
На правах рукописи
| Вид материала | Автореферат диссертации |
- Печатная или на правах рукописи, 21.09kb.
- Удк 796/799: 378 , 770.24kb.
- На правах рукописи, 399.58kb.
- На правах рукописи, 726.26kb.
- На правах рукописи, 1025.8kb.
- На правах рукописи, 321.8kb.
- На правах рукописи, 552.92kb.
- На правах рукописи, 514.74kb.
- На правах рукописи, 670.06kb.
- На правах рукописи, 637.26kb.
Палеорастительность и палеогеография
раннесуббореального периода (4 500 – 3 200 лет назад)
Рубеж атлантического и суббореального периода ознаменовалась самым массовым облесением и максимальной выраженностью всех процессов, начавшихся в позднеатлантическое время. На всех средних диаграммах именно в это время выражен абсолютный минимум пыльцы трав и максимум деревьев, в которых также максимальное содержание пыльцы широколиственных пород (см. рис. 8, табл. 4). Можно сказать, что климатический оптимум голоцена на территории центральной части Приволжской возвышенности наиболее яркое выражение имел именно в самом конце атлантического – начале суббореального периода.
Р


ис. 8. Средние спорово-пыльцевые комплексы основных таксонов SB-1 – периода
Такое предположение автор работы высказывал и ранее, но оно получило подтверждение радиоуглеродными датировками только в настоящее время: 4590±120 и 4495±85 лет назад (см. табл. 1). Возможно, это связано с более восточным положением исследуемой территории. Первая половина суббореального периода на исследуемой территории характеризуется прохладным и влажным климатом. Несмотря на это, по данным А.Л. Никитина (1978), в первой половине периода произошло значительное понижение уровня грунтовых вод. В это время происходит окончательное формирование серых лесных почв. В Западном, Северо-Западном, Восточном районах снижение уровня грунтовых вод и похолодание климата приводят к значительному расширению участия в сосново-широколиственных и широколиственных лесах Betula pendula и заметному снижению роли Quercetum mixtum. Господство переходит к сосново-березово-широколиственным лесам, сосново-березовым лесам зеленомошникам, иногда почти чиcтым березовым, а на южных склонах возвышенностей – к сосновым и сосново-березовым остепненным лесам. В Северо-Западном районе в основном господствовали березовые леса дубравные с заметным участием Tilia cordata, подлеском из Corylus avellana, Sorbus aucuparia, Euonymus verrucosa с единичными деревьями Pinus sylvestris. В травяном ярусе были распространены Carex sp. Подобные леса занимали пологие склоны возвышенностей с супесчаными и суглинистыми почвами. Такие леса в настоящее время встречаются очень широко на Приволжской возвышенности, но некоторыми исследователями считаются вторичными. Несколько иначе формировалась лесная растительность в это время в лесном Северном и лесостепном Северо-Восточном районах. Здесь роль Betula pendula практически не изменилась. Как и повсюду, на рубеже атлантического и суббореального периодов произошел широкий расцвет сосново-широколиственных лесов (в Северном районе), дубняков сложных и липовых травяных лесов (в Северо-Восточном), но они без каких-либо изменений существовали и весь раннесуббореальный период. Снижения уровня грунтовых вод в данных районах не было. В Южном районе роль березовых древостоев возрастает вдвое, что говорит о развитии здесь наряду с широколиственными лесами (дубняками сложными, дубняками остепненными) березняков дубравных. Степные ценозы занимали те же площади с теми же сообществами что и в позднеатлантическое время, то есть преобладали лугово-разнотравные, лугово-типчаковые, ковыльно-разнотравные сообщества. Но в их составе уже появляются рудеральные и пасквальные сорняки.
Первая зона суббореального периода относится к эпохе энеолита и началу эпохи бронзы (Крайнов и др., 1984). В это время резко увеличивается общая заселенность данной территории, что связано с грандиозными миграциями, охватившими население степных зон. В результате катастрофических засух из-за резкого падения уровня грунтовых вод падает биологическая продуктивность степных районов. Многие скотоводческие племена двинулись в лесную зону, где использовали для выпаса скота поймы рек и опушки лесов. На рубеже атлантического и суббореального периодов в пыльцевых спектрах Восточного и Северо-Западного районов появляются типичные «индикаторы пастбищ неолита» - Rumex acetosa, Ranunculus repens, Chenopodium album (Van Zeist, Casparie, 1974; Behre, 1981). Кривая пыльцы сорных видов на диаграммах с этого времени становится непрерывной. Во всех районах появляется пыльца Cerealia, красноречиво свидетельствующая о повсеместно начавшемся земледелии. Хотя роль и сорных, и культурных видов еще крайне мала. Таким образом, полученные палинологические данные в основных чертах подтверждают мнение В.М. Слободина (1952), Н.Я. Мерперта (1958), и А.Х. Халикова (1969) о начале земледелия в первой половине II тысячелетия до н.э.
