
«Российский государственный медицинский университет Федерального агентства по здравоохранению и социальному развитию»
| Вид материала | Автореферат |
СодержаниеВременнáя организация пролиферации обновляющихся тканей эпифизэктомированных беспородных белых крыс после совместного введения п Исследование роли эпифиза в формировании биоритмов |
- «Российский государственный медицинский университет Федерального агентства по здравоохранению, 975.58kb.
- «Российский государственный медицинский университет Федерального агентства по здравоохранению, 545.1kb.
- «Сибирский государственный медицинский университет Федерального агентства по здравоохранению, 927.55kb.
- Кубанский государственный медицинский университет федерального агентства по здравоохранению, 821.27kb.
- «Казанский государственный медицинский университет Федерального агентства по здравоохранению, 863.64kb.
- «Саратовский государственный медицинский университет имени В. И. Разумовского Федерального, 378.09kb.
- Гепатопротекторные свойства флавоноидов (фармакодинамика и перспективы клинического, 671.3kb.
- «ультразвуковая диагностика кишечной непроходимости» 14. 00. 27 хирургия, 401.81kb.
- «Рязанский государственный медицинский университет имени академика И. П. Павлова Федерального, 241.22kb.
- Программа подготовки аспирантов по кардиологии составили: д м. н., профессор, 643.17kb.
Параметры циркадианных ритмов МИ эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов эпифизэктомированных беспородных белых крыс после введения пептидов эпифиза
| Параметр | Пищевод | Кишечник | Лимфатический узел | |||
| 1 сутки | 2 сутки | 1 сутки | 2 сутки | 1 сутки | 2 сутки | |
| Мезор,‰ | 7±1.1 | 68.8±4.22 | 24±2.35 | |||
| Акрофаза,ч | 900 | 900 | 2400 | 2400 | 1200 | 1200 |
| АФ,ч | 700-1400 | 600-1400 | 2000-200 | 2000-500 | 800-1700 | 800-1700 |
| Длительность АФ,ч | 7 | 8 | 6 | 9 | 9 | 9 |
| Середина АФ,ч | 1030 | 1000 | 2300 | 030 | 1230 | 1230 |
| АА,‰ | 12 | 13.6 | 54.75 | 38.17 | 32.4 | 29.2 |
| ОА,‰ | 5.38 | 5.25 | 2 | 1.7 | 3.8 | 3.9 |
| КС,1/ч | 0.59 | 0.87 | 0.33 | 0.11 | 0.63 | 0.65 |
| РМср,‰ | 54.25±7.7 | 1636±34 | 196±44 | |||
| РМАФср,‰ | 26.25±0.6 | 654±84.1 | 99.8±7 | |||
Введение пептидов эпифиза эпифизэктомированным животным в
6 часов также привело к формированию циркадианного ритма пролиферации эпителия пищевода. Циркадианный ритм МИ эпифизэктомированных животных после введения пептидов эпифиза в 6 часов имел характер монофазного ритма с максимумом деления клеток в 21 и минимумом в 9 часов (Р<0,001). Возрастание митотической активности происходило в ночные часы. Длительность активной фазы ритма восстановилась до уровня близкого к показателям животных контрольной группы. Акрофаза ритма наблюдалась в начале темновóго периода фоторежима. Смещение активной фазы ритма по отношению к таковой интактных животных составило 10 часов. Смещение акрофазы – 12 часов. Значения амплитудных показателей и коэффициента синхронизации ритма эпифизэктомированных животных после введения пептидов эпифиза в 6 часов были близкими к таковым интактных животных. Кривая динамики МИ эпителия пищевода эпифизэктомированных животных, которым вводили пептиды эпифиза в 6 часов имеет вид синусоиды инвертированной по отношению к таковой в контроле.
