
Язычная и глоточная (непарные), образующие лимфоидное глоточное кольцо Пирогова Вальдейера, расположены в области зева, корня языка и носовой части глотки
| Вид материала | Документы |
СодержаниеЯзычная миндалина (tопsillа lingualis) Нёбная миндалина (tоnsillа pаlаtiпа) 2 - многослойный плоский эпителий, 3 - лимфоидная ткань миндалины, 4 Трубная миндалина (tоnsillа tubaria) |
- И. Г. Татаринова, преподаватель русского языка Педагогическая деятельность Н. И. Пирогова, 81.58kb.
- Учебного занятия по теме: «Факторы, образующие климат Челябинской области». Базовый, 297.34kb.
- А. Г. Ипполитова отмечала, что "Правильный ротовой выдох при опущенном корне языка, 104.63kb.
- Интеллектуально-речевое развитие обучающихся на уроках русского языка с использованием, 133.71kb.
- Правый желудочек и правое предсердие расположены по центру, поэтому наша речь и поступки, 92.63kb.
- Календарный план проведения занятий по Морфологии растений в лмш-2009 (7 класс), 38.82kb.
- Олимпиада по русскому языку для 5 класса, 145.36kb.
- Лекция I. Понятия «языковая ситуация», 419.51kb.
- Свойств, теорем и неумение их применять незнание приемов решения задач, рассматриваемых, 338.24kb.
- "Новое кольцо" для красавицы столицы, 92.68kb.
Содержание.
1. Строение миндалин 3
2. Функции лимфоэпителиального глоточного кольца 8
3. Список литературы 14
1. Строение миндалин
Миндалины - нёбные и трубные (парные), язычная и глоточная (непарные), — образующие лимфоидное глоточное кольцо Пирогова - Вальдейера, расположены в области зева, корня языка и носовой части глотки. Они представляют собой скопления диффузной лимфоидной ткани, содержащие небольших размеров более плотные клеточные массы - лимфоидные узелки. Эта ссылка скрыта и расположена вдоль дыхательного тракта. Миндалины не относятся к категории лимфоидных органов, поскольку не полностью инкапсулированы. Лимфатические фолликулы разделены на В- и Т-зависимые зоны.
^ Язычная миндалина (tопsillа lingualis) (непарная) залегает в собственной пластинке слизистой оболочки корня языка в виде одного или двух скоплений лимфоидной ткани. Слизистая оболочка над миндалиной образует углубления - крипты, стенки которых образованы многослойным плоским неороговевающим эпителием, инфильтрированным лимфоцитами. Наиболее крупных размеров язычная миндалина достигает у детей и подростков. В этом возрасте язычная миндалина состоит из 80—90 лимфоидных узелков. Начиная с юношеского возраста постепенно количество лимфоидных узелков в язычной миндалине уменьшается. Основными клеточными элементами узелков являются лимфоциты (до 95 - 98%), встречаются также молодые клетки лимфоидной ткани (бласты), располагающиеся в петлях ретикулярной стромы, плазматические клетки, макрофаги.
^ Нёбная миндалина (tоnsillа pаlаtiпа) парная, неправильной формы, располагается в миндаликовой ямке (бухте), которая представляет собой углубление между нёбно-язычной и нёбно-глоточной дужками. Латеральной стороной миндалина прилежит к соединительнотканной пластинке, представляющей собой глоточную фасцию. На медиальной свободной поверхности миндалины видно до 20 миндаликовых отверстий одноименных крипт, являющихся углублениями слизистой оболочки. Одни крипты имеют форму просто устроенных трубочек, другие разветвлены в глубине миндалины. Ширина просвета отдельных крипт равна 0,8 - 1 мм. Слизистая оболочка покрыта многослойным плоским неороговевающим эпителием, который инфильтрирован лимфоцитами.
В диффузной лимфоидной ткани миндалины располагаются плотные скопления лимфоидной ткани округлой или овоидной формы и различных размеров - лимфоидные узелки (рис. 1). Наибольшее количество их наблюдается в возрасте от 2 до 16 лет. К 8—13 годам миндалины достигают наибольших размеров, которые сохраняются примерно до 30 лет. Разрастание соединительной ткани внутри нёбной миндалины особенно интенсивно происходит после 25—30 лет наряду с уменьшением количества лимфоидной ткани. После 40 лет лимфоидные узелки в лимфоидной ткани встречаются редко, размеры оставшихся узелков относительно невелики (0,2—0,4 мм). В крупных лимфоидных узелках виден центр размножения, вокруг узелков находится диффузная лимфоидная ткань. Ретикулярная строма состоит из ретикулярных клеток и волокон, образующих петли, в которых лежат лимфоциты (до 90 - 95 %), плазматические клетки, молодые клетки лимфоидного ряда, макрофаги, гранулоциты.