Поскольку данное время знаменует очень важное событие в истории растительности, а именно: закат широчайшего распространения широколиственных пород и развитие березовых лесов дубравных, получены радиоуглеродные датировки этого «события» практически для всей изучаемой территории (см. табл. 1): 3810±80; 3400±75; 3975±80; 3500±60; 3390±60 лет назад.
Палеорастительность и палеогеография
позднесуббореального периода (3 200 – 2 500 лет назад)
Происходит дальнейшее похолодание климата, а в самом конце периода и увеличение общей влажности (Хотинский, 1970, 1977; Долуханов, 1971; Кинд, 1974; Елина и др., 1984, 2000). В почвенном покрове - важные изменения: формируется гуматно-кальциевый горизонт черноземных почв. На всей территории повсеместно снижается роль широколиственных пород и Betula pendula и резко возрастает – Pinus sylvestris, которая к концу периода достигает своего максимума (cм. табл. 4). В районах: Западном, Восточном, Северном и Северо-Западном лесная растительность развивалась схожим образом. Серьезное похолодание климата приводит, во-первых, к сокращению площади лесов и расширению – открытых участков (см. рис. 9). Во-вторых, в составе лесов резко возрастает роль Pinus sylvestris за счет постепенного сокращения участия Betula pendula и Quercetum mixtum. Уже сложно говорить о существовании чистых березовых, а тем более, широколиственных лесов. В тех же местообитаниях господство переходит сосново-березовым, сосново-широколиственным и сосновым лесам. В сообществах открытых пространств Восточного района увеличивается роль дерновинно-злаковых ценозов и сокращается – разнотравных. Повышается роль сорных видов, на некоторых диаграммах – полыней и (или) маревых. Такая же закономерность прослеживается в Северо-Восточном и Южном районах. Однако здесь общее облесение территории сокращается еще резче, а в составе лесной растительности участие широколиственных пород сокращается практически вдвое, но с учетом поправочных коэффициентов, широколиственные леса еще играли здесь заметную роль в тех же местообитаниях, что и в начале суббореального периода. Роль Pinus sylvestris в них также возрастает.
Р

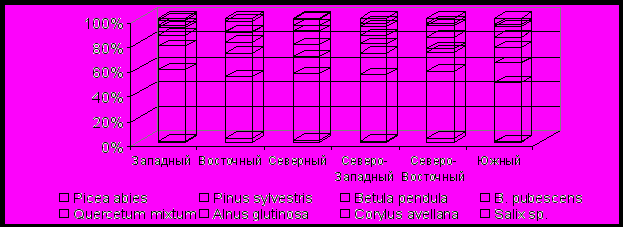
ис. 9. Средние спорово-пыльцевые комплексы основных таксонов SB-2 – периода
Характерно, что Alnus incana, игравшая небольшую, но заметную роль в сложении широколиственных и сосново-широколиственных лесов SB-1- периода, с этого времени практически выпадает из их состава. В составе степных ценозов также произошли изменения, связанные с одной стороны, с похолоданием климата: сокращение роли разнотравья и увеличение злаковых ценозов, с другой – с начавшейся заметной хозяйственной деятельностью человека (а именно, распашкой земель): вдвое увеличивается участие сорных и культурных видов, особенно в Северо-Восточном районе. С этого времени начинается рациональная кривая пыльцы Cerealia и развитие лесной и степной растительности уже нельзя рассматривать без учета хозяйственной деятельности человека.
