
Н. Ф. Экология (теории, законы, правила принципы и гипотезы) —
| Вид материала | Закон |
Содержание3. 3. Физико-химические и молекулярно-биологические |
- Н. Ф. Экология (теории, законы, правила принципы и гипотезы), 7082.09kb.
- Ы и темы рабочей программы для сам изучения. Перечень вопросов для сам изучения, 128.89kb.
- Тематика лекций (Наименование тем, содержание, объём в часах) № п/п, 74.97kb.
- Законы сохранения и принципы симметрии, 283.17kb.
- Проверка статистических гипотез, 59.38kb.
- Аннотация дисциплины «Экология человека» Общая трудоемкость изучения дисциплины составляет, 16.4kb.
- Программа вступительного экзамена в аспирантуру по специальной дисциплине 03. 02., 89.09kb.
- Авторское право. Терминология, 213.44kb.
- Требования к экзамену по философии, 41.96kb.
- Исследование о влиянии эволюционной теории на учение о политическом развитии народов, 10156.41kb.
|
Глава 2 была фактически целиком посвящена иерархии систем, прежде всего биосферы и входящего в нее живого. Общие принципы формирования иерархии: 1) дублирование относительно разнокачественных структур, составляющих в своей организованной совокупности нечто новое, т. е. наличие свойства эмерджентности (древние говорили: целое больше суммы его частей) и 2) определенность функциональной цели организации в рамках связей со средой и внутренних возможностей системы. Сам принцип иерархической организации, или принцип интегративных уровней, в биологии и экологии принимается как аксиома или эмпирически наблюдаемый факт (разд. 3.10). Столь же аксиоматически утверждается и проявление эмерджентности с переходом от одного уровня иерархии к другому. Эмерджентность — наличие у системного целого особых свойств, не присущих его подсистемам, элементам и (несистемным) блокам, а также сумме элементов и блоков, не объединенных системообразующими связями. Свойство цели как функциональное состояние и закономерность построения системы, достигаемая путем возникновения обратных связей, создает некое поле взаимодействий. Это поле не может быть бесконечным по способу организации, так как любая система существует в рамках ее характерного времени и пространства (размера).
Размер системы, или характерный размер системы,— это пространственная ее протяженность (объем, площадь) или масса, а также минимальное (максимальное) число подсистем, позволяющее системе существовать и функционировать с осуществлением саморегуляции и самовосстановления в рамках своего характерного времени. Системное время (характерное, или собственное, время системы) — это время, рассматриваемое в рамках периода существования данной системы и/или происходящих в ней процессов. Эти процессы ограничены термодинамикой системы, ее функциональными особенностями. Сочетание цели системы, ее характерного времени и пространства создает предпосылки для действия закона оптимальности, разобранного в разд. 3.2.1. В то же самое время, поскольку системы с одной и той же функциональной целью, формируемой обратными связями, располагаются на одном уровне иерархии и поэтому ограничены однотипным характерным временем и пространством, их построение подчинено одному ряду внутренних закономерностей. Это — смысловое «третье измерение» табл. 2.1, упомянутое в главе 2.
Ограничение числа и форм взаимодействия подсистем одного системного уровня ограничивает «фантазию природы», укладывает ее в строго лимитированный закономерный ряд образований. Периодический закон химических элементов Д. И. Менделеева (свойства химических элементов, проявляющиеся в простых веществах и соединениях, находятся в периодической зависимости от заряда ядер их атомов) был первой формализацией «третьего измерения» таблицы «системы систем» (см. табл. 2.1). Тот же характер имеет закон гомологических рядов и наследственной изменчивости Н. И. Вавилова. (Родственные виды, роды, семейства и др. систематические категории обладают закономерно возникающими гомологичными генами и порядками генов в хромосомах, сходство которых тем полнее, чем эволюционно ближе сравниваемые таксоны. Гомология генов у родственных форм проявляется в сходстве рядов их наследственной изменчивости. Циклы изменчивости проходят через все роды и виды, составляющие семейство). Менее известен и признан периодический закон географической зональности А. А. Григорьева — М. И. Будыко: со сменой физико-географических поясов Земли аналогичные ландшафтные зоны и их некоторые общие свойства периодически повторяются (рис. 3.2).
