
Литература
| Вид материала | Литература |
СодержаниеPinus silvestris L.) Экспериментальная часть Содержание марганца 3. Результаты и их обсуждение |
- Класс: 12 Зачёт №2 «Русская литература 1917-1941», 186.43kb.
- Литература 7 класс Зачетная работа№2 Содержание, 40.79kb.
- Литература Форш О. «Одеты камнем», 38.6kb.
- Литература английского декаданса: истоки, становление, саморефлексия, 636.47kb.
- Тема: «Библейские мотивы в творчестве Б. Пастернака», 211.88kb.
- Литература 11 класс Программы общеобразовательных учреждений. Литература, 331.28kb.
- Учебника. Учитель Кулябина Зинаида Григорьевна. Выступление на Краевой научно-практической, 48.58kb.
- Жиркова Р. Р. Жондорова Г. Е. Мартыненко Н. Г. Образовательный модуль Языки и культура, 815.79kb.
- Английская литература. , 57.37kb.
- Планирование курса литературы 8 класса по программе, 14.4kb.
45-я городская заочная конференция
Челябинского научного общества учащихся
«Интеллектуалы ХХΙ века»
Сезонная динамика содержания фотосинтетических пигментов и марганца в хвое сосны обыкновенной (Pinus silvestris L.)
Автор:
Перевышина Кристина
11 класс, МОУ
«гимназия» № 102,
Научный
руководитель:
Похлебаев, Сергей
Михайлович
д.п.н., профессор
ЧГПУ
Челябинск
2010
| Содержание |
| Введение……………………………………………………………………..
1.1. Механизмы защиты и устойчивости у растений…………….. 1.2. Влияние температуры на пигментный аппарат хвои……….. 1.3. Физиологическое значение марганца………………………..
фотосинтеза в хвое сосны обыкновенной в зависимости от сезонности………………………………………………………
обыкновенной……………………………………………………… Выводы..................................................................................................... Литература………………………………………………………………. Приложение………………………………………………………………. |
Введение
Эколого-физиологические исследования, раскрывающие механизмы взаимодействия растений со средой, имеют большое значение для понимания их географического распределения и анализа соответствия метаболизма климатическим условия [1]. Среди механизмов адаптации особое место занимает так называемый "ответ на температурный стресс". Ответ растений на действие низкой температуры сопровождается многочисленными физиологическими и биохимическими изменениями, затрагивающие многие стороны метаболизма растительной клетки [2].
Удобной моделью для изучения ответных реакций на действие низких температур являются хвойные растения. Способность большинства из хвойных сохранять листья круглый год и осуществлять фотосинтез даже зимой свидетельствует о широком температурном спектре действия ферментативных систем этих растений и специфичности состояния многих кофакторов фотосинтетического процесса. В связи с этим исследование структуры и функции их фотосинтетического аппарата представляет несомненный теоретический интерес с позиций понимания общего и фотосинтетического метаболизма листа хвойных, в частности, и растений зимующими листьями в целом [3].
Хорошо известно, что содержание пигментов и активность работы фотосинтетического аппарата во многом обусловлена накоплением целого ряда микроэлементов. Среди них большое значение имеет марганец. Исключение марганца из питательной среды ведет к уменьшению количества хлоропластов, их объема, числа гран и содержания хлорофилла, в результате чего наблюдается хлороз листьев и резкое снижение интенсивности фотосинтеза. Прямое влияние марганца на фотоокисление воды связано, прежде всего, с тем, что он входит в состав марганецсодержащих белков, участвующих в процессе выделения кислорода [4]. Согласно предположению Л.Я. Леванидова, марганец осуществляет специфическую функцию – уравновешивание отрицательного потенциала, возникшего в результате накопления больших количеств сильных восстановителей. [5].
Таким образом, литературные данные свидетельствуют, что ведущей ролью марганца является его участие в реакциях фотосинтеза. Вместе с тем, данный микроэлемент имеет исключительно большое значение в поддержании окислительно-восстановительного потенциала клетки, играющим важную роль в устойчивости растительных организмов к неблагоприятным факторам среды. Учитывая эти физиолого-биохимические особенности марганца, представляло интерес исследовать динамику его содержания в зависимости от сезонности в хвое сосны обыкновенной, которая фотосинтезирует не только при положительных температурах, но и отрицательных.