С наступлением эпохи бронзы антропогенное воздействие на растительный покров резко возрастает. По мнение Н.Я. Мерперта (1958) именно к эпохе бронзы относится массовое заселение Среднего Поволжья. Древних племен привлекали богатейшие пойменные луга, благоприятные условия для подсечно-огневого земледелия и возможность широких связей по крупнейшим рекам (Волге, Суре, Алатырю). На изучаемой территории выявлены многочисленные стоянки поселений бронзового века. В основном господствовали племена срубной культуры (Буров, 1972, 1974). Основным их занятием было земледелие и скотоводство. В археологических раскопках курганов бронзового века обнаружено большое количество костей домашних животных: коров, овец, свиней, лошадей, а также примитивные мотыги и бронзовые топоры. Выпас скота в облесенных районах приводил к остепнению и изреживанию сосново-широколиственных лесов, уничтожению естественного травяного покрова и появлению пастбищных сорных видов. Вполне допустимо, что в это время человек специально выжигал леса, чтобы вызвать на гарях более развитый травостой. Хотя в литературе имеются данные, что человек мог вырубать леса каменными и бронзовыми топорами (Iversen, 1949, 1956), начавшееся земледелие не приводило к заметному сокращению площади лесов. Для населения этого времени вполне хватало имеющихся открытых пространств для возделывания богатых черноземных и дерново-карбонатных почв. Радиоуглеродные датировки этого времени: 2950±45 и 2730±50 лет назад.
Палеорастительность и палеогеография
раннесубатлантического периода (2 500 – 700 лет назад)
К
 онец суббореального периода (и всего среднего голоцена) и начало субатлантического (позднего голоцена) на территории центральной части Приволжской возвышенности всеми исследователями голоцена европейской части России единодушно характеризуется переломным этапом в развитии климатических процессов в сторону дальнейшего похолодания и увеличения влажности с одной стороны, и поднятия уровня грунтовых вод – с другой (Кинд; 1976; Хотинский, 1977; Серебрянная, 1982; Артюшенко, 1982; Шаландина, 1993; Rukla, Mattheus, 1972 и др.). Окончательно формируются все современные типы почв. В растительном покрове на всей изучаемой территории около 2 500 лет назад происходит возрастание в лесах роли Pinus sylvestris (на большей ее площади до максимальных за весь голоцен значений), падение роли до минимума за последние 8 000 лет Betula pendula и сокращение площади широколиственных лесов (см. табл. 4). Радиоуглеродная датировка этого времени - 2485±35 лет назад.
онец суббореального периода (и всего среднего голоцена) и начало субатлантического (позднего голоцена) на территории центральной части Приволжской возвышенности всеми исследователями голоцена европейской части России единодушно характеризуется переломным этапом в развитии климатических процессов в сторону дальнейшего похолодания и увеличения влажности с одной стороны, и поднятия уровня грунтовых вод – с другой (Кинд; 1976; Хотинский, 1977; Серебрянная, 1982; Артюшенко, 1982; Шаландина, 1993; Rukla, Mattheus, 1972 и др.). Окончательно формируются все современные типы почв. В растительном покрове на всей изучаемой территории около 2 500 лет назад происходит возрастание в лесах роли Pinus sylvestris (на большей ее площади до максимальных за весь голоцен значений), падение роли до минимума за последние 8 000 лет Betula pendula и сокращение площади широколиственных лесов (см. табл. 4). Радиоуглеродная датировка этого времени - 2485±35 лет назад. Р

ис. 10. Средние спорово-пыльцевые комплексы основных таксонов SA-1- периода
Начало субатлантического периода примерно совпадает с началом железного века на изучаемой территории. Площадь лесов практически повсеместно значительно сокращается, а открытых пространств – возрастает (см. рис. 10). Северо-Западный район с этого времени приобретает лесостепной облик. В Восточном районе пыльца трав начинает доминировать над пыльцой деревьев. Судя по значительному участию пыльцы культурных злаков и сегетальных сорняков, указанные изменения происходят за счет распашки и освоения земель. На песчаных почвах возвышенных участков Западного, и Северо-Западного районов сократилось участие сосняков зеленомошников, и они продолжали существовать в основном по окраинам водораздельных болот. Более значительную роль начинают играть сосняки остепненные и сосняки травяные. В травяном ярусе здесь преобладают Poaceae, Artemisia, Chenopodiaceae, многочисленные представители сем. Asteraceae, Brassicaeae, Polygonaceae. Определенную роль продолжали играть березово-сосновые леса остепненные. На склонах возвышенностей, у их оснований и на выровненных пониженных участках плакоров Западного, Северо-Западного и Восточного районов продолжали развиваться сосново-широколиственные леса (сосново-дубовые с хорошо развитым подлеском из Corylus avellana), но участие