
«Российский государственный медицинский университет Федерального агентства по здравоохранению и социальному развитию»
| Вид материала | Автореферат |
- «Российский государственный медицинский университет Федерального агентства по здравоохранению, 975.58kb.
- «Российский государственный медицинский университет Федерального агентства по здравоохранению, 545.1kb.
- «Сибирский государственный медицинский университет Федерального агентства по здравоохранению, 927.55kb.
- Кубанский государственный медицинский университет федерального агентства по здравоохранению, 821.27kb.
- «Казанский государственный медицинский университет Федерального агентства по здравоохранению, 863.64kb.
- «Саратовский государственный медицинский университет имени В. И. Разумовского Федерального, 378.09kb.
- Гепатопротекторные свойства флавоноидов (фармакодинамика и перспективы клинического, 671.3kb.
- «ультразвуковая диагностика кишечной непроходимости» 14. 00. 27 хирургия, 401.81kb.
- «Рязанский государственный медицинский университет имени академика И. П. Павлова Федерального, 241.22kb.
- Программа подготовки аспирантов по кардиологии составили: д м. н., профессор, 643.17kb.
Результаты и их обсуждение
- Исследование роли эпифиза в формировании биоритмов
пролиферации обновляющихся тканей
Временнáя организация пролиферации обновляющихся тканей интактных беспородных белых крыс. Важным моментом оценки влияния биологически активных веществ эпифиза на пролиферацию является изучение ритма деления клеток интактных животных и животных, освобожденных от каких-либо воздействий со стороны эпифиза. Суточная динамика пролиферации эпителия пищевода интактных беспородных белых крыс представлена на рисунке 3.
У интактных животных наблюдалось периодическое изменение МИ эпителия пищевода, которое характеризовалось монофазным ритмом на протяжении суток. Результаты графически-параметрического анализа динамики МИ указывают на то, что изменения МИ интактных животных скоррелированы с режимом освещения.
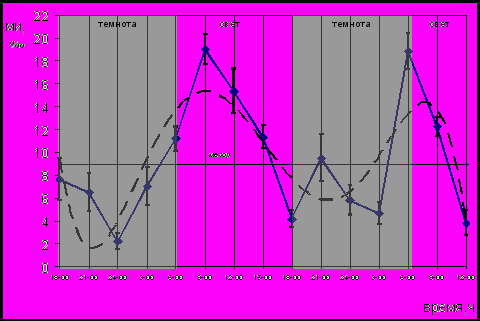
Рис. 3. Динамика МИ эпителия пищевода интактных беспородных белых крыс; (― МИ, ---- сглаженная кривая)
Аналогичный характер присущ суточной динамике пролиферации эпителия крипт тощей кишки и клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов интактных беспородных белых крыс. Однако имеется органная специфика пролиферативных процессов, которая выражается в различии акрофаз ритма, абсолютной и относительной амплитуд, коэффициента синхронизации и ряда других параметров циркадианного ритма пролиферации (табл.2).
Отличие в положении акрофаз циркадианного ритма пролиферации различных обновляющихся тканей указывает на определенную лабильность параметров циркадианного ритма пролиферации. Положение акрофаз выявленных ритмов не является строго фиксированным и может смещаться в пределах 24-х часовой шкалы. Промежуток смещения акрофаз в пределах временнóй шкалы предложено называть “зоной блуждания акрофазы”. Наиболее выраженные изменения положения акрофазы ритма отмечаются в процессе роста и развития организма, а также во время смены сезонов года. Тем не менее, взаимосвязь подъема митотической активности с определенным временем суток сохраняется, что было продемонстрировано нами для ряда обновляющихся тканей. Для максимума митотической активности эпителиев пищевода и тощей кишки, а также клеток герминативного центра лимфатических узлов расположение “зоны блуждания” приходится на вторую половину темновóго - начало световóго периода суток. Таким образом, циркадианный биоритм пролиферации характеризуется лабильностью в определенной “зоне блуждания”, которая соответствует условиям фоторежима.
Таблица 2
Параметры циркадианных ритмов МИ эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов интактных беспородных белых крыс
| Параметр | Пищевод | Кишечник | Лимфатический узел | |||
| 1 сутки | 2 сутки | 1 сутки | 2 сутки | 1 сутки | 2 сутки | |
| Мезор,‰ | 9.27±1.36 | 61.4±2.2 | 25.8±1.68 | |||
| Акрофаза,ч | 900 | 600 | 2100 | 300 | 900 | 900 |
| АФ,ч | 300-1700 | 300-1100 | 1800-300 | 2400-600 | 500-1200 | 500-1200 |
| Длительность АФ,ч | 14 | 8 | 9 | 6 | 7 | 7 |
| Середина АФ,ч | 1000 | 700 | 2230 | 300 | 1230 | 1230 |
| АА,‰ | 16.8 | 15 | 22 | 25.8 | 22.24 | 20.62 |
| ОА,‰ | 8.63 | 4.94 | 1.39 | 1.49 | 2.38 | 2.11 |
| КС,1/ч | 0.95 | 0.82 | 0.15 | 0.16 | 0.39 | 0.35 |
| РМср,‰ | 75.21±12.7 | 1466.7±29.7 | 205.7±50.2 | |||
| РМАФср,‰ | 45±9.8 | 530.15±116.14 | 68.7±0.2 | |||
Примечание: АФ - активная фаза ритма, АА - абсолютная амплитуда, ОА - относительная амплитуда, КС - коэффициент синхронизации МИ в ритме,
РМср - пул делящихся клеток в течение суток (среднее за двое суток),
РМАФср - пул делящихся клеток в течение АФ ритма (среднее за двое суток).