Приведенные выше рассуждения позволяют считать актуальным изучение динамики содержания марганца и пигментов фотосинтеза в зависимости от абиотических факторов внешней среды.
Объектом исследования выбрана сосна обыкновенная ( Pinus silvestris L.) как представитель вечнозеленых растений, у которых фотосинтез не прекращается даже с понижением температур.
Полученные результаты могут иметь значение для исследователей, занимающихся изучением механизмов адаптации растительных организмов к низким температурам. Данные настоящей работы могут быть использованы в школе при подготовке к занятиям на темы «фотосинтез», «минеральное питание», а также при изучении действия экологических факторов на растительные организмы.
1. Литературный обзор
1.1. Механизмы защиты и устойчивости у растений
Изучение причин и механизмов устойчивости живого организма к различным повреждающим агентам и его адаптации к изменяющимся условиям окружающей среды представляет собой одну из важнейших задач биологической науки [6, с. 208]. Стремление к сохранению гомеостаза – важнейшее отличие живого организма от неживой материи. Именно благодаря этому свойству поведение биологического объекта не может быть описано только на основе физических законов и химических закономерностей, которым подчиняется неживая природа.
Учение о механизмах защиты и устойчивости растительных организмов к факторам окружающей среды наиболее полно сформулировано и развито [7]. По его меткому выражению "...способность растения к оптимальной устойчивости отражает меру его надежности".
Способность к защите от действия неблагоприятных абиотических и биотических факторов среды – столь же обязательное свойство любого организма, как питание, движение, размножение и др. Эта функция появилась одновременно с возникновением первых живых организмов и в ходе дальнейшей эволюции развивалась и совершенствовалась. Поскольку повреждающих и уничтожающих факторов множество, возникшие способы защиты от них оказались самыми разными – от метаболических механизмов до морфологических приспособлений (колючек и др). Выживаемость и расселение по новым экологическим нишам определялись способностью организмов приспосабливаться к необычным условиям среды. Адаптация, то есть приспособление организма к конкретным условиям существования, у индивидуума достигается за счет физиологических механизмов (физиологическая адаптация), а у популяции организмов (вида)– благодаря механизмам генетической изменчивости и наследственности (генетическая адаптация).
Надежность растительного организма проявляется в эффективности его защитных приспособлений, в его устойчивости к действию неблагоприятных факторов внешней среды: высокой и низкой температуре, недостатке кислорода, дефиците воды, засолении и загазованности среды, ионизирующих излучений, инфекции и др. [2].
1.2. Влияние температуры на пигментный аппарат хвои
В адаптации растений к различным повреждающим факторам, в настоящее время выделяют различные пути возможной регуляции данного процесса. Одним из путей низкотемпературной адаптации хвойных пород деревьев является изменение содержания пигментов фотосинтеза в зависимости от сезонности, так как деревья хвойных пород приспособлены к переживанию морозов и сохраняют иглы в течение нескольких лет.
Пигментный состав взрослой хвои сосны представлен двумя формами хлорофилла а и в, соотношения которых в хвое колеблется в зависимости от внешних факторов: от 1,5 до 3,0; и каротиноидами: b-каротином, лютеином и др. Возрастные колебания содержания зеленых пигментов довольно значительны. Первоначально это связано с развитием молодой хвои и накоплением в ней хлорофилла, а затем более сильным распадом хлорофилла в ней в зимнее время. К началу следующей вегетации количество хлорофилла в 1- и 2-летней хвое сравнивается. Для хвои более старших возрастов характерно более высокое содержание обоих форм хлорофилла. В тоже время возраст деревьев не влияет на количество зеленых пигментов в хвое. Видимо, на накопление пигментов большее влияние оказывают эдафические и климатические условия [8].
По своей природе в естественных условиях сосна обыкновенная является и холодо- и морозоустойчивым видом [там же]. Как вечнозеленое растение, сосна в течение года подвергается воздействию неблагоприятных факторов среды, особенно действию низких отрицательных температур. С переходом в состояние зимнего покоя происходит значительная деградация структуры хлоропластов, приводящая к разрушению пигмент-белковых комплексов (ПБК) и фотоокислению хлорофилла. Поэтому годичные изменения количества зеленых пигментов значительны.
