому (так называемый редукционистский метод) — плодотворный способ познания объективного мира. Достаточно сказать, что поиски первоначал привели к открытию атомного и субатомного строения материи, вещества наследственности—ДНК и т. д.
В своем изложении мы будем стремиться по возможности расчленить сложные вопросы на более простые. Так, память животного и человека — отнюдь не элементарное явление. Изменение эффективности си-наптических связей в нейронных сетях головного мозга происходит не изолированно, а на фоне параллельных психических процессов (внимание, эмоции, мотивации), затрудняющих понимание сути дела. Поэтому поставленные Г. Унгаром вопросы мы рассмотрим на модели «памяти спинного мозга», позволяющей исключить психические процессы из анализа.
Чувство боли заметно отличается от такого элементарного ощущения, как, например, прикосновение. Некоторые ученые считают, что боль вовсе не ощущение, а вид аффекта, другие говорят о «загадке боли». Ясно, что попытка всестороннего описания проблемы «пептя-ды и боль» на данном уровне наших знаний о боли бесплодна. По этой причине рассмотрение вклада пептидов в восприятие боли мы ограничим сенсорным нейроном и событиями на первом переключении в спинном мозге, где достижения особенно впечатляющи, а результаты исследований могут быть истолкованы однозначно.
Работа мозга зависит от наследственных факторов, регулирующих структуру и химизм нервной системы, что находит отражение в поведении животных. Тем не менее непонятно, каким образом осуществляется генетический контроль поведения человека. Иное дело — мозг низших животных. Исследования на примитивной нервной системе моллюска Aplysia, позволившие связать воедино гены, пептиды и поведение, обладают чрезвычайной убедительностью.
А. Эйнштейн говорил: «Все следует сделать простым,
насколько это возможно, но не проще». Целое не является простой суммой составных частей, в особенности когда этим целым является организм. Обнаружение пептидных рилизинг-факторов впервые позволило объяснить тесную взаимосвязь работы нервной и эндокрии-ной систем. Об этом и других эффектах пептидов на уровне целого организма также пойдет речь в нашей книжке.
НЕЙРОНЫ, СИНАПСЫ И ХИМИЧЕСКИЕ СИГНАЛЫ
ОТ ПОТЕНЦИАЛА ДО ПОТЕНЦИАЛА:
МЕДИАТОРЫ И МОДУЛЯТОРЫ
Прежде чем говорить о новых идеях, возникших после обнаружения пептидов, следует вкратце рассказать о двух типах сигнализации в нервной системе:
электрической и химической.
Генератором электрических импульсов является нервная клетка (нейрон). Эта способность нейрона обусловлена трансмембранным градиентом катионов и способностью ион-селективных каналов, встроенных в его поверхностную мембрану, временно открываться при небольшом снижении мембранного потенциала. В покое наружная мембрана имеет более высокую проницаемость для К'*", чем для На"1", и ничтожную—для внутриклеточных анионов. Вследствие этого К"1" выходит наружу, а соответствующие анионы не могут последовать за К4", что приводит к разделению зарядов и появлению на мембране потенциала, отрицательного внутри и положительного снаружи. Мембранный потенциал в покое называют по этой причине калиевым диффузионным потенциалом. При снижении мембранного потенциала до некоторого критического уровня открываются NaKaHanbi. В результате в нейрон поступают положительные ионы, что приводит к перемене знака потенциала (минус снаружи, а плюс внутри). В отличие от потенциала покоя потенциал, возникающий при активации электровозбудимых ионных каналов, носит назва< ние потенциала действия. Снижение мембранного потенциала происходит не только в данной точке, но и на некотором расстоянии от нее, что активирует соседние Na+-KaHanbi и т. д. Возбуждение распространяется по нейрону и его длинному отростку—аксону. Ь-каналы
Открываются несколько позднее, а так как градиент К противоположен градиенту Na+, ионы K+ перемещаются наружу, вызывая возвращение мембране исходного потенциала (фактически он становится даже несколько выше исходного). Специальный фермент—Na+, К+ — АТФ-аза — затем восстанавливает- исходные градиенты, откачивая Na+ наружу и закачивая К+ внутрь нейрона.
Значение электрических явлений в работе нервной системы трудно переоценить. Физическое воздействие на органы чувств преобразуется в последовательности электрических импульсов, которые с большой скоростью (иногда больше 100 м/с) достигают нервной системы, осведомляя мозг о событиях внешнего мира. Электрическая активность нейронов дыхательного центра управляет сокращением дыхательных мышц, скелетные мышцы, железы, сердце, сосуды также управляются электрическими сигналами, распространяющимися по соответствующим нервным волокнам.