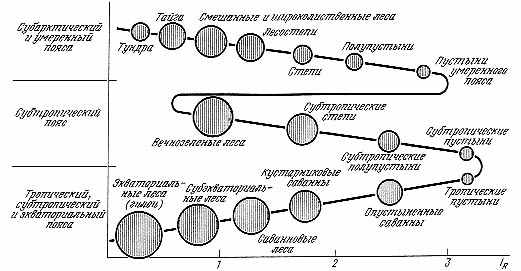 Рис. 3.2. Периодический закон географической зональности: IR — радиационный индекс сухости (отношение радиационного баланса к количеству тепла, необходимому для испарения годовой суммы осадков). Диаметры кружков пропорциональны биологической продуктивности ландшафтов |
Периодическое повторение свойств в рядах систем одного иерархического уровня, видимо, общий закон мироздания. Он был сформулирован как закон периодичности строения системных совокупностей, или системо-периодический закон. Формулировка первого варианта: конкретные природные системы одного подуровня (уровня) организации составляют периодический или повторяющийся ряд морфологически аналогичных структур в пределах верхних и нижних системных пространственно-временных границ, за которыми существование систем данного подуровня делается невозможным (они переходят в неустойчивое состояние или превращаются в иную системную структуру, в том числе другого уровня организации)*. Более обобщающая формулировка дана мною в словаре-справочнике «Природопользование»: принципы структурного построения и управления однородных природных систем в иерархическом соподчинении и особенно сложении таких же природных систем одного уровня организации (иерархии) повторяются с некоторой правильностью в зависимости от действия единого (комплекса) системообразующего фактора (факторов)**. Конкретные примеры иерархии экосистем приведены в главе 2.
| * Реймерс Н. Ф. Системные основы природопользования//Философские проблемы глобальной экологии. М.: Наука, 1983. С. 140. ** Реймерс Н. Ф. Природопользование. М.: Мысль, 1990. С. 156. |
Системопериодический закон можно считать всеобщей закономерностью, действующей во всем системном мире, в том числе в системах с участием живого — экосистемах. Поскольку они всегда составлены живым и средой, окружающей это живое, очевидно, следует обратиться к группе связей, определяющих основные принципы взаимоотношений системы с окружающей ее средой.
3.2.5. Отношения «система — среда»
|
Абсолютно изолированные системы вне связи с окружающей их средой длительное время существовать не могут в силу действия второго начала термодинамики (закона возрастания энтропии — разд. 3.2.3). Самые общие закономерности взаимосвязи система — среда обобщены в философски расширенной формулировке принципа дополнительности Нильса Бора.
Изначальный смысл этого принципа (при приборном исследовании физического микрообъекта могут быть получены точные данные либо о его поведении в пространстве и во времени, либо о его энергиях и импульсах как о двух взаимно исключающих картинах, «дополняющих» друг друга) был расширен до: две взаимосвязанные, но различные материальные системы дополняют друг друга в своем единстве и противоположности. Такой «общий принцип дополнительности» очень существенен для функционирования экологических систем (см. в разд. 9.1 принципы экологической конгруэнтности и экологической комплементарности, а также закон единства организм — среда в разд. 3.5.1).
Без относительного (динамического) равновесия в рамках «общего принципа дополнительности» взаимодействие будет кратковременным: система разрушится. Этого не происходит в силу действия принципа Ле Шателье — Брауна, формулировку которого мы повторим (разд. 3.2.3.): при внешнем воздействии, выводящем систему из состояния устойчивого равновесия, равновесие смещается в том направлении, при котором эффект внешнего воздействия ослабляется. Следствием этого служит принцип торможения развития: в период наибольших потенциальных темпов развития системы возникают максимальные тормозящие эффекты. В зависимости от силы процесса они могут быть заметны или скрыты ходом этого процесса.
Вещество и энергия для функционирования и развития системы могут поступать лишь из окружающей эту систему среды, и только за счет этой среды может существовать и прогрессировать любая система. Этот очевидный факт отражает закон развития системы за счет окружающей ее среды: любая система может развиваться только за счет использования материально-энергетических и информационных возможностей окружающей ее среды; абсолютно изолированное саморазвитие невозможно. Этот закон действует как в сфере природных, так и социальных (видимо, принципиально всех) систем природы и общества (кроме, быть может, механических).