Результаты многочисленных исследований последних лет свидетельствуют о высокой биологической активности пептидного препарата эпифиза эпиталамина в отношении нейроэндокринной, репродуктивной и иммунной систем. Неоднократно демонстрировалось ингибирующее влияние пептидов эпифиза на функциональную активность гонад, коры надпочечников, гипофиза. Физиологические эффекты мелатонина и пептидов эпифиза обнаруживают частичное функциональное сходство, в частности, отмечается их антигонадотропное, противоопухолевое, иммуномодулирующее и антиоксидантное действие (Ryzhak et al., 2007; Kozina et al., 2007; Konturek et al., 2008). В связи с тем, что физиологические эффекты мелатонина и пептидов эпифиза зачастую оказываются сходными, неоднократно высказывалось предположение о возможности реализации физиологических свойств эпифизарных пептидов через изменение синтеза мелатонина. Изучение характера синтеза мелатонина у крыс, самок макак резусов и человека показало, что введение в вечернее время эпиталамина, а также эпиталона (синтетического аналога эпиталамина), оказывает значительное стимулирующее действие на синтез мелатонина (Korkushko et al., 2004, 2007). Однако приведенные выше результаты собственного исследования, многообразие пептидов эпифиза, а также дистантность их действия и функциональная специфичность свидетельствуют о проявлении пептидами свойств гормонов. Восстановление циркадианного ритма пролиферации обновляющихся тканей эпифизэктомированных животных после введения пептидов эпифиза указывает на то, что пептиды эпифиза обладают собственными ритмогенными свойствами, не связанными с влиянием на продукцию мелатонина.
Временнáя организация пролиферации обновляющихся тканей эпифизэктомированных беспородных белых крыс после совместного введения пептидов эпифиза и мелатонина. Совместное введение пептидов эпифиза и мелатонина эпифизэктомированным животным обуславливает формирование монофазного циркадианного ритма пролиферации эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов. По большинству параметров суточного ритма МИ экспериментальные животные незначительно отличаются от интактных. Длительность активной фазы ритма, а также величины амплитудных показателей ритма (коэффициент синхронизации, абсолютная и относительная амплитуды) не выходили за пределы значений для интактных животных. Суточный пул делящихся клеток не имел статистически достоверных отличий (Р>0,05) от аналогичного показателя интактных животных. Посредством спектрального анализа и анализа методом наименьших квадратов показано наличие циркадианного (период около 24 часов) и ультрадианного (период 7-9 часов) ритмов пролиферации обновляющихся тканей эпифизэктомированных животных, одновременно получавших пептиды эпифиза и мелатонин.
Суточная динамика пролиферации эпифизэктомированных животных, которым одновременно вводили пептиды эпифиза и мелатонин сходна с суточной динамикой пролиферации эпифизэктомированных животных, которым вводили только пептиды эпифиза. Это выражается в наличии монофазного циркадианного ритма клеточной репродукции с близкими периодами и подтверждает ритмогенные свойства пептидов эпифиза.
- Исследование роли эпифиза в формировании биоритмов
продукции тканевых регуляторов пролиферации
Временнáя организация антирадикальной активности кейлон-антикейлонной системы печени интактных беспородных белых крыс. Ритмичность функционирования пролиферативной системы контролируется на различных уровнях, но действие любого фактора в конечном итоге реализуется посредством тканевых регуляторов, на что указывает наличие органной и тканевой специфичности ритмов пролиферации. На тканевом уровне деление клеток контролируется кейлон-антикейлонной системой, в которой кейлоны подавляют, а антикейлоны стимулируют размножение клеток. Соотношение их концентраций в ткани определяет уровень её пролиферативной активности и обеспечивает структурный гомеостаз.
Формирование биоритмов пролиферации осуществляется посредством ритмической продукции тканевых регуляторов деления клеток. Продукция кейлонов в тканях различной локализации характеризуется циркадианной ритмичностью. Показано, что изменения концентрации эпидермального G2-кейлона в эпителии языка и пищевода крыс носят выраженный ритмический характер с активной фазой ритма в дневное время. Наличие циркадианного ритма биосинтеза кейлонов отмечается также в препаратах асцитной опухоли Эрлиха (Антохин, 1979; Машанова, 2003), причем ритмический характер продукции свойственен всем компонентам (G1-, S- и G2-) кейлонсодержащего препарата. Продукция факторов роста также характеризуется циркадианной ритмичностью (Haus et al., 1999).