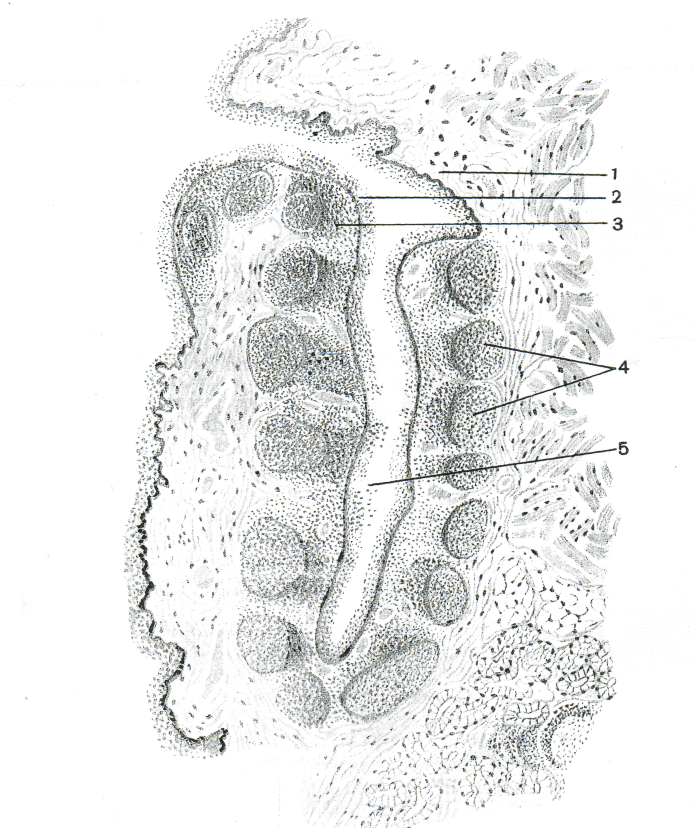
Рис. 1. Строение нёбной миндалины (по И. В. Алмазову и Л. С. Сутулову, 1978): 1 - слизистая оболочка, ^ 2 - многослойный плоский эпителий, 3 - лимфоидная ткань миндалины, 4 - лимфоидные узелки, 5 - просвет крипты
Глоточная миндалина (tоnsillа pharyngealis) непарная, располагается в области свода и отчасти задней стенки глотки между глоточными отверстиями правой и левой слуховых труб. В этом месте имеется 4—6 поперечно и косоориентированных, разделенных бороздами складок слизистой оболочки, внутри которых находится лимфоидная ткань глоточной миндалины. Свободная поверхность складок покрыта многослойным плоским неороговевающим эпителием, в глубине борозд встречаются участки многорядного мерцательного эпителия. Под эпителием в собственной пластинке слизистой оболочки и диффузной лимфоидной ткани находятся лимфоидные узелки глоточной миндалины диаметром до 0,8 мм. Глоточная миндалина достигает наибольших размеров в 8—20 лет, после 30 лет величина ее постепенно уменьшается.
^ Трубная миндалина (tоnsillа tubaria) парная, находится в области трубного валика, ограничивающего сзади глоточное отверстие слуховой трубы. Миндалина представляет собой скопление лимфоидной ткани в собственной пластинке слизистой оболочки слуховой трубы возле ее глоточного отверстия. Миндалина содержит округлой формы единичные лимфоидные узелки. Слизистая оболочка покрыта многослойным плоским неороговевающим эпителием. Трубная миндалина достигает наибольших размеров в возрасте 4—7 лет.
Региональная лимфатическая система лимфоэпителиального глоточного кольца вместе с лимфоцитами печени, пейеровых бляшек тонкого кишечника, лимфоидными фолликулами аппендикса и мочеполового тракта имеет свою собственную в некоторой степени отдельную сеть рециркуляции клеток, которая получила собирательное название - ассоциированная со слизистыми лимфоидная ткань (MALT or mucosal-associated lymphoid tissue).
Лимфоциты MALT формируются в ходе эмбрионального развития аналогично остальным, но их выживание и поддержание мало зависит от центральных органов иммунной системы. Считается, что 50% тканевых лимфоцитов ассоциированы со слизистыми оболочками, что отражает их важную роль как основного места поступления чужеродного материала. По некоторым оценкам, общая площадь слизистых оболочек в 400 раз превышает площадь поверхности тела.