пород Quercetum mixtum, тем не менее, становится менее заметным. При появлении металлических орудий труда человек уже мог не только возделывать плодородные почвы степей, но и вести вырубки лесов, расположенных на глинистых и суглинистых почвах тяжелого механического состава. Лес в это время не только выжигался, но и вырубался после подсочки. После таких вырубок он частично восстанавливался, так как древние земледельцы сеяли на одном месте только три-четыре года подряд, а затем переходили на другие поля. Однако происходили изменения в составе самих лесов. Свободные пространства в первую очередь занимала Betula pendula, как порода пионерная. Резкое сокращение лесных площадей произошло в лесостепных и степных районах. Благодаря формированию и широкому распространению здесь богатых черноземных почв, антропогенное воздействие человека было более значительным. Распашка земель приводила к значительному расширению открытых пространств. Поскольку почвы здесь очень плодородные, человеку не было необходимости менять часто места посевов. В этих условиях лес уже не восстанавливался. Вместе с Cerealia увеличивают свое содержание и сопутствующие сорняки: Centaurea cyanus, Fagopyrum tataricum, Consolida regalis, Polygonum aviculare. Растительный покров Северо-Восточного и Южного районов приобретает степной облик. Помимо агроценозов, повышается роль вторичных луговых степей с обилием сорных видов (Urtica dioica, Rumex acetosa, Cichorium intybus, Artemisia absinthium,Ranunculus repens, Taraxacum officinale и др.).
К концу SA-1- периода роль Quercetum mixtum значительно снижается и возрастает – Pinus sylvestris. Именно в это время (в конце I тысячелетия до н.э.) роль скотоводства в хозяйственной деятельности человека существенно возрастает, так как на территории Приволжской возвышенности стали распространяться кочевые азиатские племена – скифы и сарматы, а позднее и хазары, основным занятием которых было скотоводство. Интенсивный выпас скота в Восточном, Северо-Восточном и Южном районах, где в основном селились кочевники, а затем и булгары, привел к тому, что дерновинно-злаковые степи на многих участках стали превращаться в типчаковые, полынно-злаковые, полынные с большим участием сорных видов. В конце SA-1- периода (по данным археологов XIII – VII века н.э.) на территории восточной части Приволжской возвышенности сложилось государство волжских булгар, имеющих многочисленные населенные пункты. Их поселения тянулись на 500-600 м вдоль рек. Основным занятием булгар было хорошо развитое земледелие: хлеб даже вывозился в соседние государства. Такое расширенное земледелие, естественно, приводило не только к распашке свободных степных участков, но и требовало сведения лесов. Кривая пыльцы деревьев к концу SA-1 - периода на многих диаграммах разрезов всех районов резко падает. В основном уничтожались сосново-широколиственные, сосново-березово-широколиственные и широколиственные леса, расположенные на богатых почвах. В лесных ценозах возрастает роль вторичных порослевых дубняков, липняков, березняков, а также, видимо, осинников. Сокращается роль Pinus sylvestris, так как после вырубок в таких лесах ее возобновление крайне затруднительно. На месте сосняков зеленомошников на легких супесчаных и песчаных почвах после вырубок появляются сосняки травяные и сосняки остепненные, на что указывает и повышение содержания пыльцы разнотравья в типично лесных спектрах. В некоторых случаях, возможно, возникали песчаные степи. Большинство современных участков песчаных степей Западного, Северного и Восточного района, на наш взгляд, имеют раннесубатлантический возраст. Радиоуглеродные датировки этого времени: 1660±75; 1905±35; 810±45 лет назад.
Палеорастительность и палеогеография
среднесубатлантического периода (700 – 300 лет назад)
Данное сравнительное короткое время в субатлантическом периоде стало возможным выделить на основании анализа многих диаграмм центральной части Приволжской возвышенности. Формирование растительности этого времени связано как с климатическими причинами, так и историей хозяйственного освоения данной территории. 700 лет назад произошло резкое похолодание климата: так называемый «малый ледниковый период» (Кинд, 1976; Монин и др., 1979). Кроме того, начиная с XIV до конца XVII века нашей эры, на изучаемой территории произошло значительное сокращение антропогенного воздействия. Территория подверглась нападениям монгольских завоевателей и целиком вошла в государство Золотой Орды. После опустошительных походов Тамерлана на 400 лет она стала практически безлюдной (Чекалин, 1892). Антропогенное воздействие на растительность повсеместно становится минимальным, а в некоторых районах прекращается совсем. Климатические условия привели к тому, что во всех лесных районах и частично в Восточном лесостепном площадь лесов несколько расширяется (см. рис. 11).
