
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
СодержаниеСтресс у растений Количество пыльцы как передатчик экологической информации у растений Lycopersicon esculentum x Solanium pennellii |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Стресс у растений
У гермафродитных растений существует заметная тенденция увеличивать соотношение количества пыльцы к продукции семян в условиях стресса, таких как засуха (Galen, 2000), недостаток питательных веществ (Helsop-Harrison, 1957), повреждение растительноядными животными (Cobb e.a., 2002) или патогенная инфекция (Lokesha, Vasudeva, 1993).
Количество пыльцы как передатчик экологической информации у растений
Отличительной чертой растительных популяций является неподвижный образ жизни и привязанность к месту обитания. Каковы же источники экологической информации у растений? Как они “узнают” об экологической ситуации?
Конечно, если стало холоднее или жарче, влажнее или суше, информацию об этом растение получает непосредственно от изменившихся факторов среды (охлаждаясь или высыхая). Но если нарушилось оптимальное соотношение полов, скажем возник дефицит мужского или женского пола, как передается такая информация в популяции растений? Поиски механизмов передачи экологической информации у растений привели к открытию новых закономерностей, лежащих в основе взаимоотношений популяции растений со средой (Геодакян, 1977а).
Рассматривая ареал растительного вида можно считать, что в центре ареала (внутри, в глубине) существуют в среднем более оптимальные условия, в то время как на окраинах, на его периферии—более экстремальные. Само существование границы ареала означает, что в своей естественной экспансии вид наткнулся в этом месте на “стенку” экологической ниши по тому или иному фактору среды. На разных участках границы такими факторами могут быть низкие или высокие температуры или различные концентрации: влаги, питательных веществ, врагов и паразитов. Очевидно также, что в центре ареала плотность популяции максимальна, а на периферии—минимальна.
Естественно предположить, что в реализации информационной связи между растениями главную роль может играть количество пыльцы, попадающее на женский цветок. Например, у двудомных или перекрестноопыляющихся видов, при прочих равных обстоятельствах, попадание большого количества пыльцы на пестик женского растения означает, что вокруг много мужских растений. Наоборот, малое количество пыльцы—сигнал о том, что поблизости мало мужских растений. Очевидно также, что в центре ареала количество пыльцы, попадающее на рыльце женского цветка будет в среднем всегда больше, чем на периферии ареала, из-за разной плотности популяции. (Речь идет о пыльце своего вида, количество “чужой” пыльцы, очевидно будет больше, наоборот, на периферии ареала.) Далее, во всех экстремальных условиях среды в первую очередь гибнут мужские особи, что также приводит к уменьшению количества пыльцы.
Следовательно, количество пыльцы может дать информацию женскому цветку о плотности популяции, о третичном соотношении полов вокруг него, о его местонахождении в центре или на периферии ареала, и об оптимальных или экстремальных условиях среды. Получение большого количества пыльцы всегда несет информацию о благоприятных условиях среды и требует производства больше женских потомков с малой фенотипической дисперсией. Получение малого количества пыльцы, наоборот, несет информацию о неблагоприятных условиях. Это бывает либо на периферии, где сильно падает плотность популяции, либо в центре, но при наступлении там экстремальных условий, которые, как мы видели, элиминируют в первую очередь мужских особей. И то, и другое требует производства больше мужских потомков с большой фенотипической дисперсией и максимальным половым диморфизмом, чтобы форсировать поиск новых путей развития.
Таким образом, во всех оптимальных условиях среды количество пыльцы, попадающее на женский цветок, бывает в среднем всегда больше того количества, которое попадает в экстремальных условиях. Иными словами, малое количество пыльцы всегда означает “плохо”, а большое количество всегда означает “хорошо”.
По существующим в классической генетике представлениям, пыльца несет только генетическую информацию и количество пыльцы, попадающее на женский цветок, не играет никакой роли, поскольку для оплодотворения достаточно одного пыльцевого зерна. По новым представлениям пыльца несет не только генетическую информацию, но и экологическую, регуляторную, которая кодируется количеством пыльцы, попадающим на женский цветок. Описанную схему связей иллюстрирует Табл. 14.1.