В лимфоидной ткани слизистых оболочек преобладают Т-лимфоциты и содержатся до 30% В-клеток, формирующих зародышевые фолликулы. В собственной пластинке слизистой оболочки преобладают активированные Т-лимфоциты, по преимуществу, хелперы. Здесь же обнаруживаются плазматические клетки, особенностью которых считается специализация на синтез IgA.
Данные иммуноглобулины подвергаются секреции медиаторов воспаления (эндоцитолизу) и проникают в клетки эпителия слизистой оболочки через специальный полиглобулиновый рецептор, затем соединяются с секреторным компонентом эпителиального происхождения, защищающим их от ферментативного расщепления (протеолиза при трансцитозе), и, наконец, экзоцитируются в составе слизи на поверхность слизистой оболочки.
В региональных органах MALT системы происходит встреча антигена, проникающего через слизистые оболочки, со зрелыми иммунокомпентентны-ми клетками-лимфоцитами и развитие иммунного ответа. Реакции иммунитета являются адресными, так как любой рецептор или антитело относительно точно распознает только один антиген. В ходе иммунного ответа происходит преимущественная экспансия и антигензависимая дифференцировка клонов, распознающих минимальные структуры (эпитопы) того или иного антигена.
Ассоциированная со слизистыми оболочками лимфоидная ткань обособлена от остальных периферических лимфоидных органов и способна при иммунных ответах реагировать как целое. Иммуно-компетентные клетки - лимфоциты этой системы постоянно циркулируют через кровоток, т.е. находятся в состоянии рециркуляции. При этом происходит обмен клетками между кровью, лимфой и лимфоидными органами. Благодаря этому, при иммунных ответах данная система реагирует как целое, т.е. местная сенсибилизация лимфоцитов в одном ее участке через некоторое время приводит к иммунному ответу по всей ее площади. Более того, благодаря уникальной модели рециркуляции лимфоцитов, возможны перекрестные ответы с первичной энтеральной сенсибилизацией и иммунным ответом на слизистой оболочки дыхательных путей, как это наблюдается иногда при пищевой аллергии, ассоциированной с бронхиальной астмой. Это в полной мере относится к лимфоэпителиальному глоточному кольцу.
Утрата данных уникальных лимфоэпителиальных структур является невосполнимой потерей для организма, ибо после рождения эти особые популяции лимфоидных клеток не возобновляются, что подчеркивает необходимость взвешенного подхода к выбору метода лечения хронического тонзиллита и аденоидита. В случае хирургического удаления участков такого лимфоэпителиального симбиоза существенно ограничиваются защитные ресурсы соответствующей зоны слизистой оболочки и возможности иммуномодулирующей терапии.
2. Функции лимфоэпителиального глоточного кольца
Миндалины выполняют в организме важную защитную функцию, в них образуются лимфоциты, участвующие в реакциях гуморального и клеточного иммунитета.
Все компоненты лимфоэпителиального глоточного кольца входят в состав единой иммунной системы, формируя иммунологическую резистентность организма. Ее создание осуществляется с участием следующих основных функций лимфаденоидного глоточного кольца:
защитной барьерной функции и местного иммунитета миндалин;
системного иммунного ответа, запущенного путем сенсибилизации лимфоцитов миндалин.
Защитная барьерная функция и местный иммунитет миндалин формируются за счет следующих факторов:
миграции фагоцитов, экзоцитоза и фагоцитоза;
выработки защитных факторов широкого спектра действия;
секреции антител.
Неспецифические факторы защиты лимфоэпителиального глоточного кольца-это:
фагоциты;
секреторные иммуноглобулины;
интерфероны;
а также естественные антибиотики лейкоцитарного происхождения - катионные белки фагоцитов, обладающие бактерицидным и цитотоксическим эффектом (дефензины, лактоферрин и другие).
Большое значение имеют протеолитические ферменты лейкоцитов (протеазы, липазы, гликозидазы, фосфотазы) и, особенно - лизоцим.
Формирование системного иммунного ответа путем антигенной стимуляции лимфоэпителиального глоточного кольца осуществляется в несколько этапов с участием лимфоцитов лимфоэпителиального глоточного кольца и всей системы MALT.
Непосредственное формирование системного иммунитета условно можно представить поэтапно.
Первый этап - Для запуска лимфоцитов в активацию антиген должен быть представлен на поверхности специализированных клеток. Эту функцию выполняют аншиген-презентирующие (или представляющие) клетки (АПК), которые находятся в периферических лимфоидных органах, в том числе региональных лимфоузлах, дренирующих кольцо Вальдейра.