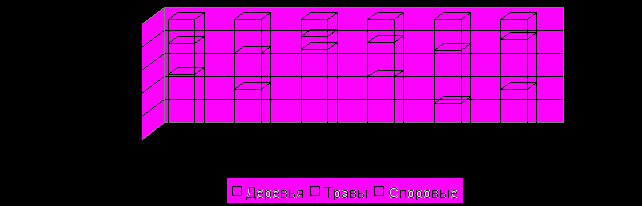

Рис. 11. Средние спорово-пыльцевые комплексы основных таксонов SА-2 – периода
Этому способствовали и супесчаные почвы, на которых, после забрасывания пашен не мог развиваться сомкнутый травостой с плотной дерниной. В составе лесов еще более повышается часто до максимальных значений роль Pinus sylvestris и снижается до минимума – широколиственных пород. Изменившиеся антропогенные условия явились причиной практически повсеместного сокращения в составе трав участия Cerealia и сорных видов. Начинают вновь господствовать сосновые леса (зеленомошники, долгомошники, папоротниковые) особенно в лесных районах. Видимо именно их многие исследователи приняли за единственную коренную формацию лесов, поскольку с этого времени уже имеются первые литературные сведения о составе растительности изучаемой территории. Меньшие площади занимали березово-сосновые, еще меньше - сосново-березово-широколиственные леса. На склонах водоразделов, а также на выровненных плакорах нижнего плато продолжали развиваться степные ценозы, как первичные, так и вторичные, возникшие на месте сведенных лесов человеком в начале периода. В их составе повышается роль Poaceae и разнотравья, уменьшается – сорных видов. Заброшенные после распашки земли зарастали злаковыми и разнотравно-злаковыми сообществами, а на суглинистых и глинистых почвах, скорее всего – ковыльно-разнотравными. Впоследствии в исторических актах XVIII века они были названы «дикими степями», «новями» и «порозжими землями». Их выделяли для освоения поселившимся здесь русичам, мордве и чувашам (Перетяткович. 1882). Именно их наблюдал в 1768 году П.С. Паллас, путешествуя вверх по Волге (Паллас, 1803). Некоторыми исследователями они были приняты за первичную и основную растительную формацию восточных и южных районов данной территории (Спрыгин, 1896, 1931). Несколько особым образом формировался растительный покров Северо-Восточного района. Здесь соотношение лесных и степных сообществ практически не изменилось. Видимо небольшие поселения здесь продолжали оставаться, и антропогенная нагрузка полностью не прекращалась, хотя и уменьшилась. Радиоуглеродная датировка этого времени - 405±55 лет назад (см. табл. 1).
Палеогеография позднесубатлантического периода
(300 – 0 лет назад)
С


приходом земледельческих племен 300 лет назад на изучаемой территории вновь увеличивается воздействие человека на природу. Повсеместно сокращается площадь лесов и расширяется – открытых пространств (см. рис. 12).
Рис. 12. Средние спорово-пыльцевые комплексы основных таксонов SА-3 – периода
Менее это заметно на диаграммах Западного и Восточного районов, на некоторых из них даже происходит некоторое увеличение пыльцы деревьев. Однако это связано отнюдь не с естественным восстановлением лесных растительных сообществ, а с лесовосстановительными работами, проводимыми в последнее столетие. В составе лесов Западного, Северо-Западного, Северного районов увеличивается роль вторичных березняков за счет вырубок сосны. Большую роль начинают играть сосняки травяные и остепненные
В районах с более богатыми почвами возрастает доля вторичных дубняков, липняков, осинников, что хорошо видно на диаграммах Северо-Восточного, Восточного, Южного районов. Содержание пыльцы трав существенно увеличивается за счет сорных и культурных видов. Естественные степные ценозы сохраняются только на неудобьях – высоких возвышенностях и их склонах. Изменения растительного покрова в результате антропогенных воздействий в течение последних десятилетий подробно рассмотрены в работах многих авторов, поэтому нет необходимости останавливаться подробно на этом вопросе.
