
Н. Ф. Экология (теории, законы, правила принципы и гипотезы) —
| Вид материала | Закон |
Содержание3.7. Биогеографические закономерности 3.7.2. Изменение особей (популяций) в пределах видового ареала |
- Н. Ф. Экология (теории, законы, правила принципы и гипотезы), 7082.09kb.
- Ы и темы рабочей программы для сам изучения. Перечень вопросов для сам изучения, 128.89kb.
- Тематика лекций (Наименование тем, содержание, объём в часах) № п/п, 74.97kb.
- Законы сохранения и принципы симметрии, 283.17kb.
- Проверка статистических гипотез, 59.38kb.
- Аннотация дисциплины «Экология человека» Общая трудоемкость изучения дисциплины составляет, 16.4kb.
- Программа вступительного экзамена в аспирантуру по специальной дисциплине 03. 02., 89.09kb.
- Авторское право. Терминология, 213.44kb.
- Требования к экзамену по философии, 41.96kb.
- Исследование о влиянии эволюционной теории на учение о политическом развитии народов, 10156.41kb.
Биогеографические, или эколого-географические, закономерности довольно сложно классифицировать. Как только что упомянуто, любые явления происходят в пространстве, а его протяженность не может быть математически определена в формальных единицах «от сих до сих», поскольку в экологии речь идет о системной величине. Для микроорганизма кончик иглы уже огромный полигон. К тому же в обширной биогеографической литературе широко освещены весьма значительные эмпирические данные. Существуют многочисленные сводки по географии растений, животных, общей биогеографии, географии сообществ, биогеографии островов и так далее. Приводимые факты как-то обобщены, сформулировано множество частных закономерностей. Из этой массы ценных данных здесь сделана попытка выделить наиболее общие принципы, объединяемые в три основные группы: ареал и распространение видов в его пределах, изменения особей (популяций) в пределах ареала и закономерности распространения сообществ. Многие смежные вопросы, обычно также относимые к биогеографическим, рассмотрены в следующем разделе 3.8.
3.7.1. Ареал и распространение видов в его пределах
|
Существуют два основных способа изображения ареалов на картах: путем вычерчивания их границ в виде сплошной или пунктирной (в случае сомнений в верности проведения) линии и методом нанесения точек в географические места обнаружения особей вида. В первом случае обычно производится довольно смелая экстраполяция, как правило, путем соединения крайних точек обнаружения вида, т. е. оба способа отображения границ ареала фактически совпадают.
Если в какой-то географической точке никто никогда не вел фаунистические или флористические исследования, точек находок, естественно, не возникает. Кроме того, при многолетнем изучении широких регионов, как это приходилось делать автору в Средней Сибири, нетрудно убедиться, что некоторые птицы, например, коростель, порой фактически исчезают на краю своего ареала на ряд лет, а затем вновь появляются. Для упомянутого вида территориальная изменчивость границ ареала составляет сотни километров, так что выразима в самом мелком масштабе. Для менее подвижных животных и растений такие колебания границ ареала невозможны, но в годы депрессий численности экологическая роль видов, находящихся в минимуме, практически равна нулю или стремится к нему. Исследователь может вид не обнаружить по чисто техническим причинам.
Возможности сопряжения и прогнозирования границ ареалов связаны с конгруэнтностью видов в составе биоценозов и сообществ (разд. 3.8.). Как показывает опыт, имеется соответствие видов и ценозов (см. ниже), и эти физиономически ясные черты позволяют отображать такие закономерности на картах. Однако довольно трудно (хотя и возможно) найти два вида, которые бы имели абсолютно сходное распределение в пространстве. Эту особенность Л. Г. Раменский (1924) и Г. А. Глизон (1926) сформулировали в виде индивидуалистической гипотезы, правила или принципа экологической индивидуальности видов: каждый вид пространственно распределен в соответствии со своими генетическими, физиологическими и другими биологическими особенностями и глубоко специфично относится к факторам среды, в том числе к другим видам. Как нет абсолютно идентичных особей (кроме однояйцевых близнецов, тем не менее нередко фенотипически отличающихся друг от друга), так нет и биоэкологически идентичных видов. Следовательно, их распространение сугубо индивидуально, а потому подвержено столь же индивидуальным нормам реакции на возникающие изменения среды жизни. Как сказано выше, этот принцип не абсолютен.