Однако нервная система не составляет единого целого. Между нервным окончанием одного нейрона и поверхностью другого имеется щель, которую нервный импульс преодолеть не в состоянии. Чтобы цепь электрических явлений не была прервана, природе пришлось создать синапс — специализированное устройство для цепочки электрохимических преобразований.
Рассмотрим наиболее существенные черты химической передачи нервного импульса на примере идеализированного синапса центральной нервной системы. Нервная клетка имеет несколько отростков характерной древовидной формы, которые поэтому называются ден-дритами. Самый длинный тонкий отросток — аксон — оканчивается вздутием, которое иногда называют си-наптическим бутоном или бляшкой, а мы будем называть просто нервным окончанием. Диаметр утолщения — примерно 1 мкм. Нервное окончание соприкасается с поверхностью другого нейрона, но не сливается с ним. Ширина щели между нейронами составляет примерно 20—50 нм. Нервное окончание, щель и специализированный участок мембраны второго нейрона представляют в функциональном смысле единое целое, называемое синапсом (областью контакта). Учитывая направление распространения нервного импульса, нейрон, которому принадлежит нервное окончание, называют
рреп.паптическим (предшествующим), а второй — пост-си на пти чески м {следующим за синапсом). Эти же обозначения переносят » на мембраны соответствующих нейронов.
Нервное окончание одного нейрона может контактировать с дендритом, телом или аксоном другого. В нервном окончании синтезируется химическое вещество, выполняющее роль передатчика импульса (нейро-медиатора). Молекулы медиатора концентрируются внутри большого числа специализированных образований—синаптических пузырьков (везикул). В покое везикулы сталкиваются с пресинаптической мембраной, однако редко случайным образом сливаются с ней, еб-разуя пору, через которую медиатор диффундирует в синаптическую щель. Для запуска процесса высвобождения необходимы ионы кальция, которых в покое внутри клетки очень мало.
Рассмотрим принцип действия синапса на крайне упрощенной модели (рис. 1), предположив, что в тер-минали везикула содержит единственную молекулу медиатора. В постсинаптической мембране также условно мы разместим единственную опознающую медиатор молекулу—рецептор, сопряженную с единственным каналом, пропускающим либо Na+, либо К"1'. Итак, везикула слилась с мембраной, возникла водная пора, через которую молекула медиатора проникла в синаптическую щель и была связана рецептором. Если взаимодействие произошло по принципу ключа и замка, то есть строго специфически, рецептор претерпевает структурные, или, как говорят, конформационные, изменения, что, в свою очередь, приводит к открыванию сопряженного с ним ионного канала. Эти каналы постсинаптической мембраны в отличие от тех, которые расположены на теле нейрона или аксона, нечувствительны к измененкю мембранного потенциала.