Изменение окраски хвои весной происходит при установлении теплой погоды с положительными среднесуточными температурами воздуха и почвы и, видимо, совпадает с началом сокодвижения и возможно притоком цитокининов к хвое. Зимняя окраска хвои появляется при переходе к устойчивым отрицательным температурам [там же ].
Диапазон температур воздуха, при которых возможен фотосинтез хвои сосны обыкновенной, довольно широк: от -70 С до 530 С причем температуры выше 400 С вызывают временное подавление фотосинтеза. По данным ряда авторов прохладная погода благоприятствует накоплению органического вещества, так как при этом снижаются затраты на дыхание [9].
Продолжительность фотосинтетического периода у сосны зависит от климатических условий произрастания. Для умеренно холодного климата период ассимилирующей активности хвои довольно длинный. Так, в условиях Карелии фотосинтетическая активность у хвои отмечается в течение 7 месяцев [10]. Период низких температур необходим для жизнедеятельности сосны обыкновенной. Потребность в холоде может быть снижена лишь у растений, адаптированных к тепличным условиям или увеличенному фотопериоду.
Развитию зимостойкости способствует осеннее накопление в тканях сосны основных запасных веществ – сахаров, белков, жиров [11]. Накопившиеся к холодному времени года соединения, играющие в клетке роль криопротекторов (низкомолекулярные сахара и белки), не имеют столь большого значения для древесных растений по сравнению с травянистыми.
Характерным признаком морозостойкости растений, в том числе и сосны обыкновенной, является продолжение жизнедеятельности при отрицательной температуре, хотя и резко ослабленном виде. Способность к ассимиляции СО2 при фотосинтезе у сосны сохраняется при снижении температуры до -50 С. . Дыхание продолжается и при более низких температурах, вплоть до -200С. В зимний период, прежде чем дыхательный процесс полностью не инактивируется, возможен переход растений на анаэробный путь дыхания, менее чувствительный к повреждающему действию низких температур [12].
Даже слабо протекающий фотосинтез и дыхание позволяют сосне поддерживать более высокую внутриклеточную температуру по сравнению с температурой окружающей среды. При сравнении температуры ткани хвои и окружающего воздуха в первой декаде апреля отмечено, что при полном солнце и слабом ветре температура хвои превышала температуру воздуха на 12–140 С [8 ].
Адаптация к крайним условиям существования предполагает компенсаторные изменения обмена веществ, позволяющие поддерживать метаболические реакции на определенном уровне, обеспечивающем выполнение основных жизненных функций. Одним из наиболее широко распространенных способов низкотемпературной адаптации является изменение свойств ферментов, определяющих скорость и направленность метаболических процессов в организме. Следует отметить, что изменение активности и изоэнзимного спектра ферментов является общей ответной реакцией организма на любое стрессовое воздействие и поэтому неспецифично к холоду.
В целом изучение механизма низкотемпературной адаптации у различных климатипов сосны обыкновенной позволяет лучше понять особенности ее метаболизма в различных частях ареала и выявить физиологические и биохимические признаки устойчивости сосны к низкотемпературному фактору среды.
1.3. Физиологическое значение марганца
Наиболее широко распространенным способом адаптации к низким отрицательным температурам является изменение активности белков-ферментов. Хорошо известно, что в состав белков-ферментов входят микроэлементы. Среди них большое значение имеет марганец. Роль марганца и пигментов фотосинтеза в адаптации хвои сосны обыкновенной к низким температурам может проявляться как прямо, так и косвенно. Марганец входит в состав ферментов, которые катализируют синтез низкопротекторных соединений (свободной аминокислоты, полиаминов), играющих исключительно важную роль в морозоустойчивости растений. В этом выражается его опосредованное влияние. Прямое влияние марганца на процесс адаптации заключается в том, что он входит в состав флавиновых ферментов, участвующих в синтезе порфобиллиногена, а также участвует в упрочнении связи хлорофилл–белок, предохраняя пигмент от преждевременного окисления.