Данные спектрального анализа и анализа методом наименьших квадратов динамик МИ свидетельствуют о ритмической организации митотической активности эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов интактных животных с периодом, приближающимся к 24 часам. Кроме того, во всех указанных тканях выявлен ультрадианный ритм пролиферации c периодом около 9 часов. Совокупность биологических ритмов функций организма с различными периодами является одним из видов временнóй организации живых систем (Романов, 1995). Вероятно, первичными следует считать эндогенные ультрадианные ритмы. Внешние синхронизаторы, координируя работу регуляторных систем, формируют на их основе циркадианные ритмы. Возникновение ультрадианных биоритмов обусловлено цикличностью метаболических процессов в ткани (Бродский и др., 2006).
Временнáя организация пролиферации обновляющихся тканей беспородных белых крыс после эпифизэктомии. Эпифиз является необходимым звеном формирования циркадианных ритмов ряда функций организма. Он осуществляет преобразование нервных импульсов в гормональный сигнал. В системе управления циркадианными ритмами комплекс “супрахиазматические ядра – эпифиз” обеспечивает гармоничную адаптацию к меняющимся средовым факторам, в частности, к свету. Результаты нашего эксперимента подтверждают упомянутую функциональную схему, свидетельствуя о важной роли эпифиза в фотопериодическом контроле циркадианного ритма пролиферативной активности тканей организма. С целью выявления роли эпифиза в регуляции циркадианного ритма пролиферации была проведена экстирпация железы и изучена динамика МИ обновляющихся тканей эпифизэктомированных животных.
Эпифизэктомия привела к исчезновению циркадианного ритма пролиферации всех изучавшихся тканей. Результаты спектрального анализа и анализа методом наименьших квадратов показали отсутствие циркадианного ритма МИ. В то же время обнаружился выраженный ультрадианный ритм репродукции клеток с периодом от 7 до 13 часов.
На рисунке 4 представлена суточная динамика пролиферации эпителия пищевода эпифизэктомированных беспородных белых крыс. Колебания МИ на протяжении двух суток не обнаружили какой-либо связи с фоторежимом. Уровень пролиферации эпителия пищевода эпифизэктомированных животных был таким же, как у интактных, что выражалось в отсутствии различий (Р>0,05) между: 1) среднесуточными значениями МИ (8,45±0,97‰ и 9,27±1,36‰); 2) суточными пулами пролиферирующих клеток (66,4±16,2‰ и 75,2±12,7‰) животных обеих групп.
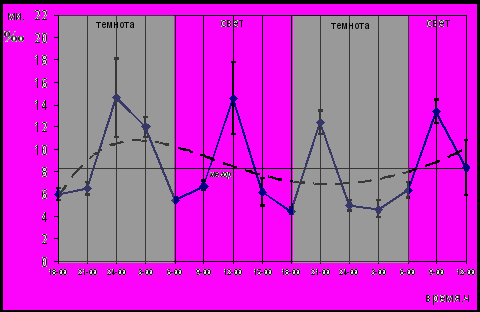
Рис. 4. Динамика МИ эпителия пищевода эпифизэктомированных беспородных белых крыс; (― МИ, ---- сглаженная кривая)
Аналогичный характер присущ суточной динамике пролиферации эпителия крипт тощей кишки и клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов. Пролиферативная активность эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатических узлов в световой и темновой фазах фотопериода не имела статистически достоверных (Р>0,05) отличий. Таким образом, удаление эпифиза привело к рассогласованию циркадианной составляющей деятельности пролиферативной системы с условиями освещенности, и исчезновению циркадианного ритма пролиферации всех изученных тканей на 40-е сутки после эпифизэктомии. У эпифизэктомированных животных динамика митотической активности характеризуется наличием только ультрадианного ритма, периоды которого составили 11 часов в эпителии пищевода, 9 часов в эпителии крипт тощей кишки, 7 и 13 часов для клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов. Это указывает на важную роль эпифиза в фотопериодическом контроле циркадианного ритма пролиферации. Регуляторная роль эпифиза установлена нами на различных моделях репродукции клеток. Кроме представленных в работе данных, нами показано исчезновение циркадианного ритма пролиферации сперматогоний беспородных белых крыс после эпифизэктомии (Слесарева и др., 2005).
Эпифизэктомия не вызывает морфологических изменений эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов (рис.5-7) беспородных белых крыс.
Исчезновение циркадианного ритма пролиферации у эпифизэктомированных животных позволяет предполагать участие биологически активных веществ эпифиза в его формировании. В настоящее время ритмогенную функцию эпифиза связывают с продукцией мелатонина, синтез и секреция которого имеет циркадианный характер (Кветная и др., 2005). Способность мелатонина подавлять пролиферацию различных тканей отмечена как в культуре, так и in vivo (Blask et al., 2005; Pizarro et al., 2008). Наличие рецепторов к мелатонину продемонстрировано для большинства клеток тканей организма (Korf et al., 2006). Однако вопрос о роли мелатонина в регуляции циркадианного ритма пролиферации продолжает оставаться дискуссионным.