Если ионный канал селективен для Na+, то его от-крыьание снизит мембранный потенциал мембраны под синаптическим окончанием, тогда как открытие К+-канала приведет к увеличению поляризации мембраны (гиперполяризация). Находящийся рядом с синаптиче-ской областью обычный электровозбудимый Ма+-канал при снижении мембранного потенциала до критического уровня также откроется, что приведет к возникновению и распространению потенциала действия. Следует
иметь в виду, что одна везикула содержит несколько тысяч молекул медиатора, но даже при высвобождении всей порции везикулярного содержимого (кванта медиатора) возникают только локальные синаптические потенциалы: деполяризационные (возбуждающие пост-
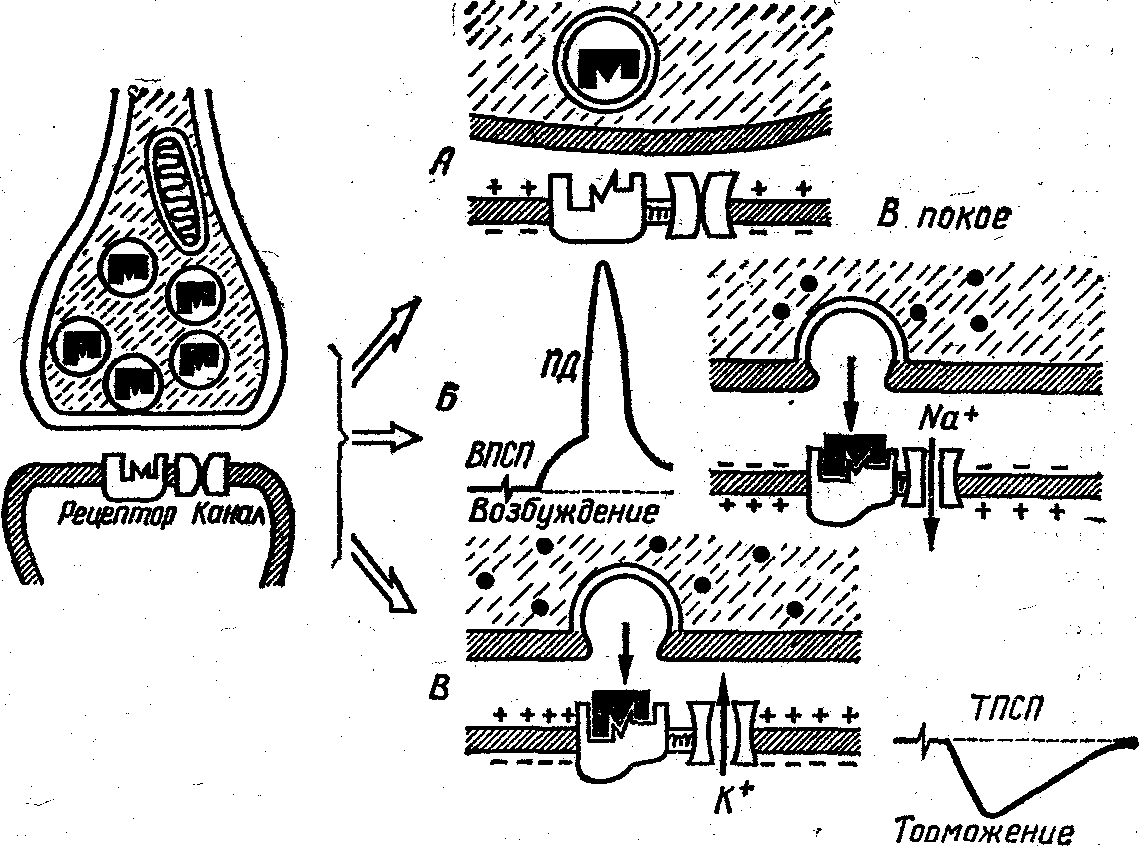
Рис. 1. Принцип действия возбуждающего и тормозного химического синапса.
Слева приведена схема химического синапса. Везикулы в нервном окончании содержат нейромедиатор. В постсинаптической мембране имеется рецептор нейромедиатора, сопряженный с ион-селективным каналом.
Справа: А — синапс в состоянии покоя; Б — нервный импульс вызывает поступление ионов Са в нервное окончание (показаны зачерненными кружочками), что запускает слияние везикулы с пресинаптической мембраной и высвобождение медиатора в синаптическую щель. В результате взаимодействия медиатора с рецептором открывается сопряженный с ним Na+—селективный канал и поступление Na+ в постсинаптический нейрон вызывает деполяризацию постсинаптической мембраны — возбуждающий постсинаптический потенциал (ВПСП). По достижению ВПСП критического уродня открываются потенциал-зависимые Ма+-каналы (на схеме не показаны) и возникает распространяющийся потенциал действия (ПД); В — аналогично 6, но рецептор сопряжен с К+-кана-лом, поэтому открывание ион-селективных каналов приводит к возникновению кратковременного повышения мембранного потенциала — тормозного постсинаптического потенциала (ТПСП)
синаптические потенциалы, ВПСП) или гиперполярнза-ционные (тормозные постсинаптические потенциалы, ТПСП). Из-за малой величины они носят название миниатюрных.
Теперь вернемся к реальной ситуации, учитывающей факт большого количества везикул в нервном окончании. Когда нервное окончание деполяризуется нервным импульсом, открываются ион-селективные Са-каиа-лы и Са проникает в нервное окончание, вероятность слияния везикул с пресинапти ческой мембраной резко возрастает. Следствие—почти синхронное высвобождение нескольких сотен квантов медиатора. Суммарный постсинаптический потенциал в этом случае оказывается достаточным для активации потенциал-зависимых каналов. Возникает распространяющийся потенциал действия.