7.2. Основные сукцессии палеорастительности в голоцене
Анализ средних спорово-пыльцевых комплексов всей территории показал, что растительный покров территории менялся от степного в древнем голоцене к лесостепному в раннем, лесному – в среднем и вновь к лесостепному - в позднем (см. рис. 13, табл. 4).
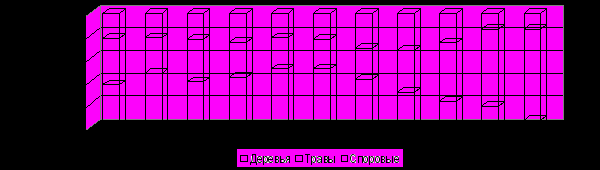
Рис. 13. Спорово-пыльцевые комплексы общего состава по периодам голоцена
На основе анализа вышеуказанного фактического материала, а также динамики пыльцы древесных пород и травянистых растений в голоцене (см. рис. 14, 15) основные этапы сукцессии лесных сообществ можно представить следующим образом. AL/DR- 3, DR-3: разреженные кустарниковые ивово-березовые (из Betula humilis) редкостойные сообщества → PB: разреженные сосновые леса зеленомошные, лишайниковые и остепненные, горные сосняки → ВО-1: настоящие сосняки зеленомошники, долгомошники, остепненные, папоротниковые; березово-сосновые зеленомошники, сероватовейниковые; на юге березовые остепненные и крупнотравные (аналог березовой лесостепи Урала) → ВО-2: повсеместный расцвет сосновых (зеленомошников, долгомошников, лишайниковых) и березово-сосновых (в зависимости от местообитания остепненных, сероватовейниковых) лесов → АТ-1: широкое распространение сосново-березовых и березово-сосновых (зеленомошных, крупнотравных, папоротниковых, остепненных) и березовых (крупнотравных) лесов. Появление в лесах широколиственных пород → АТ-2, АТ/SB: расцвет сосново-широколиственных (сосново-дубовых, сосняков сложных), широколиственных (дубняков сложных, на юге - остепненных); на севере сосново-елово-широколиственных лесов → SB-1: сосново-березово-широколиственные, сосново-березовые и березовые (зеленомошные); березовые (дубравные, заболоченные); реже сосново-широколиственные и широколиственные → SB-2: сокращение лесистости в результате сильного похолодания. Господство сосновых; изреживание и закат сосново-широколиственных лесов → SA-1: сокращение площади лесов (антропогенный фактор). Господство вторичных: сосновых (травяных, остепненных), реже березово-сосновых лесов. Появление вторичных березняков, дубняков, липняков, осинников → SA-2: расширение площади лесов за счет снижения антропогенной нагрузки: сосновых (зеленомошников, папоротниковых, долгомошников), реже сосново-березовых → SA-3: резкое сокращение лесистости (антропогенный фактор); господство вторичных сосновых (травяных, остепненных), березовых (дубравных, остепненных, крупнотравных), широколиственных, мелколиственных и смешанных лесов.