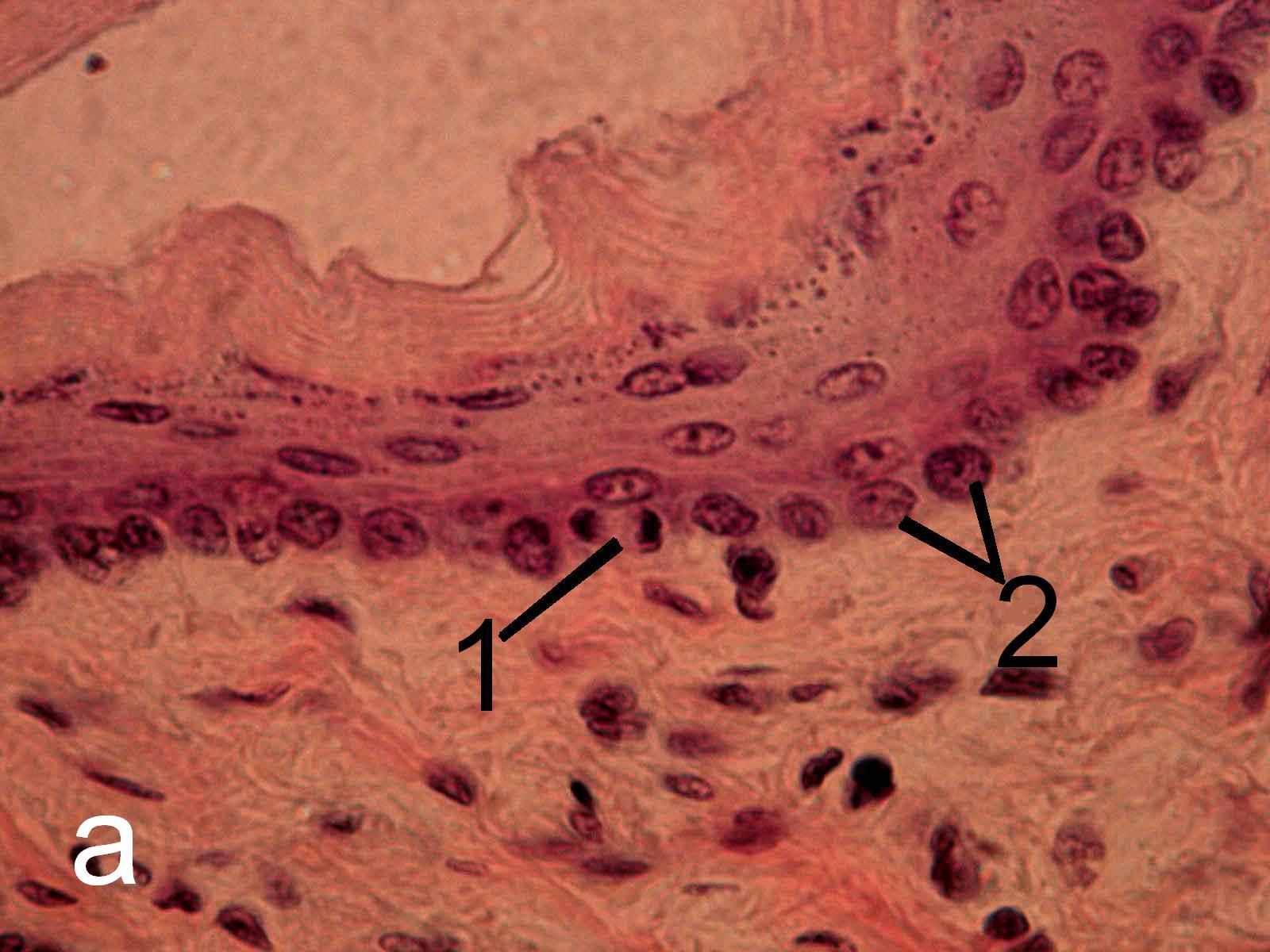
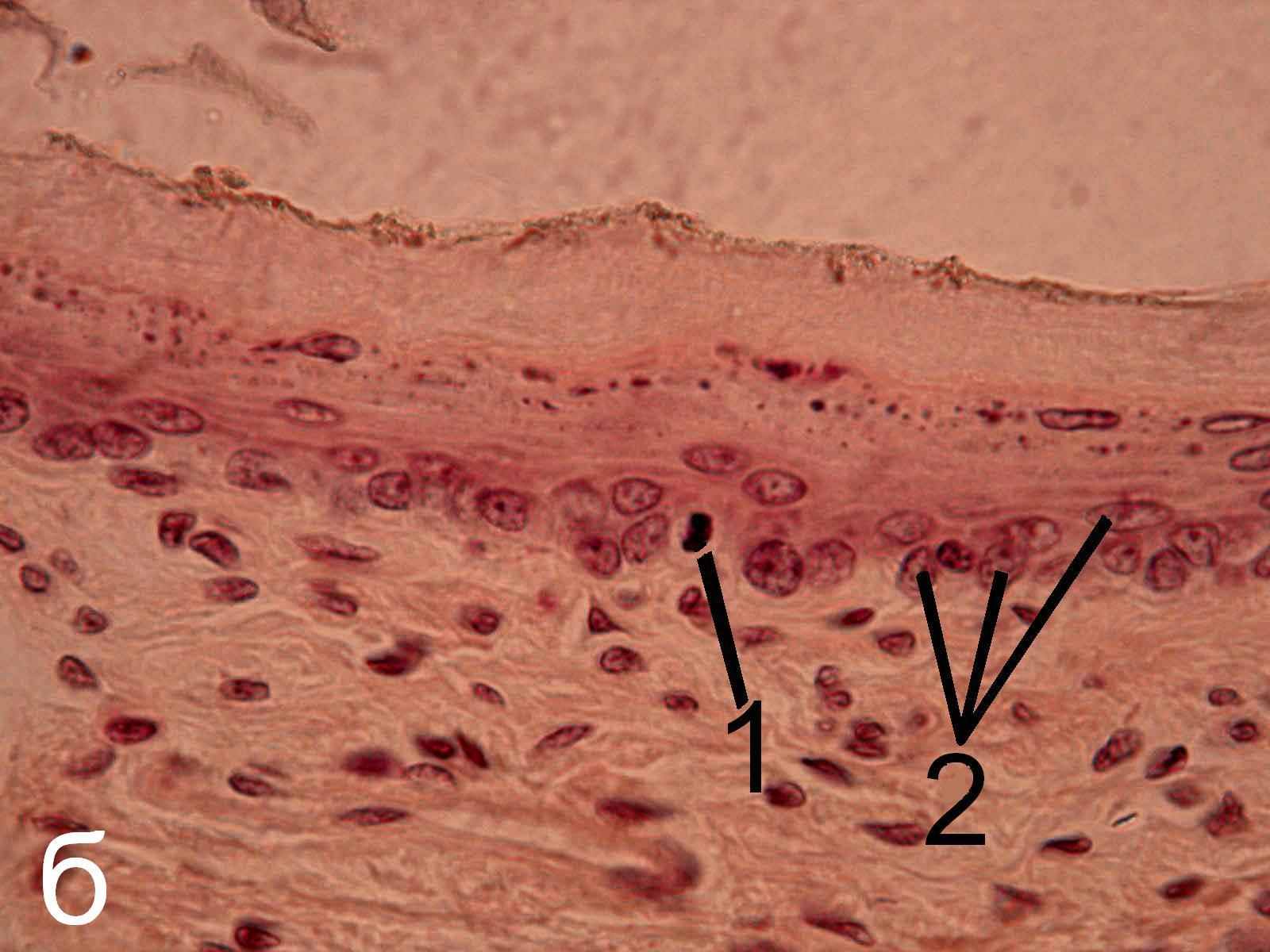
Рис. 5. Многослойный плоский неороговевающий эпителий пищевода интактных (а) и эпифизэктомированных (б) беспородных белых крыс. Окраска гематоксилин-эозином. х1000. 1 – фигура митоза, 2 – ядра клеток,
находящихся в интерфазе.