В виде регистрации факта можно сформулировать правило колебания границ ареала: многие виды не имеют четко отграниченной постоянной области распространения и временами либо исчезают на границе ареала, либо достигают исчезающе малой численности. В. Тишлер в 1955 г. сформулировал принцип воздействия факторов, согласно которому границы, размер и характер ареала вида или местообитания популяции обусловлены их биологическими особенностями, и наоборот, биологические особенности вида или популяции могут указывать (быть индикаторами) места, где их можно найти*.
| * Очень примитивно говоря, рыбы живут в воде — одна в реке, другая в озере, третья в море; одна на глубине, другая на поверхности и т. д. |
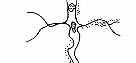
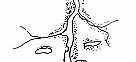
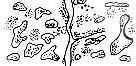
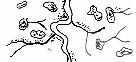
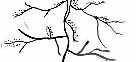
Рис. 3.5. Структура поселений и особенности стационарного размещения водяной крысы в различных ландшафтных зонах Западной Сибири (рис. из кн.: Максимов А. А. Водяная крыса и борьба с ней в Западной Сибири. Новосибирск: 1959. С. 48) | ||||||||||||||||||||||||

Как показывают широкомасштабные исследования в Сибири и на Дальнем Востоке, виды очень четко соответствуют ценозам, а если наблюдается разница в постоянно регистрируемом видовом составе всего в один вид, то ценоз явно резко отличается от географически или топографически соседних.
Как правило, смена видового состава происходит не на один, а на группу видов, и действительно, вид служит индикатором того сообщества, куда он входит.
Там, где природа сильно нарушена человеческой деятельностью, описанное выше правило соответствия вида и ценоза заметно не столь четко. Лоскутные природные образования складываются не в рамках бывших границ элементарных биогеоценозов, а составляют объединенные экосистемы топографически (но не экологически) более высокого иерархического уровня. Вид как бы вписывается не в «свой» изначальный ценоз, а в мозаику биогеоценотических парцелл разных экосистем, его экологическая ниша оказывается соотнесенной с этой мозаикой. Отсюда следует вывод, что правило соответствия вида и ценоза в этом случае распространяется на более географически широкие образования, а колебания границ ареалов могут в этом случае быть менее выраженными и не столь заметными.
Необходимо обратить внимание на то, что в ходе упомянутых процессов нарушается принцип территориальной общности физико-географических единиц (разд. 3.7.3), они укрупняются из-за мозаичности возникающих «обрывков» экосистем и их возрастающей отдаленности друг от друга. Иногда «промежуточные» экосистемы (по иерархии соподчинения) вообще исчезают, как, впрочем, и элементарные, в своем первозданном виде. Иерархия экосистем резко упрощается по числу уровней — сохраняется как целое лишь 2-й снизу уровень биома или ландшафтной полосы (см. главу 2) и только им соответствуют по распространению виды, занимая микроучастки, едва ли составляющие полноценные экосистемы (скорее, их парацеллы).
Там, где сохранилась первичная структура природных образований, соответствие вида и ценоза дает очень четкую пространственную картину кружева ареала, закономерно меняющегося при движении в географическом пространстве. Эта закономерность в разд. 3.6 была названа правилом топографического, или популяционного, кружева ареала. Это правило было сформулировано для топографического пространства одной географической точки. Теперь следует обратить внимание на изменения этого кружева в более широком пространстве — правило географической изменчивости кружева ареала. Обычно оно состоит в том, что на севере вид приурочен к приречным, низменным участкам, затем к югу заселяет более широкие пространства, в горах обитает лишь в локальных участках, а на южном пределе распространения вновь приурочен к приречным более влажным и прохладным местообитаниям. У каждого широко распространенного вида характер изменения кружева ареала специфичен, но смены происходят закономерно. Такие смены, пожалуй, лучше всего известны для водяной крысы в Сибири* (рис. 3.5). Пользуясь выявленными закономерностями, можно получить более надежные критерии для экстраполяции границ ареалов видов.
| * Очень показательная схема была опубликована (с. 48) в сборнике «Водяная крыса и борьба с ней в Западной Сибири». Новосибирское кн. изд-во, 1959 (Максимов А. А. Распределение водяной крысы по зонам и ландшафтам Западной Сибири. С. 19—48). См. также Пантелеев П А Популяционная экология водяной полевки и меры борьбы. М.: Наука, 1968. 255 с. ** РеймерсН. Ф. Птицы и млекопитающие южной тайги Средней Сибири. М.-Л.: Наука, 1966. 420 с.; Атлас Сахалинской области. М.: ГУГК, 1967. |
Правило соответствия вида и ценоза позволило автору предложить новый тип зоогеографических и шире — биогеографических — карт**, основанных на экосистемном подходе. При этом правило колебаний границ ареала в мелком масштабе удалось отразить лишь частично. Зато в крупном масштабе оно вполне может быть реализовано — ценозы, где наблюдается пульсация ареалов, графически на картах отличаются от тех, где вид постоянно присутствует.