Антирадикальная активность кейлон-антикейлонной системы печени крыс изучалась на протяжении одних суток. Данные спектрального анализа и анализа методом наименьших квадратов свидетельствуют о ритмической организации антирадикальной активности кейлон-антикейлонной системы печени интактных животных с периодом, приближающимся к 24 и 7 часам.
У интактных животных наблюдалось периодическое изменение антирадикальной активности кейлон-антикейлонной системы печени, которое характеризовалось монофазным ритмом на протяжении суток. Его активная фаза приходилась на ночные часы. Значения антирадикальной активности кейлон-антикейлонной системы печени были максимальными в 21 час, достоверно превышая минимальные значения в 14 часов (Р<0,05). Изменения антирадикальной активности кейлон-антикейлонной системы печени интактных животных скоррелированы с режимом освещения (табл.4). Антирадикальная активность в темновой фазе фотопериода превышала (Р<0,05) таковую в световой фазе (рис.11).
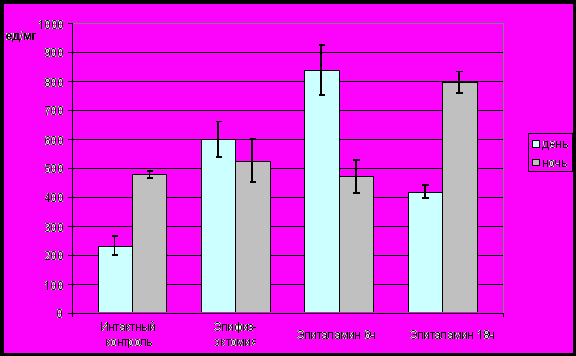
Рис. 11. Средние значения антирадикальной активности (усл. ед/мг белка) кейлон-антикейлонной системы печени в световую и темновую фазы фотопериода
С целью выявления роли эпифиза в формировании циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени была проведена экстирпация железы и изучена динамика антиради-
кальной активности кейлон-антикейлонной системы печени эпифизэктомированных беспородных белых крыс. Эпифизэктомия привела к исчезновению циркадианного ритма антирадикальной активности кейлон-антикей-лонной системы печени. Результаты спектрального анализа и анализа методом наименьших квадратов показали отсутствие циркадианного ритма. В то же время обнаружился выраженный ультрадианный ритм антирадикальной активности с периодом около 9,5 часов.
Колебания антирадикальной активности на протяжении суток не имели какой-либо связи с фоторежимом. На общем монотонном фоне изменений антирадикальной активности наблюдалось два статистически недостоверных подъема (Р>0,05) с максимумами в 24 и 9 часов. Антирадикальная активность кейлон-антикейлонной системы печени в световой и темновой фазах фотопериода не имела статистически достоверных (Р>0,05) отличий (рис.11).
Таким образом, удаление эпифиза привело к рассогласованию циркадианной составляющей деятельности кейлон-антикейлонной системы печени с условиями освещенности, и исчезновению циркадианного ритма продукции тканевых регуляторов пролиферации на 40-е сутки после эпифизэктомии. У эпифизэктомированных животных динамика антирадикальной активности характеризуется только ультрадианной ритмичностью.
Для решения вопроса о взаимосвязи организменного (эпифизарного) и тканевого уровней регуляции биоритмов пролиферации было изучено влияние введения пептидов эпифиза эпифизэктомированным животным на формирование циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени. Пептиды эпифиза вводили одной группе эпифизэктомированных животных в 6 часов, другой группе в 18 часов в дозе 2.5 мг/кг ежедневно с 24 по 41 день после эпифизэктомии.
Введение пептидов эпифиза эпифизэктомированным животным как в 6, так и в 18 часов привело к формированию циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени. Посредством спектрального анализа и анализа методом наименьших квадратов были выявлены циркадианный ритм антирадикальной активности с периодом, приближающимся к 24 часам и ультрадианный с периодом около
8 часов.