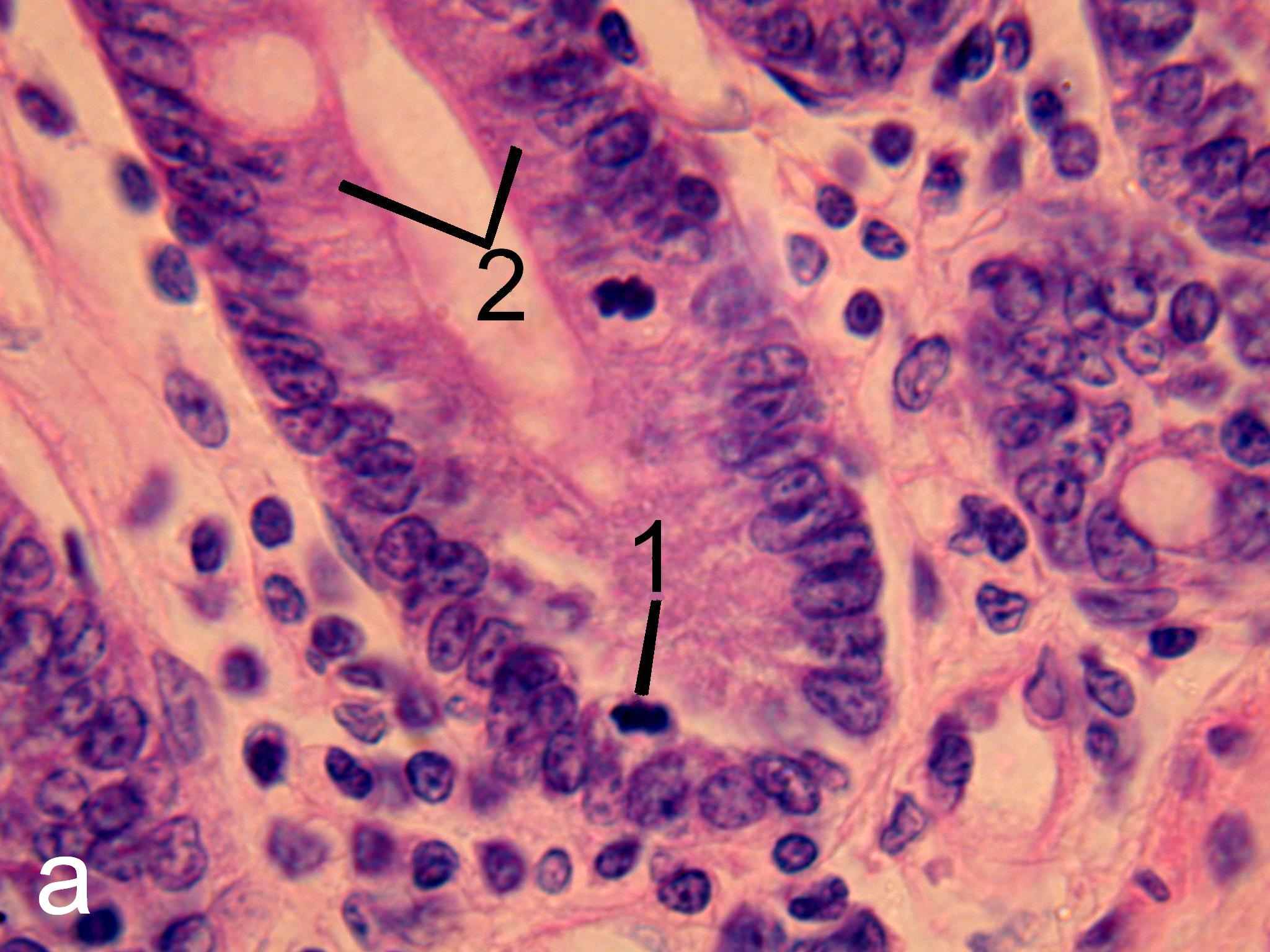
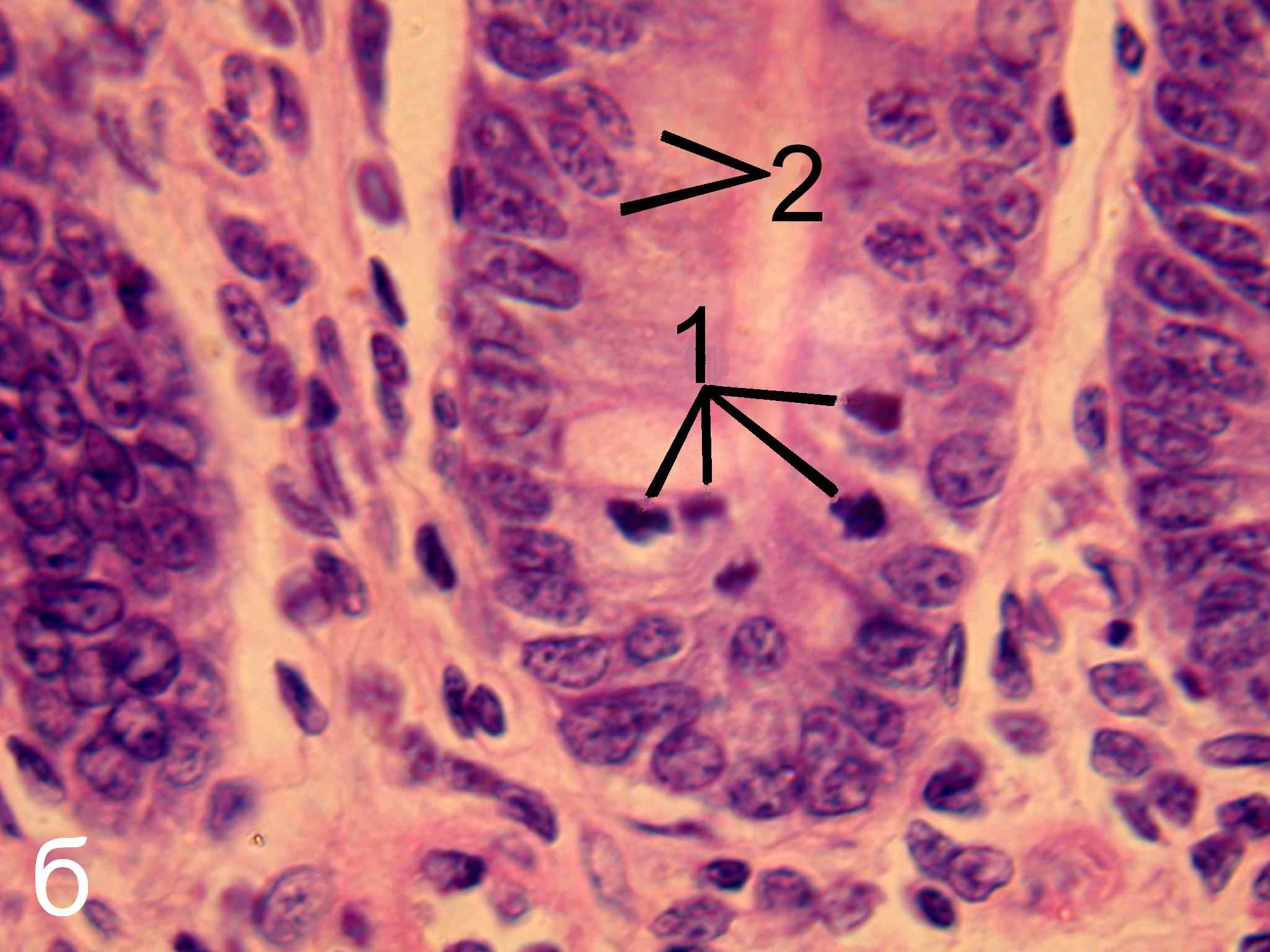
Рис. 6. Эпителий крипт тощей кишки интактных (а) и эпифизэктомированных (б) беспородных белых крыс. Окраска гематоксилин-эозином. х1000.
1 – фигуры митоза, 2 – клетки, находящиеся в интерфазе.