Главная особенность АПК - это способность к поглощению антигена путем той или иной разновидности эндоцитоза, а также к его обработке для предъявления Т-лимфоцитам. Антигеном является молекула, способная вызвать иммунный ответ. Антигенны все те молекулы, которые обладают достаточной пространственной структурой, чтобы участвовать в комплементарных взаимодействиях, распознавать и распознаваться. Иммунный ответ всегда требует кооперации АПК и лимфоцитов, в большинстве случаев, как Т-, так и В-популяций.
Ключевым звеном иммунного ответа является главный комплекс гистосовместимости (ГКГС или HLA). Это высокополимерные мембранные гликопротеиды, участвующие в маркировке собственных клеток и распозновании «своего» во время иммунологических межклеточных взаимодействий. Его продукты контролируют трансплантационный иммунитет и принимают участие в детальной регуляции других иммунных реакций.
Второй этап - стимуляция и клональная экспансия тех В- и Т-эффекторных клонов, которые отвечают данному антигену. Одним из главных событий вслед за распознаванием антигена Т-лимфоцитом является так называемая клональная экспансия (соответствующий клон Т-лимфоцитов претерпевает несколько циклов деления клетки). Экспансия предусматривает клеточную пролиферацию, и перемещение клеток из одного скопления лимфаденоидной ткани в другое, поэтому механизмы иммунитета распространяются по всей системе в целом. Т-клеточная пролиферация происходит главным образом под управлением интерлейкина 2 - цитокина, секретируемого теми же клетками.
Участие Т-клеток в иммунном ответе многообразно. Различные Т-клеточные функции зависят от разных субпопуляций Т-лимфоцитов, имеющих уникальные поверхностные маркеры (CD-антигены) и отличающихся локализацией и путями миграции, благодаря наличию рецепторов к различным органо- и тканеспецифическим адгезивным молекулам адрессинам. Особый интерес представляют TCR-1 лимфоциты, которых в крови немного (до 10% Т-клеток). Дело в том, что именно данные клетки составляют подавляющее большинство внутриэпителиальных Т-лимфоцитов в системе MALT во всех лимфоэпителиальных органах, включая миндалины. Их важными свойствами являются:
внутриэпителиальное проживание;
рециркуляция между MALT-системой, лимфоузлами и кровью;
специфический репертуар Т-клеточных рецепторов, направленных против антигенов бактерий и простейших, попадающих на слизистые оболочки или проживающие там;
способность после активации вырабатывать высокоэффективный цитотоксический лимфокин-гранулозин, представляющий собой важнейший эффектор антибактериального иммунитета слизистых оболочек;
высокая продукция интерферона у и ИЛ-5, способность осуществлять иммунный ответ в отношении собственных мутировавших или зараженных внутриклеточными паразитами клеток.
Цитотоксические Т-лимфоциты (Тцит) при контакте с антигенном распознают уникальные антигены на поверхности любых клеток и уничтожают такие клетки-мишени при помощи перфоринового <лизиса> или гранулозинового механизма, особенно актуального для TCR-1 yb клеток системы MALT. Они несут CDS-маркеры и поражают свои мишени напрямую, а также обеспечивают развитие иммунного ответа за счет секретируемых ими медиаторов иммунного ответа.
Т-хелперы контролируют функции Т и В клеток путем различных Т-Т и Т-В взаимодействий.
В ходе иммунного ответа неактивированные Т-хелперы участвуют в первичном распознавании антигена и, в зависимости от вида антигена, его дозы и пути введения, а также индивидуальных особенностей иммунной системы, при ответе на антиген превращаются в Т-хелперы 1-го или 2-го типа (Txl и Тх2). Между собой Txl и Тх2 различаются по набору секретируемых медиаторов иммунного ответа (цитокинов).
Их активация и взаимодействие с CD-8 положительными Тцит происходят в ответ на персистирующие внутри клеток микроорганизмы. Тх2 стимулируют В-лимфоциты к превращению в плазматические клетки и секреции антител и способствуют тем самым реакциям гуморального иммунного ответа, связанного с выработкой антител, т.е. гиперчувствительности немедленного типа. Эти формы иммунного ответа универсальны, но служат преобладающими, если антиген не персистирует в организме, например, при защите от микробов, не выживающих внутри фагоцитов.