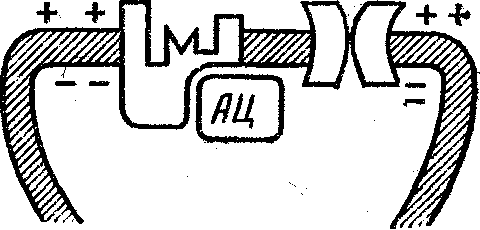
Таким образом, мы проследили цепочку событий от потенциала в нервном окончании до потенциала действия в постсинаптическом нейроне, то есть весь процесс химической передачи нервного импульса. Мы описали синапс, в котором рецептор объединен с ионным каналом. В другом случае (рис. 2) рецептор постсинаптиче-ской мембраны сопряжен с ферментом аденилатцикла-зой. Этот фермент превращает аденозинтрифосфат (универсальный источник химической энергии в клетке) в циклический аденозинмонофосфат (цАМФ), рассматриваемый ныне в качестве «второго сигнала», преобразующего влияние нейромедиаторов, гормонов и других факторов в специфические для данного типа клеток изменения метаболизма. Посредством ферментов (про-теинкиназ) цАМФ фосфорилирует функционально важные белки, например белки ионных каналов. В зависимости от эффекта фосфорилировання (открывание, закрывание) и ионной избирательности канала произойдет деполяризация или гиперполяризация постсннап-тической мембраны, как и при воздействии на каналы, сопряженные с рецептором. Конечно, для возникновения постсинаптического сдвига потенциала в этом синапсе потребуется больше Времени.
Для беспрепятственного проведения нервного импульса через синапс медиатор должен находиться в связи с постсинаптической мембраной лишь ограниченное время, что достигается двумя способами. Такие нейро-медиаторы, как аминокислоты или катехоламины, ак-
10
тивно захватываются нервным окончанием и могут использоваться повторно. Ацетилхолин расщепляется ферментом ацетилхолинэстеразой, находящейся на постсинаптической мембране, на уксусную кислоту и холии. В нервное окончание в этом случае поступает холин.
Итак, мы вкратце ознакомились с принципом действия химического синапса. Что же это нам дало? В начале века Ч. Шеррингтон, анализируя деятельность нервной системы, впервые описал центральное возбуждение и центральное торможение в качестве основных синаптических процессов, которые определяют рефлек-
Покой
Пратеинкиназр, Возбуждение
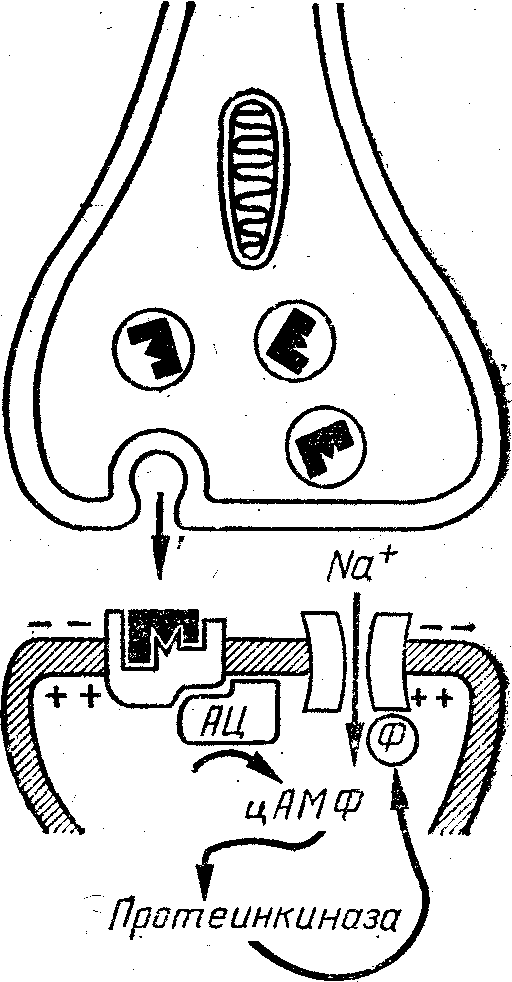
Р и с. 2. Принцип действия пептидергического «медленного» w напса.