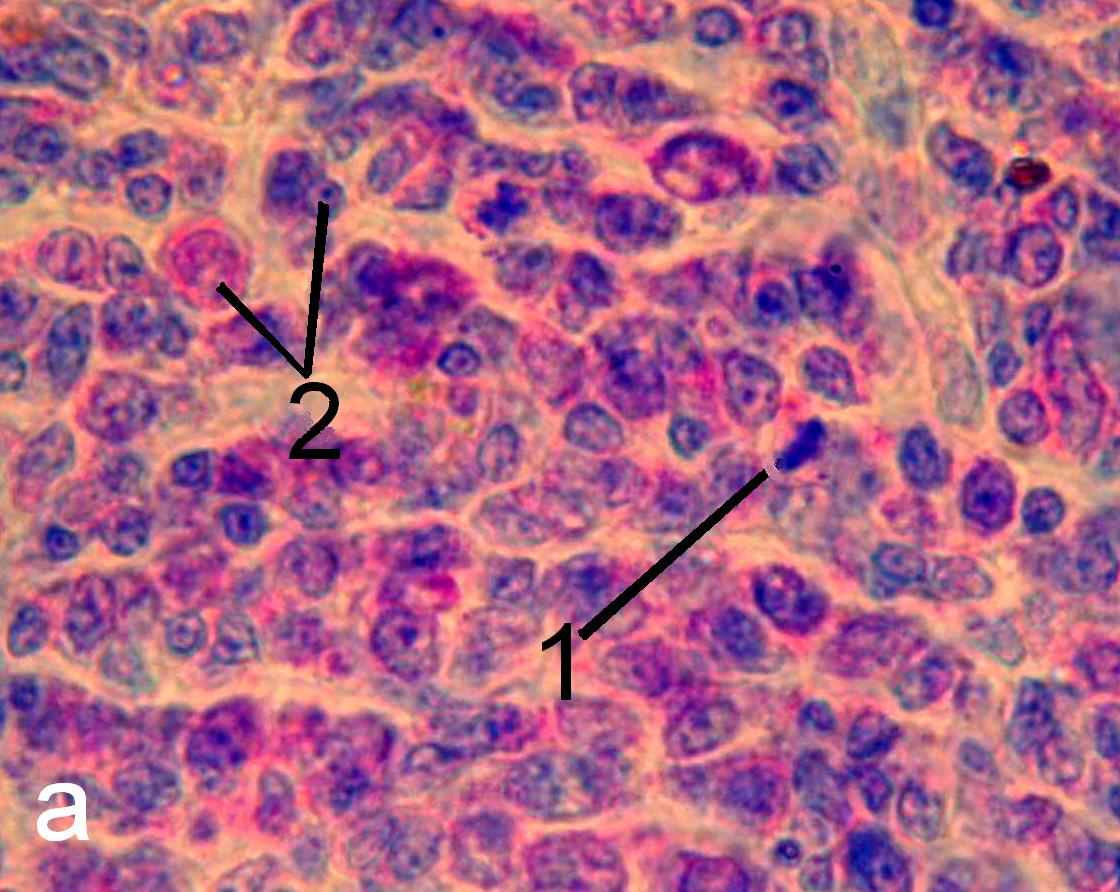
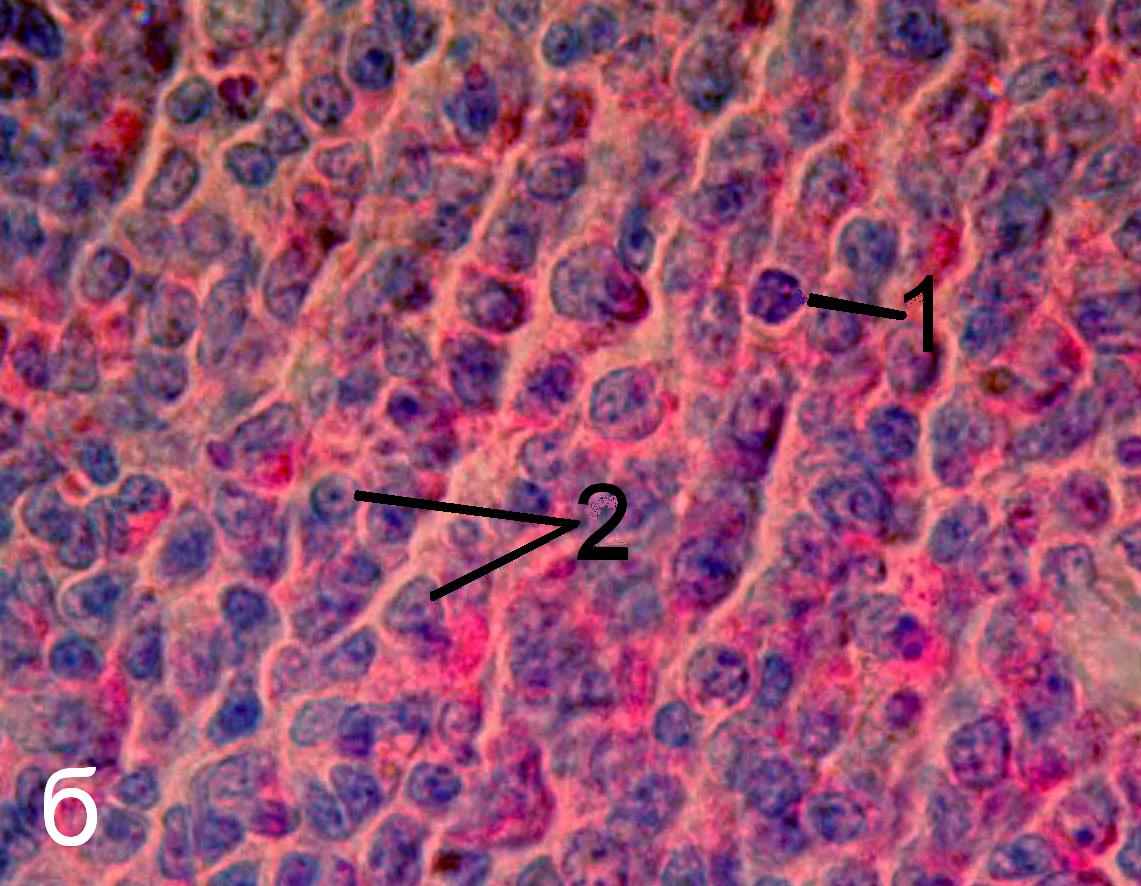
Рис. 7. Герминативный центр лимфатического узелка трахеобронхиально-го лимфатического узла интактных (а) и эпифизэктомированных (б) беспородных белых крыс. Окраска пиронином-метиловым зеленым по Браше. х1000. 1 – фигура митоза, 2 – ядра клеток, находящихся в интерфазе.
Влияние мелатонина на суточную динамику пролиферации обновляющихся тканей эпифизэктомированных животных. Результаты проведенного исследования указывают на отсутствие ритмогенного эффекта мелатонина при его разовом в течение суток введении эпифизэктомированным животным (10 мг/кг, 18 часов, 14 дней). Суточная динамика пролиферации эпителиев пищевода и крипт тощей кишки характеризовалась только ультрадианными колебаниями с периодом около 12 часов в эпителии пищевода и около 9 часов в эпителии крипт тощей кишки.