Наиболее общим объяснением причин формирования границ ареала вида служит правило ограничивающих факторов: факторы среды, наиболее удаляющиеся от оптимума экологических потребностей вида, лимитируют возможности его существования в данных условиях. Поскольку к лимитирующим факторам относятся любые условия существования вида — как абиотические, так и биотические, включая антропогенные,— правило ограничивающих факторов, ведущее свое начало от группы законов минимума (см. разд. 3.5.2), включая закон толерантности Шелфорда (см. разд. 3.5.1), практически дополнительно ничего не объясняет, а лишь резюмирует перечисленные закономерности.
Для объяснения механизма действия принципа воздействия факторов В. Тишлера (см. выше) было предложено несколько теоретических оснований. Одно из них — правило стимулирующего действия температур В. Шелфорда — Т. Парка, в умеренных широтах определяющее стимулирующее воздействие, которое оказывают изменения температуры среды обитания (правило сформулировано независимо друг от друга В. Э. Шел-фордом в 1929 г. и Т. Парком в 1930 г.). Это правило в 1972 г. А. Н. Голиков и О. А. Скарлато дополнили правилом лимитирующего значения крайних летних и зимних температур, или «биогеографическим методом» указанных авторов: в умеренных широтах Северного полушария распространение видов ограничено необходимыми для размножения температурами. Эта потребность является генотипически закрепленным видовым признаком и прямо не связана с диапазоном температур выживаемости особей вида. У видов, переносящих широкие колебания температур, температура размножения часто более контрастна, чем у видов с относительно низким порогом выносливости к переменам количества тепла.
Очевидно, нельзя считать, что температура — абсолютно определяющий фактор в формировании границ ареала. Существенную роль играют влажность, субстраты, межвидовые отношения, преследование человеком, а в водной среде также скорость движения ее потоков и соленость вод. Саму температуру регулируют складывающийся мезо-, микро- и биоклимат. Играют роль и генетические закономерности взаимоотношения видов.
Среди них выделяется правило викариата Д. Джордана: ареалы близкородственных форм организмов (правило установлено для животных) обычно занимают смежные территории и существенно не перекрываются; родственные формы, как правило, викарируют, т. е. географически замещают друг друга. Согласно правилу представительства рода одним видом А. Монара (1919 г.), в однородных условиях и на ограниченной территории систематический (таксономический) род, как правило, представлен только одним видом. Видимо, это связано с близостью экологических ниш видов одного рода.
В связи с этим необходимо вспомнить группу закономерностей, связанных с принципом конкурентного исключения, или законом (теоремой) Г. Ф. Гаузе (1934 г.): два вида с близкими экологическими требованиями длительное время не могут занимать одну экологическую нишу и, как правило, входить в одну экосистему. Следовательно, одновременно происходит и географическое замещение видов, что констатирует принцип видо-родового представительства Й. Иллиеса: поскольку два близкородственных вида не могут занимать одинаковые экологические ниши в одном биотопе, и, соответственно, в биоценозе, богатые видами роды обычно представлены в экосистеме единственным своим представителем. Математически подобные отношения были доказаны уравнениями А. Лотки — В. Вольтерры (независимо этими авторами в 1925 и 1926 гг.). Однако поскольку принцип конкурентного исключения, видимо, не абсолютно универсален, был сформулирован принцип сосуществования, или парадокс Дж. Хатчинсона: два вида в порядке исключения могут сосуществовать в одной экологической нише, и если близкие виды вынуждены использовать одни и те же ресурсы, то естественный отбор может благоприятствовать их сосуществованию, и они способны эволюционировать в близком направлении. Парадокс Дж. Хатчинсона наблюдается главным образом в водных экосистемах*.
| * Кроме того, особи морфологически сходных видов, обитающих совместно, статистически различаются по размерам. |
Ясно, что перечисленные биогеографические закономерности неравнозначны по глубине обобщения и вкладываемому смыслу.
Совершенно очевидно, что все закономерности системы «организм — среда» (разд. 3.5) отражаются на формировании границ ареалов видов. Наибольшее индикаторное значение имеет правило соответствия вида и ценоза, которое хотя и не объясняет причин такого соответствия, но отражает реально существующий факт, возникающий на почве сложного переплетения многих закономерностей генетического, аутоэкологического и синэкологического характера. Они формируют кружево видового ареала. Его географические пределы зависят от толерантности особей вида к физическим условиям среды и способности выработать экологическую нишу в одном или в нескольких ценозах, само существование которых определяется свойствами не столько отдельных видов, сколько их сообществ (разд. 3.8). Однако и тут важны характеристики видов-средообразователей — эдификаторов среды. Лишь во взаимодействии свойств популяций и их системных объединений складывается картина ареала. Поэтому все эти закономерности важны для анализа его границ и особенностей.