В лимфоидной ткани слизистых оболочек, помимо Т- и В-клеток обнаруживаются и плазматические клетки, которые ориентированы на синтез IgA.. Данный иммуноглобулин, проходя через эпителиоциты эпителиального пласта или клетки желез слизистой оболочки, подвергается эндоцитозу в клетки эпителия слизистой оболочки через специальный полиглобулиновый рецептор и, соединяясь с секреторным компонентом эпителиального происхождения, переходит в секреторную форму (slgA), которая защищена от протеолиза при трансцитозе и устойчива к агрессивным условиям среды на поверхности эпителиального покрова и в слизи:
IgA является:
главным секреторным продуктом системы лимфоидной ткани верхних и нижних дыхательных путей;
урогенитального и пищеварительного трактов;
фактором иммунной защиты кожных секретов, молозива и молока.
Антитела класса IgA особенно эффективны в обеспечении местной антимикробной защиты в различных участках слизистых оболочек, включая миндалины.
Третий этап - поддержание и регуляция иммунного ответа. Регуляция осуществляется Т-лимфоцитами - Т-хелперами-1 и Т-хелперами-2, в которые в процессе иммунного ответа в периферических лимфоидных органах превращаются выходящие из тимуса Т-хелперы-
Особой функцией лимфоцитов является предотвращение и подавление иммунного ответа - супрессия. Как прямая антиген-специализированная супрессия, так и опосредованная антиген-специфическая супрессия играют значительную роль в различных фазах иммунорегуляции, периферической толерантности и сдерживании аутоиммунитета.
Специальных клеток, функциями которых было бы только подавление иммунного ответа, не существует. Из-за продукции медиаторов с противоположным характером действия Txl подавляют функции Тх2 и сдерживают развитие гуморального иммунитета, а Тх2 подавляют проявление функций Тх 1 и поддержание реакций хронического воспаления, т.е. Txl являются супрессорами для Тх2 и наоборот. Однако одновременное включение функций Txl и Тх2 тормозит развитие любой формы иммунного ответа и приводит к внешним признакам иммунологической недостаточности.
Таким образом, супрессия как явление есть, хотя супрессоров, как отдельной субпопуляции, не принадлежащей ни к киллерам, ни к хелперам, ни к В-клеткам - нет. Периферические Т-клетки являются долгоживущими и, будучи активированы определенными сигналами, выполняют следующие функции: пролиферацию, дифференцировку, продукцию лимфокинов. Т-клетки (как и В-клетки) могут быть активированы специфически - антиген-специфической (моноклональной) активацией и неспецифически - индуцированием поликлональными активаторами (митогенами). В первом случае происходит бласт-трансформация лимфоцитов с последующими серийными делениями, во втором случае бласт-трансформацин не происходит - митогены вызывают однократное митотическое деление.
Важным результатом любого иммунного ответа появляется формирование клеток памяти, которые (как и Txl и Тх2) формируются в результате иммунного ответа в периферических лимфоидных органах.
Как нет чистых супрессоров, так и нет клеток, которые только <помнят и больше ничего не делают>.
Клетки памяти - это все те долгоживущие лимфоциты, которые появились после первичного иммунного ответа. При контакте с антигеном они способны к усиленному, по сравнению с родоначальником, ответу на антиген. Это и Т-, и В-клетки. В-лимфоциты при вторичном иммунном ответе начинают антителообразование не с иммуноглобулинов М, как их предки, а сразу с иммуноглобулинов G. При последующих ответах эффекторные клетки рекрутируются из числа клеток памяти путем клональной экспансии.
Таким образом, клетки памяти защищают организм в случае повторного контакта с тем же антигеном от повторного инфицирования. В отличие от других иммунокомпентентных клеток срок их жизни составляет многие месяцы и годы.
Итак, функционирование лимфэпителиального глоточного кольца предполагает постоянную текущую дезинфекцию его слизистых оболочек с участием полиморфонуклеарных фагоцитов-источников животных антибиотиков и кислородных радикалов. Макрофагальные фагоциты и иные АПК обрабатывают антигены, появляющиеся в органах кольца и запускают иммунный ответ с участием лимфоцитов MALT.
Список литературы
- Новиков Д. К. Медицинская иммунология Минск, Высшая школа, 2005 г 301 с.
- Сапин М. Р. Билич Г. Л. Анатомия человека М., ОНИКС; Альянс-В. 2000 г 463 с.
- Цвeткoв Э. А. Нaумeнкo Н. Н. Лимфоэпителиальное глоточное кольцо в иммунной системе организма. Электронный ресурс - Эффективная медицина. - ссылка скрыта