Энергия, вещество и информация, поступающие в систему извне и выступающие как факторы ее жизни, действуют не в «чистом» виде, а селектируются и видоизменяются этой системой. Если они проходят предварительно через надсистемы рассматриваемой системы, то эти процессы идут многократно и до нее доходят в трансформированном всеми надсистемами виде. Поиск прямых связей между очень далекими по иерархическому уровню системными образованиями (например, между активностью Солнца и массовыми размножениями организмов), если эти связи не настолько сильны, что проходят «транзитом» через промежуточный ряд иерархии систем, как правило, бывает очень затруднителен. Действует принцип преломления действующего фактора в иерархии систем. При этом не следует сбрасывать со счета и саму рассматриваемую систему как преобразующий фактор члена иерархии: фактор, действующий на систему, преломляется через всю иерархию ее надсистем и через функциональные особенности самой системы. В связи с этим, как правило, воздействия надсистем не равны по силе и не совпадают по времени с интенсивностью и моментом их возникновения.
Вероятно, лучше рассматривать отдельно принцип преломления действующего фактора внутри системы (внешние для систем воздействия, как правило, проявляются не прямо, а опосредованы механизмами функционирования этой системы; они могут быть ослаблены ее буферными свойствами или усилены возникающими цепными реакциями), справедливый для не моментально гибельных для системы воздействий, и только что сформулированный принцип преломления действующего фактора в иерархии систем. В первом случае буфером оказываются механизмы самой системы, а во втором — ее надсистем. В данной формулировке в иерархию систем включена и сама система, на которую воздействует тот или иной фактор, что равно можно принимать или оспаривать в зависимости от подхода.
Упрощенное, механистическое представление о действующих факторах очень распространено и приводит к многочисленным ошибкам.
В силу преломления действующего фактора в иерархии систем и наличия многих «фильтров» этот фактор либо ослабляется, либо усиливается, а чаще всего оказывается неравномерным по силе воздействия с ходом времени. Система немедленно или с задержкой реагирует на возникающие флуктуации. Этот факт констатируется законом функционально-системной неравномерности: темпы реакций и прохождения фаз развития системы (в ответ на действие внешних факторов) закономерно неравномерны — они то убыстряются (усиливаются), то замедляются (ослабевают) .
Ритмика таких колебаний обычно кратна трем. Об этом говорят эмпирические факты, прямые наблюдения в природе. Была выдвинута (советскими учеными Г. Розенбергом и С. Рудерманом) гипотеза и сформулирован принцип скользящих среднемаксимальных случайного статистического ряда: периоды между максимумами временного (или любого другого случайного условно бесконечного статистического) ряда величин имеют определенную цикличность вне зависимости от характера и происхождения самих наблюдаемых величин, т. е. математическое ожидание величины расстояния от максимума до максимума средних, равномерно расположенных значений признака, не зависит от характера самого случайного ряда. Чаще всего наблюдаются периоды, близкие к n · 3, 3n, 4 и 5. Таковы циклы солнечной активности, колебания численности организмов. Весьма вероятно, что в основе упорядоченности циклов колебаний на Земле лежат космические процессы.
Если речь идет не об условно бесконечном ряде статистических величин, а о конечном явлении, то обычно в развитии систем действует правило затухания процессов: насыщающиеся системы с увеличением степени равновесности с окружающей их средой или внутреннего гомеостаза (в случае изолированности системы) характеризуются затуханием в них динамических процессов. Это затухание может быть постепенным при линейности процесса или идти скачкообразно по затухающей синусоиде, лавинообразно (экспоненциально и сверхэкспоненциально). Такое явление характерно для насыщающихся растворов, термодинамических процессов, темпов размножения акклиматизированных организмов, экономического развития стран и регионов и других природных и социальных явлений.
Частным, но важным случаем правила затухания процессов и одновременно следствием закона оптимальности (разд. 3.2.1), перекликающегося в данном случае со вторым принципом термодинамики (разд. 3.2.3), служит закон растворения системы в чуждой среде, выведенный советским геофизиком Г. Ф. Хильми. Он действует в любых системах, в том числе социальных (при большой разнице «культурного острова» и среды, в которой он находится, этот остров бывает уничтожен, что и было в результате репрессий после 1917 г. в нашей стране). Особенностью социальных «островов» является то, что в них малые системные образования, растворяясь, увлекают за собой окружающую их среду, что в совокупности приводит к явлению пассионарности, детально исследованному Л. Н. Гумилевым в ряде работ, в том числе в нашумевшей книге «Этногенез и биосфера Земли» (Л.: Изд-во ЛГУ, 1989. 495 с.). Формулировка закона растворения системы в чуждой среде приведена в особой трактовке в разд. 3.7.3 при обсуждении биогеографических закономерностей.