Большое значение для внутриклеточного обмена веществ имеют соединения марганца с ферментами, обуславливающие усиление окислительно-восстановительных процессов в растениях. Марганец благодаря своей способности к перемене валентности при окислении-восстановлении имеет существенное значение в процессе фотосинтеза. Положительное влияние этого элемента на фотосинтез показал ещё Мак-Арг (1922). Этот микроэлемент элемент способен регулировать направленность окислительно-восстановительных процессов в растениях в зависимости от различных условий питания: при аммиачном питании он действует как сильный окислитель, при нитратном питании – как восстановитель [13]. Так как фотосинтез является процессом окислительно-восстановительным, а марганец способен регулировать окислительно-восстановительные процессы, можно было предположить, что он будет оказывать существенное влияние на интенсивность фотосинтеза.
Марганец значительно повышает скорость потребления кислорода суспензией хлоропластов не только на свету, но и в темноте. Дефицит марганца (и меди) в растениях ведет к уменьшению содержания в них не только хлорофилла, но каротина и ксантофила. Из клеточных органелл больше всего марганца содержится в хлоропластах, около 30% всего количества марганца, находящегося в клетке [14; 15]. Весь марганец хлоропластов находится в прочносвязанном с белком состоянии, образуя соединения хелатного типа.
Экспериментальная часть
- Цель и задачи исследования
Цель работы: изучить динамику содержания марганца и пигментов фотосинтеза в хвое сосны обыкновенной в зависимости от сезонности.
Задачи:
Выявить динамику содержание основных и дополнительных пигментов в хвое сосны обыкновенной в зависимости от сезонности.
- Определить содержание марганца в хвое сосны обыкновенной фотометрическим методом в зависимости от сезонности.
- Провести статистическую обработку полученных данных, результаты свести в таблицы и оформить графически.
- На основании полученных результатов и теоретического материала предположить механизмы адаптации хвойных растений к действию неблагоприятных факторов внешней среды.
Объекты и методы исследования
Объектами исследования служили хвоинки сосны обыкновенной (Pinus silvestris L.).
Pinus silvestris L.- сосна обыкновенная- дерево 20-40 м высоты и 80-100 см в диаметре. Ствол в насаждении прямой, высоко очищенный от сучьев, на открытом месте сбежистый, ветвистый, с кроной, начинающейся почти у земли. Кора снизу ствола красно-бурая, глубоко-бороздчатая, в верхней части и на старых ветвях желтовато-красная, почти гладкая, на молодых ветвях серо-зеленая. Естественно произрастает почти по всей Европе (от Испании и Греции до Заполярья), в Сибири. Встречается в различных экологических условиях- на бедных и сухих песчаных почвах, на каменистых породах в горах, на торфяно-болотных почвах. Быстрорастущая древесная порода. Светолюбива и морозостойка.
Образцы хвои первого и второго года отбирали из средней части кроны 20-25-летних деревьев. Содержание основных и дополнительных пигментов определяли с помощью спектрофотометра СФ-26 по методике описанной в спиртовых растворах пигментов (этанол 96%). [16 ].