Циркадианный ритм антирадикальной активности кейлон-антикейлонной системы печени эпифизэктомированных животных после введения пептидов эпифиза имел характер монофазного ритма. Возрастание антирадикальной активности отмечалось в дневное время при введении пептидов эпифиза в 6 часов и в ночное время при их введении в 18 часов. Длительность активной фазы ритма восстановилась до уровня близкого к показателям животных контрольной группы (табл.4). При введении пептидов эпифиза в 6 часов акрофаза ритма наблюдалась в начале световó-го периода фоторежима. Максимум антирадикальной активности отмечался в 9 и минимум в 3 часа (Р<0,05). При введении пептидов эпифиза в
18 часов акрофаза ритма наблюдалась в начале темновóго периода. Максимум антирадикальной активности отмечался в 21 и минимум в 9 часов (Р<0,05). Смещение активной фазы ритма по отношению к таковой интактных животных составило 11 часов при введении пептидов эпифиза в 6 часов и 1 час при их введении в 18 часов. Смещение акрофазы на 12 часов наблюдалось только при введении пептидов эпифиза в 6 часов. Значения амплитудных показателей и коэффициента синхронизации ритма антирадикальной активности эпифизэктомированных животных после введения пептидов эпифиза были близкими к таковым интактных животных (табл.4).
Таблица 4
Параметры циркадианных ритмов антирадикальной активности кейлон-антикейлонной системы печени интактных, эпифизэктомированных и эпифизэктомированных с последующим введением пептидов эпифиза животных
| Параметр | Интактный контроль | Эпифиз-эктомия | Эпиталамин (18 часов) | Эпиталамин (6 часов) |
| Мезор, усл. ед/мг белка | 324.5±46.88 | 541±37 | 605±64 | 628.7±71.5 |
| Акрофаза,ч | 2100 | Ритм отсутст-вует | 2100 | 900 |
| АФ,ч | 1900-500 | 1800-500 | 600-1600 | |
| Длительность АФ,ч | 10 | 9 | 10 | |
| Середина АФ,ч | 2400 | 2230 | 1100 | |
| АА, усл. ед/мг белка | 308.6 | 471 | 607 | |
| ОА, усл. ед/мг белка | 2.55 | 2.25 | 2.5 | |
| КС,1/ч | 0.17 | 0.18 | 0.41 |
У эпифизэктомированных животных, которым вводили пептиды эпифиза среднесуточные значения антирадикальной активности не имели статистически достоверных отличий (Р>0,05) от значений соответствующих показателей для интактных и эпифизэктомированных животных. В то же время динамика антирадикальной активности, в отличие от эпифизэктомированных животных, характеризовалась наличием циркадианного ритма. Антирадикальная активность в световой фазе фотопериода (Р<0,05) превышала таковую в темновой фазе при введении пептидов эпифиза в 6 часов (рис.11). Введение пептидов эпифиза в 18 часов обусловило превышение (Р<0,05) антирадикальной активности в темновой фазе фотопериода (рис.11). Результаты графически-параметрического анализа указывают на сходство биоритмологической организации кейлон-антикейлонной системы печени интактных и эпифизэктомированных животных, получавших пептиды эпифиза в 18 часов. Введение пептидов эпифиза эпифизэктомированным животным в 6 часов привело к формированию инвертированного циркадианного ритма антирадикальной активности.
III. Оценка биологической активности кейлон-антикейлонных препаратов интактных, эпифизэктомированных и эпифизэктомированных с последующим введением пептидов эпифиза беспородных белых крыс
Для оценки биологической активности препарата системы кейлон-антикейлон интактных, эпифизэктомированных и эпифизэктомированных с последующим введением пептидов эпифиза крыс частично гепатэктомированным мышам вводили комплексный экстракт тканевых регуляторов пролиферации, полученный в дневные или ночные часы.
Проверка биологической активности кейлон-антикейлонного препарата, полученного от интактных беспородных белых крыс показала выраженную митозингибирующую активность его дневных фракций и способность ночных фракций усиливать пролиферацию гепатоцитов частично гепатэткомированных мышей. Это свидетельствует о том, что дневные фракции кейлон-антикейлонного препарата печени содержат преимущественно его кейлонный компонент, в то время как ночные фракции – антикейлонный. Дневные и ночные фракции препарата, выделенные у эпифиз-эктомированных животных не оказывают влияние на репродукцию клеток.