Таблица 4
| Средние спорово-пыльцевые комплексы голоцена всей территории центральной части Приволжской возвышенности, % | |||||||||||
| | | | | | | | | | | | |
| Растения | Периоды голоцена | ||||||||||
| SA-3 | SA-2 | SA-1 | SB-2 | SB-1 | AT-2 | AT-1 | BO-2 | BO-1 | РВ | DR3 | |
| Деревья: | 37,5 | 47,0 | 40,0 | 44,0 | 51,5 | 51,5 | 42,0 | 30,5 | 23,0 | 18,5 | 6,0 |
| Picea abies | 3,5 | 2,5 | 2,5 | 1,5 | 2,0 | 2,0 | 1,0 | 0,1 | 0,0 | 0,0 | 0,0 |
| Pinus sylvestris | 51,5 | 60,5 | 55,5 | 52,0 | 37,5 | 49,0 | 44,5 | 49,5 | 39,5 | 34,0 | 28,0 |
| Betula pendula | 22,0 | 16,0 | 16,5 | 16,0 | 23,0 | 15,0 | 31,5 | 19,0 | 13,5 | 4,5 | 4,0 |
| B.humilis | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,5 | 2,0 | 3,5 | 7,5 | 13,0 |
| B. pubescens | 10,0 | 5,5 | 7,5 | 8,0 | 9,5 | 9,0 | 12,0 | 13,0 | 9,0 | 4,5 | 3,0 |
| Quercus robur | 4,0 | 3,0 | 4,0 | 3,5 | 7,0 | 6,0 | 2,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| Tilia cordata | 2,5 | 2,0 | 2,5 | 2,5 | 3,5 | 4,0 | 1,0 | 0,1 | 0,0 | 0,0 | 0,0 |
| Ulmus spp. | 1,0 | 0,5 | 1,0 | 2,0 | 3,5 | 2,5 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 |
| Quercetum mixtum | 7,5 | 5,5 | 7,5 | 8,0 | 14,0 | 12,5 | 3,5 | 0,1 | 0,0 | 0,0 | 0,0 |
| Alnus glutinosa | 3,5 | 5,5 | 4,5 | 5,5 | 7,5 | 7,5 | 4,0 | 0,5 | 0,0 | 0,0 | 0,0 |
| A. incana | 0,0 | 0,0 | 0,2 | 0,1 | 0,5 | 0,1 | 0,2 | 0,0 | 0,0 | 0,0 | 0,0 |
| Corylus avellana | 2,5 | 2,0 | 2,0 | 4,0 | 4,0 | 4,0 | 1,5 | 0,0 | 0,0 | 0,0 | 0,0 |
| Acer platanoides | 0,5 | 0,1 | 0,2 | 0,5 | 0,1 | 0,2 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 |
| Salix spp. | 2,5 | 3,5 | 4,5 | 3,5 | 2,0 | 2,0 | 2,5 | 15,5 | 35,5 | 48,0 | 53,0 |
| Травы: | 40,5 | 31,5 | 37,0 | 30,5 | 27,0 | 25,5 | 27,5 | 37,0 | 51,0 | 67,5 | 80,0 |
| Poaceae | 27,5 | 26,5 | 26,5 | 28,0 | 25,0 | 26,0 | 24,0 | 17,5 | 18,0 | 11,0 | 10,0 |
| Cerealia | 9,0 | 2,0 | 4,5 | 1,0 | 0,5 | 0,1 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| Cyperaceae | 20,0 | 20,0 | 23,5 | 23,0 | 28,5 | 28,0 | 24,5 | 25,5 | 19,0 | 9,5 | 3,0 |
| Artemisia | 12,0 | 10,0 | 12,5 | 14,0 | 13,5 | 11,5 | 11,5 | 19,0 | 23,0 | 42,0 | 47,0 |
| Chenopodiaceae | 7,5 | 4,5 | 5,0 | 7,0 | 6,5 | 6,0 | 8,0 | 19,0 | 27,0 | 35,5 | 39,0 |
| сорные травы | 10,0 | 8,0 | 9,0 | 1,7 | 2,0 | 0,5 | 0,1 | 0,0 | 0,0 | 0,0 | 0,0 |
| Ericaceae | 1,0 | 1,5 | 0,7 | 1,0 | 1,0 | 1,0 | 0,1 | 0,0 | 0,0 | 0,0 | 0,0 |
| Varia | 16,0 | 18,5 | 14,0 | 14,5 | 16,5 | 17,5 | 21,5 | 12,0 | 8,0 | 2,0 | 0,0 |
| водные травы | 2,5 | 7,0 | 5,5 | 7,0 | 4,0 | 5,5 | 7,0 | 5,0 | 3,5 | 0,5 | 0,5 |
| Споровые: | 22,0 | 21,5 | 23,0 | 25,5 | 21,5 | 23,0 | 30,5 | 32,5 | 26,0 | 14,0 | 14,0 |
| Sphagnum | 30,0 | 17,0 | 23,5 | 27,0 | 23,5 | 22,0 | 10,0 | 0,5 | 0,0 | 0,0 | 0,0 |
| Bryales | 57,5 | 70,5 | 65,0 | 61,0 | 62,0 | 59,0 | 60,0 | 43,5 | 38,0 | 25,0 | 21,0 |
| Polypodiopsida | 10,5 | 8,5 | 9,0 | 11,0 | 14,0 | 18,0 | 27,5 | 43,5 | 38,0 | 34,0 | 30,0 |
| Equisetum sp. | 2,5 | 3,5 | 1,5 | 1,0 | 0,5 | 1,0 | 2,0 | 12,0 | 24,0 | 41,0 | 49,0 |
| Lycopodium sp. | 0,0 | 0,5 | 0,6 | 0,3 | 0,3 | 0,2 | 0,5 | 0,5 | 0,0 | 0,0 | 0,0 |
В степных сообществах ход сукцессий проявлялся следующим образом.