На отсутствие циркадианного ритма пролиферации эпителия пищевода эпифизэктомированных животных, которым вводили мелатонин, указывает характер сглаженной кривой динамики МИ (рис.8).
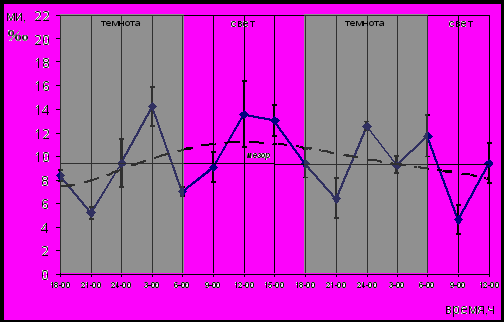
Рис. 8. Динамика МИ эпителия пищевода эпифизэктомированных беспородных белых крыс после введения мелатонина; (― МИ, ---- сглаженная кривая)
По отношению к сглаженной кривой динамики МИ эпителия пищевода интактных животных (рис.3) на протяжении двух суток она утратила вид синусоиды. Увеличение митотической активности (Р<0,05) приходилось как на темновой, так и световой периоды, что указывает на отсутствие синхронизации колебаний митотической активности с суточным свето-темновым циклом. Среднесуточное значение МИ составило 9,5±0,77‰, а суточный пул делящихся клеток – 71,64±34‰. Оба показателя существенно не отличались (Р>0,05) от значений соответствующих показателей интактных и эпифизэктомированных животных.
Однократное в течение суток введение мелатонина эпифизэктомированным животным также не привело к восстановлению циркадианного ритма пролиферации эпителия крипт тощей кишки. Суточная динамика пролиферации эпителия крипт тощей кишки эпифизэктомированных животных после введения мелатонина была аналогична таковой эпителия пищевода. Пролиферативная активность эпителиев пищевода и крипт тощей кишки эпифизэктомированных животных, которым вводили мелатонин, также как у эпифизэктомированных животных, в течение световой и темновой фаз фотопериода не имела достоверных (Р>0,05) отличий. Суточная динамика пролиферации у животных рассмотренных экспериментальных групп соответствует в целом изменениям митотической активности эпифизэктомированных животных и характеризуется ультрадианной ритмичностью.
Введение мелатонина привело к формированию циркадианного ритма пролиферации клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов беспородных белых крыс. Динамика МИ клеток герминативного центра лимфатических узлов эпифизэктомированных животных, которым вводили мелатонин, представлена на рисунке 9.
В отличие от сглаженной кривой суточной динамики МИ эпителиев пищевода (рис.8) и крипт тощей кишки, сглаженная кривая суточной динамики МИ клеток герминативного центра имеет вид синусоиды с высокой амплитудой колебаний. Данные спектрального анализа и анализа методом наименьших квадратов показали, что динамика изменений МИ клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов эпифизэктомированных животных после введения мелатонина характеризовалась колебаниями с циркадианной (период около 24 часов) и ультрадианной (период около 10 часов) составляющими. Значение МИ в светлое время суток (36,58±3‰) было достоверно выше (P<0,05), чем в темное (18±2,1‰). Среднее значение МИ клеток герминативного центра составило 24,29±2,65‰, что не отличалось от уровня пролиферативной активности интактных животных (Р>0,05).
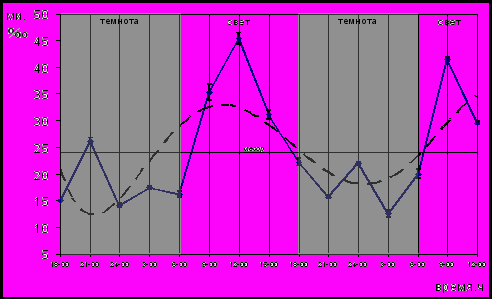
Рис. 9. Динамика МИ клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов эпифизэктомированных беспородных белых крыс после введения мелатонина; (― МИ, ---- сглаженная кривая)
Реакция клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов на эпифизэктомию и введение мелатонина имеет специфические особенности. В отличие от реакции эпителиев пищевода и крипт тощей кишки, эпифизэктомия обусловила увеличение митотической активности клеток герминативного центра. По данным В.Х. Хавинсона (2001) эпифизэктомия вызывает увеличение пролиферативной активности лимфоидных клеток в герминативном центре лимфатических узелков селезенки белых крыс. Это соответствует представлениям об ингибирующем влиянии эпифиза на пролиферативные процессы лимфоидной и сперматогенной тканей. Максимальный уровень пролиферации в органах иммунной системы лабораторных грызунов отмечается на фоне сниженной концентрации глюкокортикоидных гормонов и уменьшения активности ферментных систем эпифиза, приводящей к снижению уровня мелатонина в крови. В темный период суток в плазме крови, с одной стороны, содержится наименьшее количество гормонов тимуса, соматотропного, тиреотропного гормонов, регулирующих пролиферацию и дифференцировку клеток иммунной системы, а с другой, повышена концентрация глюкокортикоидов, оказывающих иммуносупрессивный эффект.
Тот факт, что введение мелатонина эпифизэктомированным животным не привело к восстановлению циркадианного ритма пролиферации эпителиев пищевода и крипт тощей кишки указывает на то, что формирование циркадианного ритма МИ клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов эпифизэктомированных животных с последующей инъекцией мелатонина вызвано не ритмогенным эффектом последнего, а выделением глюкокортикоидов в ответ на процедуру введения гормона. По-видимому, периодические инъекции (в одно и то же время в течение 14 дней) обусловили формирование биоритма функциональной активности надпочечников и щитовидной железы, гормоны которых оказывают влияние на пролиферацию клеток иммунной системы.