Введение пептидов эпифиза эпифизэктомированным белым крысам привело к восстановлению циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени. Его активные фазы отмечались в разное время суток в зависимости от времени введения пептидов эпифиза. Исследование биологической активности дневных и ночных фракций кейлон-антикейлонного препарата, полученного из печени эпифизэктомированных крыс, которым вводили пептиды эпифиза в 6 часов показало, что введение частично гепатэктомированным мышам ночной фракции препарата приводит к уменьшению МИ гепатоцитов, а введение его дневной фракции повышает митотическую активность гепатоцитов, оказывая инвертированное действие по сравнению с препаратами, полученными от интактных животных. Введение препаратов, полученных от эпифизэктомированных беспородных белых крыс, которым вводили пептиды эпифиза в 18 часов, оказывает аналогичное действие с препаратами, полученными от интактных животных.
Кейлон-антикейлонная система печени белых крыс является динамической системой, в которой на протяжении суток поддерживается баланс между стимулятором и ингибитором пролиферации. В норме у белых крыс баланс между кейлоном и антикейлоном в ночные часы смещен в сторону последнего, что приводит к усилению митотической активности. У эпифизэктомированных крыс происходит исчезновение циркадианного ритма продукции тканевых регуляторов пролиферации, что определяет аналогичность эффектов введения, как ночных, так и дневных фракций препарата на пролиферацию. Введение пептидов эпифиза эпифизэктомированным животным привело к восстановлению циркадианного биоритма продукции тканевых регуляторов пролиферации. В результате эффект введения дневных и ночных фракций препарата кейлона и антикейлона приобретает сходство с эффектом введения такового, полученного от интактных животных. Последнее свойственно для кейлон-антикейлонного препарата, полученного от эпифизэктомированных животных с введением пептидов эпифиза в 18 часов.
Наши исследования показывают, что реализация ритмогенных эффектов эпифиза осуществляется через тканевые регуляторы пролиферации. Это подтверждается исчезновением циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени после эпифизэктомии и его восстановлением в результате введения эпифизэктомированным животным пептидов эпифиза. В то же время возникновение ультрадианных биоритмов пролиферации связано, по-видимому, с цикличностью метаболических процессов в тканях и ультрадианным ритмом продукции тканевых регуляторов пролиферации. Это подтверждается тем, что у эпифизэктомированных животных динамика антирадикальной активности характеризуется только ультрадианной ритмичностью. Тот факт, что введение пептидов эпифиза эпифизэктомированным животным в момент смены фоторежима с темновой фазы на световую (6 часов) привело к формированию инвертированного циркадианного ритма антирадикальной активности кейлон-антикейлонной системы печени указывает на то, что продукция эпифизом биологически активных пептидов, контролирующих ритмичность функционирования кейлон-антикейлонной системы, должна происходить в ночные часы.
Таким образом, система регуляции пролиферативных процессов представляет собой многоуровневый комплекс, в основе которого лежит регуляция деления отдельной клетки. На основании результатов собственного исследования и принимая во внимание данные литературы, можно заключить, что формирование циркадианного ритма пролиферации обеспечивается участием следующих структур: 1) супрахиазматические ядра, воспринимающие информацию от сетчатки глаза о режиме освещения и задающие ритм секреторной активности эпифиза; 2) эпифиз, трансформирующий нервный импульс в эндокринный сигнал; 3) периферические эндокринные железы, модулирующие параметры ритма; 4) эндогенные тканевые регуляторы пролиферации – кейлоны и антикейлоны, формирующие биоритм пролиферации конкретной ткани; 5) внутриклеточные регуляторы пролиферации.