Нейромедиатор-пептид взаимодействует ,с рецептором, сопряжет ным с аденилатциклазой, что приводит к увеличению содержаний цАМФ в цитоплазме постсинаптического нейрона и в результата активации протеинкиназ — к фосфорилиротванию ион-селективный каналов. В зависимости от избирательности каналов для ионоа' возникает либо возбуждение, либо торможение постсинаптическогД нейрона .- *
11
торную деятельность ЦИС. Из сказанного выше ясно, что указанные процессы отражают два варианта пост-сйнаптического ответа в химическом синапсе. Поначалу казалось, что должны быть какие-то принципиальные различия между веществами, являющимися тормозными и возбуждающими медиаторами. В самом деле, в центральной нервной системе теплокровных нейтральные аминокислоты (глицин, гамма-аминомасляная кислота (ГАМК) опосредуют тормозные влияния, а кислые (глутаминовая, аспарагиновая кислоты) — возбуждающие. Однако, как показали исследования нервной системы других животных, ГАМК успешно выполняет и роль возбуждающего медиатора. Нейромедиаторы сами по себе не несут информации о физиологическом ответе—они лишь запускают цепочку событий. Такие важные характеристики, как продолжительность торможения или возбуждения, определяются временем пребывания постсинаптических каналов в открытом состояния. Тем не менее, если представить поверхность нейрона в виде мозаики структурных элементов, реагирующих запрограммированным образом, то включение каждого исполнительного устройства осуществляется специфическим химическим сигналом, являющимся не только исполнителем, но и «визитной карточкой» пресинаптиче-ского нейрона, входящего в цепочку нейронов, которая осуществляет определенную форму нервной деятельности.
Особенно наглядно важность полноценной работы химических синапсов выявляется в условиях патологии, когда изучение синаптической передачи может дать ключ к пониманию механизма нарушения функций нервной системы.
Клиническая картина столбняка и ботулизма (отравление колбасным ядом) совершенно различна: неконтролируемые судороги при столбняке и мышечная слабость при ботулизме. Тем не менее механизм действия обоих бактериальных токсинов сходен: они повреждают механизм секреции медиаторов. Столбнячный токсин нарушает секрецию тормозных аминокислот в центральной нервной системе, что-приводит к беспрепятственному распространению возбуждения. Колбасный яд блокирует секрецию' ацетилхолина в нервно-мышечном соединении, таким образом, делает невозможным управление мышцами.
12
То, что именно нервные окончания являются ми< шенью действия столбнячного токсина, было показано в нашей лаборатории. Мы выделили из мозга нервные окончания, ив течение одного-двух часов воздействовали на них столбнячным токсином. Отравленные столбнячным токсином окончания высвобождали гораздо меньше тормозной аминокислоты в ответ на деполяризацию, чем в контроле. Таким образом удалось в пробирке воспроизвести основной синаптический дефект, лежащий в основе возникновения столбнячных судорог,
Другим ярким примером прееинаптической патологии является болезнь Паркинсона, при которой вследствие гибели части тормозных дофаминергических нейронов возникает мышечный тремор (дрожание), делающий невозможным осуществление точных произвольных движений. После выяснения природы нарушения синаптической функции стали использовать введение (больным предшественника дофамина — L-диоксифенилала-нина (L-ДОФА) с целью усиления синтеза дофамина в нервных окончаниях. Препарат успешно устранял симптомы болезни.
Теперь, когда мы познакомились с работой химического синапса, остановимся на распространенных определениях химических сигналов. Нейромедиатором (трансмиттером) называют вещество, передающее нервный импульс. Результатом его действия является возникновение электрического потенциала. Это обстоятельство потребовало возникновения специализированной анатомической структуры—синапса. Быстрое действие -достигается за счет малого расстояния между секрети-рующей клеткой и мишенью, а также молниеносными способами уборки медиатора с мишени. Рецепторы и сопряженные с ними каналы собраны на ограниченном участке под окончанием, что способствует возникновению максимального постсинаптического потенциала, причем в совершенно определенной точке клетки (клеточный контакт). Рецепторы химического сигнала находятся на поверхности мембраны, и это важно для опознания клеткой нейромедиаторной роли вещества. Так, аминокислота глицин внутри нейрона—заурядный участник множества биохимических реакций. Тот же глицин, приложенный извне, вызывает опосредуемый рецептором тормозной постсинаптический потенциал. Нейромодулятором называют вещество, изменяющее
13
эффективность синаптической передачи. Оно может секретироваться вместе с,медиатором (ко-медиатор) одной и той же нервной терминалью, соседним нервным окончанием или секреторной клеткой железы, далеко отстоящей от нервной системы. Ни быстрое действие, ни генерация электрического потенциала не являются обязательными характеристиками эффекта нейромоду-лятора. Рецепторы для нейромодуляторов могут находиться как на внешней стороне мембраны в любой точке поверхности, так и внутри клетки. Из сказанного ясно, что понятие «нейромодулятор» является более широким, чем иейромедиатор. В роли нейромодуляторов могут выступать и специализированные продукты желез внутренней секреции—гормоны. Так, у самок крыс половые гормоны могут влиять на рецепторы дофамина в мозге и, таким образом, изменяя эффективность до-фаминергической " передачи, вызывать специальные формы поведения животного. Подобные отношения мозга и тела очень важны для адаптации (приспособления) животного к условиям окружающей среды. Стоит указать на условность отождествления конкретного вещества с гормоном или нейромедиатором. Окситоцин и вазо-прессин синтезируются в нейронах гипоталамуса и по аксонам перемещаются в нервные окончания, находящиеся в задней доле гипофиза. Здесь они высвобождаются в кровь, достигают тканей — мишеней, в результате чего регулируется тонус мышц матки и реабсорб-ция воды. Другие нейроны гипоталамуса направляют свои аксоны к различным отделам мозга, где долговременно изменяют эффективность синаптической передачи (память!), о чем пойдет речь ниже.