Такова теоретически ожидаемая картина. Теперь посмотрим, насколько она соответствует действительности. Несет ли пыльца помимо генетической информации еще и экологическую, регуляторную? Существует ли на самом деле зависимость эволюционной пластичности популяции, определяемой соотношением полов, дисперсией и половым диморфизмом потомства от количества пыльцы, попадающей на рыльце женского цветка?
Табл. 14.1 Связь основных характеристик раздельнополой популяции
с условиями среды и эволюционной пластичностью вида
(Геодакян, 1977).
| Центр | ← | Место в ареале | → | Периферия |
| | | ↓ | | |
| Оптимальные | ← | Условия среды | → | Экстремальные |
| | | ↓ | | |
| Max | ← | Плотность популяции | → | Min |
| | | ↓ | | |
| Max | ← | Пыльца своего вида | → | Min |
| | | ↓ | | |
| Min | ← | Пыльца других видов | → | Max |
| | | ↓ | | |
| Min | ← | Дисперсия Соотношение полов Половой диморфизм | → | Max |
| | | ↓ | | |
| Min | ← | Эволюционная пластичность | → | Max |
Влияние количества пыльцы на разнообразие фенотипов потомства было обнаружено Д. В. Тер-Аванесяном. У хлопчатника, вигны, подсолнечника и пшеницы при опылении малым количеством пыльцы, когда в процессе оплодотворения могут участвовать неконкурентоспособные гаметы, наблюдалось увеличение фенотипической дисперсии потомства (Табл. 14.2 Приложение В). Тер-Аванесян так прямо и пишет, что ограниченное опыление рылец цветка приводит к тому, что “в результате вместо выравненных сортов получаются популяции” (Тер-Аванесян, 1949; Ter-Avanesian, 1978).
Ограниченное опыление межвидового гибрида Lycopersicon esculentum x Solanium pennellii приводило к увеличению дисперсии по ряду количественных признаков проростков (число листьев, длина гипокотиля, масса проростков), а у гибрида lut1C x racemigerum—к повышению выхода рекомбинантных форм по маркерным генам (Жученко, Король, 1985, с. 318).
Эволюционная трактовка наблюдаемых явлений в этих и других работах отсутствует. Таким образом, зависимость соотношения полов и дисперсии потомства от количества пыльцы действительно существует. Что же касается полового диморфизма, то вряд ли кто-нибудь исследовал специально его зависимость от количества пыльцы. Это предсказание теории возможно проще проверить не в специальном эксперименте, а в наблюдениях в центре и на периферии ареала растительного вида. Например, как известно, у тополя листья женских деревьев имеют более продолговатую форму, а мужских—более округлую. Значит, на границах ареала тополя эти различия должны быть резче выражены, чем в глубине ареала.
Знание и учет этих зависимостей может повысить эффективность практических работ, в частности селекционных. Так, если стоит задача сохранения существующих форм, например в коллекциях генофонда растений, то при пересевах семян необходимо опылять большим количеством пыльцы и создавать растениям оптимальные условия (поливать, удобрять, защищать от холода), то есть необходимо искусственно создавать оптимальные условия центра ареала, с тем чтобы свести к минимуму изменчивость и сохранить существующий генотип. В этом случае, надо думать, так и делается.
В тех же случаях, когда стоит обратная задача—селекционая, когда пытаются получить новые, до того не существовавшие формы, необходимо создавать условия “периферии” природного ареала, то есть опылять малым количеством пыльцы и создавать растениям, по возможности, экстремальные условия (не поливать, не удобрять). По всей видимости мало кто из селекционеров поступает таким образом сознательно.