AL/DR-3, DR-3: перигляциальные холодные степи из разреженных полынно-маревых, полынных, кустарничково-зеленомошных сообществ → PB: сомкнутые полынно-марево-злаковые ценозы → ВО-1: настоящие полынно-марево-злаковые; злаково-разнотравные; каменистые степи → ВО-2: луговые злаково-разнотравные и дерновинно-злаковые степи → АТ-1, АТ-2: луговые (разнотравно-злаковые из мезофильных трав, тырсовые, перистоковыльные); и песчаные степи. Появление рудеральных сорных видов → SB-1: луговые степи с заметным участием пасквальных сорных видов, каменистые и песчаные степи → SB-2: сокращение разнотравных и расширение дерновинно-злаковых ценозов с участием полыней, маревых и сорных видов (пасквальных и сегетальных). Появление первых агроценозов → SA-1: вторичные луговые степи с обилием сорных видов (рудеральных, пасквальных и сегетальных), вторичные типчаковые на месте дерновинно-злаковых, вторичные полынно-злаковые и полынные степи, вторичные песчаные степи на месте сведения сосновых лесов. Расширение агроценозов → SA-2: вторичные злаково-разнотравные, ковыльно-разнотравные степи. Сокращение агроценозов → SA-3: современные вторичные луговые, каменистые, песчаные степи и агроценозы.
7.3. Лесообразующая роль древесных пород в голоцене
Анализ средних спорово-пыльцевых диаграмм и выявленные основные сукцессий палеорастительности показали, что лесообразующая роль различных древесных пород в отдельные периоды голоцена была далеко неодинаковой и определялась комплексом причин: климатическими, эдафическими, экологическими и антропогенными. В работе подробно рассмотрено участие каждой породы в сложении древесных сообществ, лесообразующая роль и причины ее изменения в различные периоды голоцена, время появления на изучаемой территории, расцвета и угасания.
Наибольшее распространение и роль в лесах играла Pinus sylvestris. Тем не менее, были периоды, когда она уступала другим породам или значительно сокращала свое участие в лесных фитоценозах: в АТ-1- периоде из-за экспансии березы; в SB-1 - из-за массового распространения широколиственных пород в эпоху климатического оптимума голоцена; в современное время (SA-3) - в результате хозяйственной деятельности человека. Самое значительное участие Betula pendula было в составе лесов АТ-1- и SB-1- периодов. В это время она не только играла равную роль в сосново-березовых лесах, но и повсеместно образовывала чистые березовые леса. Betula humilis в древнем и раннем голоцене входила в состав растительных сообществ не только пойм, но и перигляциальных холодных степей повсеместно, и ее участие было максимальным. Перестала играть какую-либо роль в сложении растительных сообществ около 6 000 лет назад и в современном растительном покрове практически не встречается. Широколиственные породы (в основном Quercus robur, Tilia cordata, менее Ulmus laevis, U. glabra, Acer platanoides, A. tataricum, Corylus avellana, Fraxinus excelsior, Alnus incana) появились лишь в самом конце ВО- периода (около 8 000 лет назад), начало их массового распространения относится к середине АТ- периода (около 6 000 лет назад), наибольшего развития – в климатический оптимум голоцена (5 000 – 4 000 лет назад), заката – в SB-2- период (начиная с 3 200 лет назад). Таким образом, не подтверждается мнение о рефугиуме широколиственных пород в последнее оледенение и значительном участии в лесах, начиная с древнего голоцена. Picea abies появилась в лесах в ВО-2- периоде, но на протяжении всего голоцена существенной роли в лесах никогда не играла. Мнение исследователей, что в доагрикультурное время, начиная с древнего голоцена, она была повсеместно и широко распространена, не подтверждается. Salix spp. играла значительную роль в древостоях древнего и раннего голоцена. Начиная с АТ-1- периода, резко сокращает свое участие.