Общесистемные закономерности, изложенные выше, распространяются на экологические системы и действуют либо сами по себе, либо в специфическом для природных систем с участием живого виде. Нередко они сформулированы как узкоэкологические законы, правила, принципы и так далее. В дальнейшем изложении необходимо будет обратить внимание на это.
Перейдем к общим физико-химическим и молекулярным закономерностям возникновения, построения и фукнционирования живого.
3. 3. ФИЗИКО-ХИМИЧЕСКИЕ И МОЛЕКУЛЯРНО-БИОЛОГИЧЕСКИЕ
ОСНОВЫ СУЩЕСТВОВАНИЯ ЖИВОГО
| |
| | |
| * В отечественной литературе встречается иная транскрипция термина — киральный. ** Эти вопросы подробно изложены в ряде статей рано скончавшегося Л. Л. Морозова и его соавторов. Можно рекомендовать две научно-популярные работы: Л. Л. Морозов. Несохраняющаяся четность в молекулярном мире организмов//Природа. 1977. № 1. С. 32 — 45. Он же (посмертная статья). Поможет ли физика понять, как возникла жизнь?//Природа. 1984. № 12. С. 35 — 48. *** Это легко доказать математически. Возникновение существующих цепей из 2000 ферментов, характерных для живого, требует перебора 1040000 вариантов их сочетаний (Викрамасингхе Ч. Размышления астронома о биологии//Курьер ЮНЕСКО. 1982. Июнь. С. 36— 38). Возникновение генома человека, сложность которого оценивается числом 101000, требовало бы в эволюции живого фантастического перебора вариантов, равного более 10900 на одну особь (Реймерс Н. Ф. История эволюции//Человек и природа. 1982. № 11. С. 50 — 61). Числа эти абсолютно нереальны — см. закон направленности эволюции в следующих разделах главы, а также разд. 3.11. |
Хиральная чистота обуславливает специфику живого, несводимость его к неживому и практическую невозможность получения живого из неживого в современных условиях Земли. Для этой планеты возникновения жизни — уникальный, катастрофический процесс**. Закон хиральной чистоты и тот факт, что синтезировать хирально чистые вещества в лабораторных условиях возможно лишь с помощью весьма сложных методик асимметрического синтеза, делают бессмысленными спекулятивные рассуждения об отсутствии граней между неживым и живым. Искусственное конструирование живого, если говорить о природе Земли, практически неосуществимо***. Вместе с тем, хиральная асимметричность, свойственная всему живому, объективно указывает на его физико-химическое единство, что и отражает закон физико-химического единства живого вещества В. И. Вернадского: все живое вещество Земли физико-химически едино.
Этот закон не исключает количественной биогеохимической специфики видов живого, даже половых и возрастных, возможно индивидуальных химических и физических отличий у тканей организмов. Однако качественно жизнь едина, и потому подчиняется единым «правилам игры», в том числе термодинамическому правилу Вант-Гоффа — Аррениуса, правилу поверхностей и биогеохимическим принципам В. И. Вернадского.
Термодинамическое правило Вант-Гоффа — Аррениуса в биолого-экологической модификации гласит, что подъем температуры на 10°С приводит к 2 — 3-кратному ускорению химических процессов. Фактически обмен веществ в одних случаях усиливается многократно (до 7,4 раз), а в других повышение температуры его замедляет.
Правило Вант-Гоффа — Аррениуса существенно углубляется закономерностью, формулируемой для теплокровных животных в виде правила поверхностей. Формулировку и расшифровку этого правила технически и по смыслу удобней оказалось привести в разд. 3.4.2.