Наоборот, в стремлении получить крепкое, жизнестойкое потомство, селекционер, видимо, никогда не жалеет пыльцы при скрещиваниях и все время опыляет большим её количеством. Кроме того он старается, надо полагать, создать растениям оптимальные условия. Как мы видели, и то и другое способствует появлению минимального количества мужских особей, уменьшает фенотипическое разнообразие и половой диморфизм, то есть стабилизирует популяцию. Выходит, селекционер невольно ухудшает свой шанс найти новые формы. Он работает как бы в условиях центра ареала, в то время как ему надо работать в условиях “периферии”. Как это сделать?
Сначала надо исследовать зависимость разнообразия потомства от количества пыльцы. С увеличением числа пыльцевых зёрен, попадающих на женский цветок, фенотипическая дисперсия потомства сначала может возрастать до некоторого максимума, а при дальнейшем увеличении количества пыльцы должна падать до какого-то минимума. Максимум разнообразия соответствует оптимальному значению количества пыльцы для периферии ареала данного вида, а минимум соответствует условиям глубины ареала (Рис. 14.1).
Р
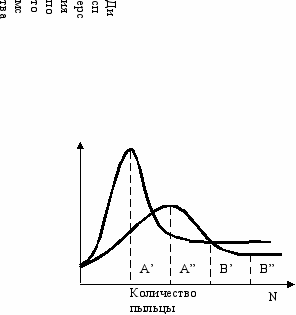 ис. 14.1
ис. 14.1Теоретические зависимости фенотипической
дисперсии потомства от количества пыльцы.
A’, A”—оптимумы для периферии ареала,
или для селекции
B’, B”—минимумы для глубины ареала, или
для сохранения генотипа.
Очевидно, что для разных видов, в зависимости от их биологии и экологии (энтомофилы, анемофилы и др.), значение этого максимума будет разным. Например, по данным Тер-Аванесяна, он имеет значения: для хлопчатника—около сотни пыльцевых зерен, для вигны— 40–50, для пшеницы— 10–20 (Тер-Аванесян, 1949). Определив этот максимум, дальше необходимо опылять женские растения именно этим, минимальным, но достаточным количеством пыльцы, и создавать растениям по возможности экстремальные условия периферии ареала. Это повысит эффективность селекции. Ведь известно, что естественное формообразование идет интенсивно именно на границах ареалов. Следовательно, при селекции, ставящей целью искусственное формообразование, создание новых сортов, необходимо по возможности повторять условия границ ареалов (экстремальные условия, мало пыльцы своего вида и много чужой). С этих позиций становятся понятными также описанные неоднократно в научной литературе эффекты “чужой” пыльцы (гетеропыльцы, пыльцы других видов), для понимания и объяснения которых существующих представлений классической генетики было недостаточно. Здесь мы имеем яркий пример того, как теория, будучи не в состоянии объяснить наблюдаемые факты, просто игнорировала или отвергала их.
Поскольку селекцией человечество занимается многие столетия, и она фактически была и продолжает в значительной мере оставаться не столько наукой, сколько искусством, естественно предположить, что некоторым выдающимся селекционерам удавалось интуитивно уловить те закономерности, о которых идет речь. В этой связи представляется очень интересным следующее высказывание И. В. Мичурина. Говоря об условиях успеха в получении новых сортов гибридизацией, он пишет “... ни в коем случае не следует давать сеянцам тучного состава почвы, а тем более надо избегать применения каких-либо удобрений ... Необходимость такого режима воспитания гибридов настолько резко выразилась в деле, что вынудила меня в 1900 г. продать бывший под питомником черноземный участок земли и подыскать для перемещения питомника другой участок с наиболее тощей песчаной почвой. Иначе я никогда не достиг бы успеха в выведении новых сортов плодовых растений и в введении в культуру у нас новых видов растений” (Мичурин, 1950).
Новое понимание дает возможность уже целенаправленно искать конкретные механизмы реализации, осуществляющие связи количества пыльцы с дисперсией, соотношением полов и половым диморфизмом потомства, и открыть эти, неизвестные ещё науке, но безусловно существующие и имеющие большое значение для практической селекции явления.