В настоящее время существуют надежные способы определения химической специализации синапсов в мозге. Ниже приведены данные об удельном весе синапсов с указанием времени обнаружения соответствующего медиатора (по Д. Кригер). Указанные вещества считают классическими нейромедиаторами (табл. 1).
Приведенные в таблице медиаторы (за исключением ацетилхолина) являются аминокислотами или производными аминокислот. Благодаря тому что число соответствующих синапсов в мозге неодинаково, содержание этих веществ также различно. Из таблицы следует также, что на долю так называемых классических нейроме-
14
Таблица 1 Классические нейромедиаторы (по Д. Кригер)
Год обнаружения
Доля от общего числа синапсов, %
Наименование
25-40 0,5 0,5
остальные
1920 1940
5—10 0,5
Ацетилхолин
Норадреналин
Аминокислоты:
1950—1960 1960—1970
глутаминовая аспарагиновая у-аминомасляная Дофамин Серотонин
диаторов приходится менее 50% синапсов, имеют другую химическую специализацию.
ПЕПТИДЫ- НОВЫЙ КЛАСС ХИМИЧЕСКИХ СИГНАЛОВ В МОЗГЕ
КАК ИХ НАЗЫВАТЬ?
Распространенные названия пептидов отражают их физиологическое действие, особенности структуры, источник, из которого впервые был выделен пептид, и т. п. Так, после обнаружения опиоидных пептидов они Получили название эндогенных морфинов (эндорфинов), а позднее другой пептид с еще более сильным морфи-ноподобным действием так и был назван динорфин (дина = «сила» + окончание слова «морфин»). Холеци-стокинин в дословном переводе означает пептид, вызывающий движение желчного пузыря. Наконец, название «вещество Р» отражает этап получения активного препарата.
Ни локализация, ни функция пептида не могут быть основой рациональной классификации. В самом деле, исследования показали, что аналогичные или весьма сходные с пептидами млекопитающих химические соединения могут быть обнаружены у более простых организмов, то есть в эволюционном плане пептидные регуляторы возникают раньше, чем новая структура и новая функция, для выполнения которой природа приспосабливает уже существующий пептидный регулятор. Знание структуры очень важно, оно позволяет на логически ясной основе объединить пептиды животных раз-
13
вых видов в семейства и сделать заключение о возможном сходстве действия. Однако следует иметь в виду, что физиологический эффект определяется не только структурой пептида, но и особенностями рецептора, с которым он реагирует, каналом или ферментом, с которым связан рецептор. Следовательно, по необходимости приходится изучать и сам пептид, и его биологическое действие у каждого вида. Ниже приводится неполный перечень известных пептидов, в котором указаны локализация пептида, название и структурное сходство с другим (в табл. 2 соответствующий пептид помечен значком §§ и помещен ниже гомологичного).
Таблица 2 КЛАССИФИКАЦИЯ ПЕПТИДОВ
I. Пептиды мозга и желудочно-кишечного тракта:
Опиоидные пептиды (эндорфины)
Метионин-энкефалин Лейцин-энкефалин §§ а-эндорфин §§ динорфин (ы) §§ у-эндорфин §§ р-нео-эндорфин ' §§ Р-эндорфин §§ а-нео-эндорфин
И. Гипоталамические рилиэичг-факторы (РФ), действующие на переднюю долю гипофиза:
РФ лютеннизирующего гормона (ЛГ—РФ)