Концентрацию пигментов рассчитывали по формулам (Wintermans, De Mots, 1965):
а) для хлорофиллов
СА= 13,70*Д665 - 5,76*Д649
СВ= 25,80*Д649 - 7,66*Д665
СА+В = 6,10*Д665 + 20,04*Д649
б) для каротиноидов (Веттштейн,1949)
Скар=
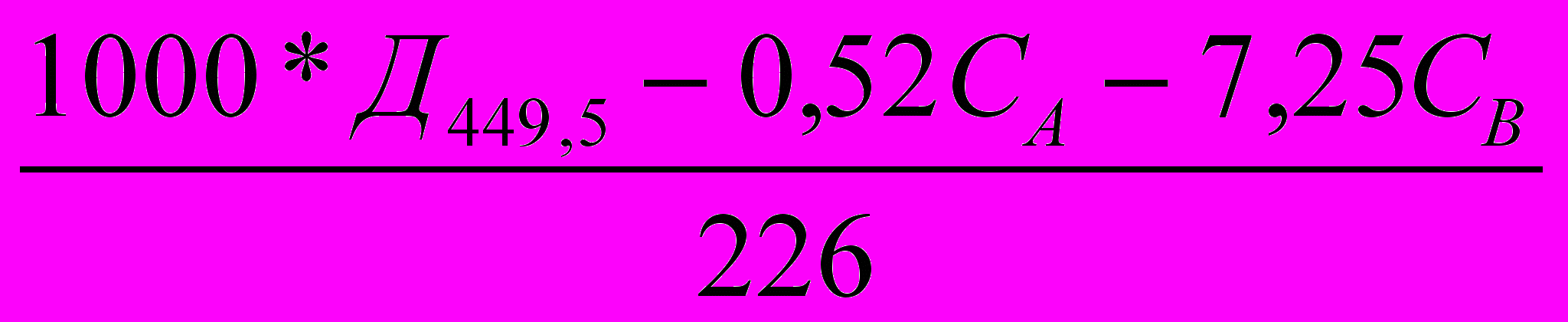
Содержание пигментов в растительном материале в мг/г сырого веса рассчитывали по формуле:
А =
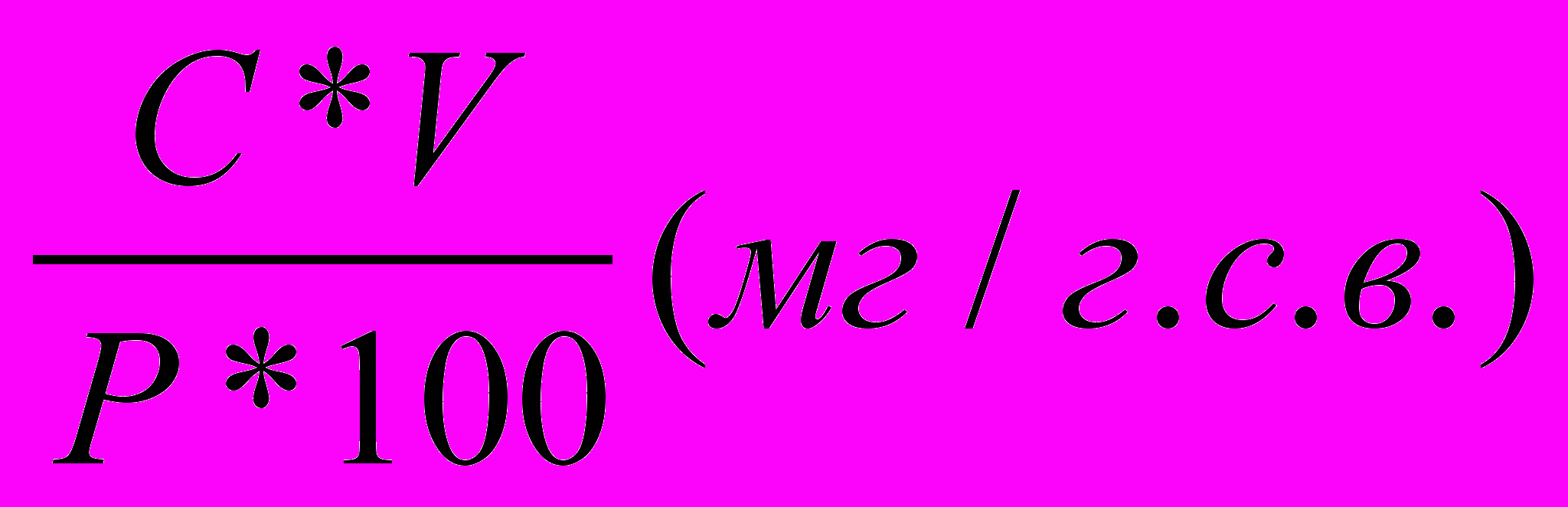 , где
, гдеС- концентрация пигмента в мг/л;
объем вытяжки пигмента в мл;
Р- навеска растительного материала в г;
А- содержание пигмента в растительном материале в мг/г сырого веса.
Содержание марганца определялось фотометрическим методом, описанным А.Я. Полухиной [13 .