Эпифизарная регуляция биоритмов пролиферации заключается в формировании сезонного и циркадианного ритмов митотической активности и не затрагивает ультрадианную составляющую. Действие биологически активных веществ эпифиза на пролиферацию может быть опосредовано периферическими эндокринными железами и кейлон-антикейлонной системой, а также осуществляться непосредственным воздействием на ткань. В последнем случае мелатонин задерживает клетки в G1- или
G2-периодах клеточного цикла. Продукция эпифизом мелатонина носит циркадианный характер. Концентрация мелатонина в крови достигает максимума к середине ночи, что соответствует пассивной фазе ритма пролиферации. Увеличение митотической активности начинается, как правило, во второй половине темновóго периода, когда наблюдаются падение активности ферментных систем эпифиза и снижение уровня мелатонина в крови. Действие биологически активных пептидов эпифиза на формирование циркадианного ритма пролиферации осуществляется через кейлон-антикейлонную систему. Продукция тканевых регуляторов пролиферации характеризуется циркадианной ритмичностью с активной фазой ритма продукции кейлонов в дневные и антикейлонов в ночные часы.
Таким образом, механизм формирования эпифизом циркадианного ритма пролиферации может быть следующим (рис.12):
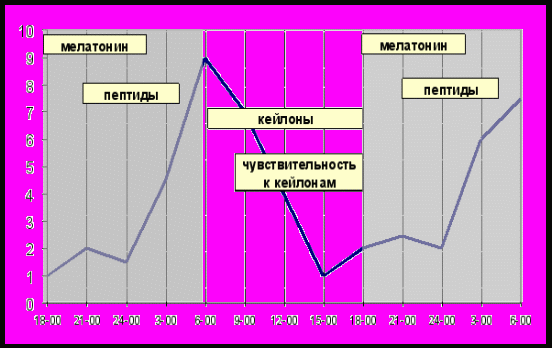
Рис.12. Фазограмма циркадианных ритмов продукции биологически активных веществ эпифизом, концентрации кейлона в ткани и чувствительности клеток к действию кейлона. (Отрезками показаны активные фазы ритма. — суточная динамика пролиферации ткани)
Выделяющийся с наступлением темноты мелатонин, обладает цитостатическим эффектом и тормозит пролиферацию клеток. Вместе с тем, он повышает проницаемость клеточных мембран пинеалоцитов для синтезированных в них олигопептидов, облегчая тем самым попадание последних в кровоток. Выделяющиеся пептиды эпифиза подавляют продукцию кейлонов, изменяя баланс кейлон-антикейлонной системы ткани в пользу последних. На фоне прекращения синтеза мелатонина и повышения уровня антикейлонов во второй половине темновóго периода происходит увеличение митотической активности ткани. В дальнейшем при отсутствии мелатонина и пептидов эпифиза повышается продукция кейлонов, что вызывает снижение пролиферативной активности. Последующий синтез биологически активных веществ эпифизом в ночные часы приводит к повторению описанных выше процессов в результате чего и происходит формирование циркадианного ритма пролиферации.
Выводы
- Разработанный способ анатомического доступа к эпифизу беспородных белых крыс и основанный на нем метод эпифизэктомии является оптимальным для использования в хронобиологических исследованиях.
- Эпифизэктомия вызывает исчезновение циркадианного ритма пролиферации эпителиев пищевода, крипт тощей кишки и клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов беспородных белых крыс и не влияет на ультрадианный биоритм митотической активности указанных тканей.
- Однократное в течение суток введение мелатонина эпифизэктомированным беспородным белым крысам в начале темновóго периода в течение 14 суток не привело к восстановлению циркадианного ритма пролиферации эпителиев пищевода и крипт тощей кишки.
- Эпиталамин, как препарат комплекса биологически активных пептидов эпифиза восстанавливает циркадианный ритм пролиферации обновляющихся тканей (эпителиев пищевода, тощей кишки и клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов беспородных белых крыс) после эпифизэктомии.
- В результате эпифизэктомии у беспородных белых крыс происходит исчезновение циркадианного ритма продукции тканевых регуляторов пролиферации – кейлонов и антикейлонов с сохранением ультрадианного ритма их выработки.
- Введение биологически активных пептидов эпифиза (эпиталамина) эпифизэктомированным беспородным белым крысам приводит к восстановлению циркадианного ритма продукции тканевых регуляторов пролиферации (кейлонов и антикейлонов).
- Регуляция циркадианного ритма митотической активности осуществляется при участии пептидов эпифиза, которые формируют циркадианный ритм продукции тканевых регуляторов пролиферации.