В реализации эффектов мелатонина наиболее существенны такие факторы как время суток и сезон года, когда вводится препарат, а также доза, режим и способ введения. В нашем эксперименте введение мелатонина эпифизэктомированным животным производилось непосредственно перед сменой фоторежима со световой фазы на темновую, что должно смоделировать ночной подъем уровня мелатонина у интактных животных. В литературе отмечается, что введение мелатонина экспериментальным животным и человеку в дневные часы приводит, как правило, к нарушению поведенческих реакций, вызывает сонливость и снижение температуры тела.
Важное значение в реализации эффектов мелатонина имеет выбор дозы вводимого препарата. В частности, антипролиферативный эффект действия мелатонина в отношение некоторых типов опухолей не выявлялся при введении его в высоких надфизиологических концентрациях, тогда как в физиологических дозах он тормозил рост новообразований.
Заслуживающим внимания является тот факт, что действие мелатонина зависит не только от уровня гормона в крови, но и от продолжительности его секреции в течение ночи, так как время циркуляции мелатонина в кровотоке не превышает 30 минут. На это указывают также данные о кратковременном (в течение часа) цитостатическом эффекте мелатонина на пролиферацию эпителия роговицы после его однократного введения.
Таким образом, непродолжительная циркуляция мелатонина в кровотоке, с одной стороны, однократное в течение суток введение гормона, с другой, могли стать основными причинами, из-за которых не произошло восстановления циркадианного ритма пролиферации эпителиев пищевода и крипт тощей кишки эпифизэктомированных животных. Отсутствие ритмогенного эффекта мелатонина также продемонстрировано нами при изучении его влияния на пролиферацию сперматогоний эпифизэктомированных беспородных белых крыс (Арав и др., 2006). По-видимому, условием для формирования циркадианного ритма пролиферации является поддержание стабильной концентрации мелатонина в течение всего темновóго периода фоторежима. Отмеченное может рассматриваться обоснованием необходимости разработки форм и способов обеспечения пролонгированного действия мелатонина.
Влияние пептидов эпифиза на суточную динамику пролиферации обновляющихся тканей эпифизэктомированных животных. Суточная динамика продукции биологически активных пептидов эпифизом продолжает оставаться неизученной. В связи с этим при исследовании роли пептидов эпифиза в регуляции биоритмов пролиферации одной группе эпифизэктомированных животных вводили эпиталамин в 18, а второй в
6 часов. В первом случае суточная динамика пролиферации эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов исследовалась в течение двух суток. Изучение влияния утреннего (6 часов) введения пептидов эпифиза эпифизэктомированным животным на восстановление циркадианного ритма пролиферации эпителия пищевода проводилось в течение суток.
Введение пептидов эпифиза эпифизэктомированным животным в
18 часов привело к восстановлению циркадианного ритма пролиферации эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов. На рисунке 10 представлена суточная динамика пролиферации эпителия пищевода эпифизэктомированных беспородных белых крыс после введения пептидов эпифиза.
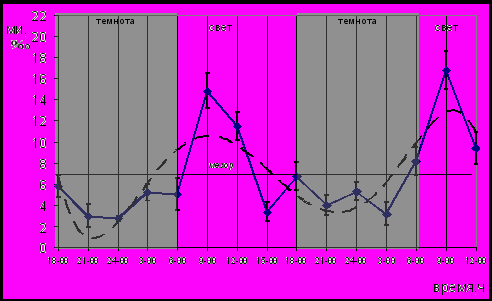
Рис. 10. Динамика МИ эпителия пищевода эпифизэктомированных беспородных белых крыс после введения пептидов эпифиза в 18 часов; (― МИ, ---- сглаженная кривая)
Циркадианный ритм пролиферации эпителия пищевода эпифизэктомированных животных после введения пептидов эпифиза в 18 часов имел характер монофазного ритма. Смещение активной фазы ритма по отношению к таковой интактных животных составило 4 часа в первые сутки и
3 часа во вторые сутки опыта. Смещение акрофазы на 3 часа наблюдалось только во вторые сутки эксперимента. Обращает на себя внимание тот факт, что значения амплитудных показателей ритма и коэффициента синхронизации ритма МИ у эпифизэктомированных животных после введения пептидов эпифиза были выше по сравнению с таковыми интактных животных (табл.2,3).
Аналогичный характер присущ суточной динамике пролиферации эпителия крипт тощей кишки и клеток герминативного центра лимфатического узелка трахеобронхиальных лимфатических узлов эпифизэктомированных с последующим введением пептидов эпифиза беспородных белых крыс.
Посредством спектрального анализа и анализа методом наименьших квадратов были выявлены две составляющие ритма пролиферации всех изучавшихся тканей эпифизэктомированных животных, которым вводили пептиды эпифиза, с периодами, приближающимися к 24 и 9 часам. Особенности суточной динамики пролиферации всех видов тканей обнаруживали сходство с таковыми интактных животных, что подтверждается характером сглаженных кривых и результатами графически-параметрического анализа циркадианных ритмов пролиферации. У эпифизэктомированных животных, которым вводили пептиды эпифиза среднесуточное значение МИ и суточный пул пролиферирующих клеток эпителиев пищевода и крипт тощей кишки, а также клеток герминативного центра не имели статистически достоверных отличий (Р>0,05) от значений соответствующих показателей для интактных животных (табл.2,3).
Несмотря на общность временнóй организации пролиферации изученных тканей, которая выражается в наличии монофазного циркадианного и ультрадианного ритмов МИ, имеется органная специфика пролиферативных процессов, которая выражается в различии акрофаз ритма, абсолютной и относительной амплитуд, коэффициента синхронизации и ряда других параметров (табл.3).
Таблица 3