Концентрацию полученного раствора вычисляли по методу сравнения оптической плотности стандартного и исследуемого растворов по уравнению:
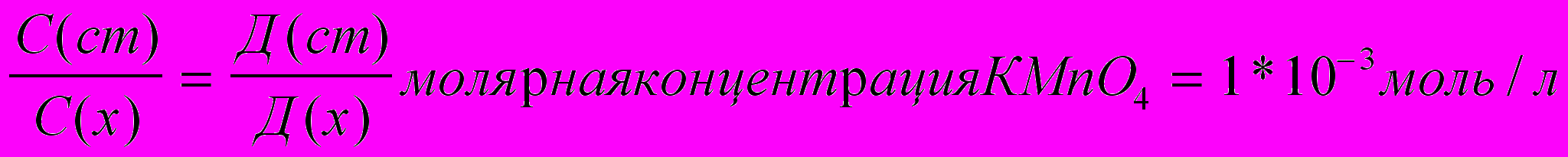
Массу марганца в мг/кг абсолютно сухого вещества вычисляли по формуле:
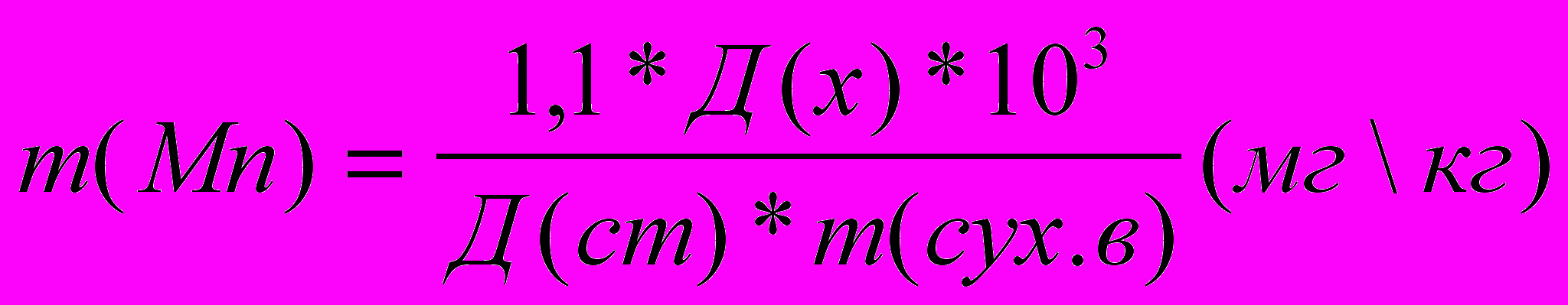
Эксперимент проводился в течение двух лет (2007–8 годы). Все опыты были проведены не менее чем в 4-х кратной биологической и химической повторностях. Результаты опытов обрабатывались статистически. Данные на рисунках представлены в виде средних значений со стандартными ошибками (n=5)
3. Результаты и их обсуждение
3.1. Влияние абиотических факторов на динамику пигментов фотосинтеза в хвое сосны обыкновенной в зависимости от сезонности
Эволюция фотосинтеза во многом предопределила не только высокий уровень организации растений, но и всех других организмов на Земле. Появление различных модификаций световых и темновых реакций явились одним из ответов на изменяющиеся условия внешней среды и позволили занять растительным организмам различные экологические ниши.
В процессе эволюции у хвойных вечно-зеленых растений в условиях континентального климата выработался ряд адаптивных реакций фотосинтеза. В связи с этим изучение сезонных и возрастных изменений первичных фотосинтетических процессов позволяет исследовать пластические возможности фотосинтетического аппарата и представляет значительный интерес для прикладных эколого-физиологических исследований.
Исследования пигментного состава фотосинтетического аппарата представлены на рис. 1–3; табл. 1. Как видно из рисунков содержание основных и дополнительных пигментов не остается постоянным в течение года. Максимальное их количество отмечается в летние месяцы. Это согласуется с данными Ю.Е. Новицкой, П.Ф. Чикиной [17 ], согласно которым с переходом в состояние зимнего покоя происходит значительная деградация структуры хлоропластов, приводящая к разрушению пигмент–белковых комплексов (ПБК) и фотоокислению хлорофилла.