Экологический эффект всемирного повышения температуры на 10°С довольно трудно предсказуем. Скорее всего изменение среднеглобальной температуры в таком размере катастрофично. Среднеглобальная температура атмосферы у поверхности Земли около 15°С. За последний 1 миллион лет она изменялась в пределах 5°С похолодания и 2°С потепления. При изменении среднеглобальной температуры на 10°, т. е. в 1,5 раза от современного уровня, скорее всего будет нацело нарушено действие принципа Ле Шателье — Брауна (разд. 3.2.3) — биота как бы сама себя «съест», так как процессы обмена веществ, усиливаясь, приведут не к сопротивлению изменениям в окружающей биоту среде, а к быстрой самодеструкции биосферы.
При этом будут глубоко нарушены первые два биогеохимических принципа В. И. Вернадского, а третий его принцип будет существенно искажен. Первый биогеохимический принцип В. И. Вернадского гласит: биогенная миграция атомов химических элементов в биосфере всегда стремится к максимальному своему проявлению. Этот принцип уже в наши дни нарушен, хотя и обратимо, как сказано выше при обсуждении принципа Ле Шателье — Брауна (разд. 3.2.3). Второй биогеохимический принцип В. И. Вернадского: эволюция видов в ходе геологического времени, приводящая к созданию устойчивых в биосфере форм жизни, идет в направлении, усиливающем биогенную миграцию атомов. Этот принцип при антропогенном измельчании средних размеров особей биоты Земли в ходе процессов экологического дублирования (лес сменяется лугом, крупные животные мелкими) начинает действовать аномально интенсивно (см. правило Ю. Одума в разд. 3.8.1), что также нарушает действие принципа Ле Шателье — Брауна.
Поскольку, согласно третьему биогеохимическому принципу В. И. Вернадского, живое вещество находится в непрерывном химическом обмене с космической средой, его окружающей, и создается и поддерживается на нашей планете космической энергией Солнца, биосферная солнечно-земная связь с нарушением первых двух биогеохимических принципов В. И. Вернадского и принципа Ле Шателье — Брауна резко изменяется. Космические воздействия могут из системы поддержания биосферы планеты превратиться в агенты, ее разрушающие. Процесс может стать саморазвивающимся и необратимым. Пока биосфера еще находится в обратимом состоянии, но угроза ее самодеструкции все время растет. К этому вопросу целесообразно вернуться после рассмотрения всей группы закономерностей, регулирующих функционирование природных систем планеты. Начнем этот перечень с экологических законов, правил и принципов жизни отдельного организма.
| * Летунов В. Н. Термодинамические аспекты теории адаптаций//Тр./Зоол. ин-т АН СССР. 1987. Т. 160. С. 31 — 40. |
Чтобы закончить с проблемами термодинамики и энтропии и перейти к биосистемам, обратимся к группе теорем, или афризмов, сформулированных для уровня отдельной особи. Первой в этом ряду стоит теорема (афоризм) Э. Шредингера (1944) о «питании» организма отрицательной энтропией. Эта теорема утверждает, что упорядоченность организма (особи) выше, чем окружающей его среды и он (организм) отдает в эту среду больше неупорядоченности, чем получает:
deS — diS = —S, если diS ≥ deS,
где diS — внешняя энтропия, a deS — внутренняя энтропия.— S выступает как мера упорядоченности. Совершенно очевидна связь теоремы Э. Шредингера с теоремой сохранения упорядоченности И. П. Пригожина (разд. 3.2.3).
Следующие три обобщения носят столь же уточняющий характер. Их ряд начинается теоремой (афоризмом) Хаасе (1963): организм «питается» положительной энтропией, т. е. энергетическая ценность пищи выше, чем этот же показатель продуктов диссимиляции. Иначе — организм существует до тех пор и постольку, поскольку имеется положительный энергетический баланс.
Теорему (афоризм) Э. Шредингера дополняет теорема Г. Атлана (1968), утверждающая, что организм «питается» не только отрицательной энтропией, но и «шумом». Часть информации, содержащейся в пище, в ходе ассимиляции неизбежно теряется.
Завершает группу сходных обобщений теорема Бриллуэна (1956) о том, что энтропия отходов больше энтропии пищи. По сути дела, по содержанию это повторение теоремы Э. Шредингера:
Sns — Sab == — S, если Sab ≥ Sns,
где Sns — энтропия пищи, a Sab — энтропия продуктов диссимиляции. Перечисленные теоремы «оголяют» физический смысл достаточно очевидных истин.