Нельзя сказать, чтобы внутренние механизмы формирования ареалов были до конца ясны, а главное, складывались в совокупность знаний, вполне пригодную для практической деятельности. Лучше обстоит дело с описанием внешних свойств географически сменяющих друг друга популяций, особенно животных. Эти закономерности сведены в следующем разделе.
3.7.2. Изменение особей (популяций) в пределах видового ареала
|
Группа закономерностей, характерных для пространственных изменений особей в популяциях, сменяющих друг друга при движении с севера на юг (или наоборот) и живущих в различных условиях островной изоляции, наиболее известна. Кажется, еще Фридрих I Барбаросса (1125 — 1190) знал правило измельчания размеров тела теплокровных животных при движении с севера на юг, названное затем правилом К. Бергмана (1847): у теплокровных животных, подверженных географической изменчивости, размеры тела особей статистически (в среднем) больше у популяций, живущих в более холодных частях ареала вида. Правилу Бергмана подчиняются около 50% видов теплокровных животных, подавляющая часть которых (до 90%) птицы. Механизм действия правила Бергмана очевиден — укрупнение размеров особей термодинамически позитивно меняет отношение объема и площади тела, снижает уровень основного обмена. В то же самое время правило не может действовать в тех случаях, когда наблюдается недостаток кормов или колеблется их доступность: при увеличении размеров тела общая масса потребляемого корма возрастает*. Очевидна связь правила Бергмана с правилом поверхностей (разд. 3.4.2).
| * Но если вид способен к временному снижению метаболизма, а соответственно и температуры тела, то особи могут не укрупняться при распространении с юга на север. К тому же результату ведет способность использовать терморегулирующие укрытия, что характерно для млекопитающих. |
Столь же очевидны причины действия закона покровов (покрытия) тела: плотность (густота и терморегулирующая способность) покровов тела млекопитающих и птиц достигает максимума в холодных и засушливых областях. Это помогает животным противостоять экстремальным температуре и влажности. Кстати, этому закону следуют и люди, одевая наиболее плотную одежду в тех же местах (меховая одежда на севере, стеганые халаты и длинная одежда в пустынях). Уточнение к закону покровов (покрытия) тела служит правило густоты мехового покрова: у млекопитающих холодных местообитаний мех как правило гуще, чем у живущих в теплых географических зонах и поясах. Часто это правило справедливо и для млекопитающих сухого климата, где густой мех предохраняет от излишнего испарения воды с поверхности тела, а также от перегревания.
Как бы частным случаем правила Бергмана, а скорее, более общего правила поверхностей (разд. 3.4.2) служит правило Д. Аллена (1877): увеличение выступающих частей тела одного вида теплокровных животных (конечностей, хвоста, ушей и т. п.) при продвижении с севера на юг. Достаточно ясно, что удлинение выступающих частей тела увеличивает поверхность тела и его теплоотдачу.
Для рыб существует особое правило числа позвонков, или правило А. Жордана, согласно которому (у сельдей, трески и др.) в водоемах с повышенной соленостью и более низкими температурами в хвостовой части тела возрастает число позвонков. Правило Жордана не имеет отношения к правилу Аллена как таковому, а связано с приспособлением к движению в более плотной среде. Подобного рода закономерностей много у почвенных и глубоководных организмов — их удлиненность, часто червеобразность и т. п.
| * Так называемый индустриальный меланизм — потемнение некоторых животных, например бабочек, в задымленных регионах — видимо, имеет подосновой необходимость более темной защитной окраски. |
Многие признаки островных популяций видов отличаются от имеющихся у материковых. Объясняет это явление принцип основателя: единичные особи-основатели новой (изолированной) колонии или популяции несут в себе лишь часть общей генетической информации, присущей виду. Нехватка потенциала наследственной изменчивости может лежать в основе правила островного измельчания: популяции видов животных, обитающие на островах, как правило, образуются более мелкими особями, чем материковые, живущие в аналогичных условиях. Правило островного измельчания имеет много исключений, особенно на крупных островах (например, кабарга Сахалина крупнее чем материковая), но, видимо, статистически верно.
Закономерности географического распространения сообществ также привлекли внимание экологов и биогеографов. Перейдем к краткому их перечислению.