По результатам этих исследований и литературным данным можно сказать, что возрастные колебания содержания зеленых пигментов довольно значительны. Для хвои II года характерно более высокое содержание обоих форм хлорофилла и каротиноидов по сравнению с хвоей I года. Первоначально это связано с развитием молодой хвои. Более низкое содержание пигментов в молодой хвое обусловлено с одной стороны тем, что в ней только начинает формироваться фотосинтетический аппарат в летний период ее развития, а с другой стороны частичной деградацией хлоропластов и пигментов в зимний период.
Эту общую закономерность подтверждают данные представленные на рис. 1, которые показывают содержание хлорофилла а в хвое сосны обыкновенной первого и второго года в зависимости от сезонности. Можно констатировать, что с апреля по август месяц происходит повышение содержания хлорофилла а. Именно в этот период начинается активный рост побегов для которых необходим энергетический и пластический материал.
Максимум содержания хлорофилла а приходится на август месяц, когда количество пигмента в хвое 1 и 2-го года выравнивается. Начиная же с сентября месяца происходит постепенное понижение содержания хлорофилла а в хвое как 1-го так и 2-го года. Уменьшение содержания хлорофилла в осенне-зимний период вероятно связано с подготовкой фотосинтетического аппарата хлорофилл-содержащих тканей к зимним условиям при переходе деревьев в состояние покоя. Та же самая зависимость наблюдается и в сезонном изменении содержания хлорофилла в в хвое 1-го и 2-го года (рис.2). Известно, что в зимний период происходит ряд изменений в организации и функционировании фотосинтетического аппарата хвои. Наблюдается пожелтение хвои, обусловленное фотоокислением хлорофиллов [18]. Данный автор считает, что причина зимнего обесцвечивания хлорофилла в том, что низкие температуры при достаточном освещении приводят к диспропорции между световыми и темновыми реакциями фотосинтеза и фотоокислению хлорофилла. Полученные в нашей работе результаты согласуются с данным А.А. Кривошеева, С.А. Шавнина и др. (1991), что сезонные изменения в содержании хлорофилла у сосны обыкновенной обусловлены, с одной стороны, изменениями физиологического состояния деревьев, а с другой – обратимым повреждением хлоропластов хвои в зимний период, поскольку деревья хвойных пород способны переносить низкие отрицательные температуры, сохраняя иглы в течение нескольких лет. При этом следует отметить, что у хвойных вечнозеленых растений было обнаружено наличие зимнего синтеза хлорофилла. В работах показано, что при отрицательной температуре (-20С) происходит включение С14 в молекулу хлорофилла Т.Н. Годнева [19]. В связи с этим высказывается предположение о том, что в цепи биосинтеза хлорофилла у хвойных имеется одно или несколько звеньев, образование которых из их предшественников, протекает при температуре не ниже -20 С. [20 ].
Изменение окраски хвои весной происходит при установлении теплой погоды с положительными среднесуточными температурами воздуха и почвы, и видимо, совпадает с началом сокодвижения и возможно притоком цитокининов к хвое. Это сопровождается повышением содержания обоих форм хлорофиллов (рис.2-3).
Интенсивность фотосинтеза зависит не только от содержания хлорофилла а и хлорофилла в, но и от их соотношения в хлоропластах. В этой связи сравнение коэффициентов, отражающих соотношение хлорофилла а и хлорофилла в в зависимости от сезонности представляет не сомненный интерес. Полученные в нашей работе данные (табл. 1) свидетельствуют, что соотношение хлорофилла а к хлорофиллу в колеблется в зависимости от внешних факторов от 3 (в зимний период) до 3,3 (в летний период). Эти результаты согласуются с данными Э.В.Ходасевич [18 ], что зимние изменения структуры хлоропластов хвои сосны обыкновенной проявляются в распаде гран и уменьшения числа тилакоидов в них до 2–8, тогда как в зрелом хлоропласте их количество достигает 7–20 на грану. В тоже время сохраняется целостность пластид. В большей степени разрушается хлорофилл в, поэтому увеличивается соотношение хлорофиллов. Зимой хлорофилл исчезает из тилакоидов гран, сохраняясь в межгранных ламеллах.
Кроме зеленых пигментов важную роль в процессе фотосинтеза играют каротиноиды, которые локализуются преимущественно в мембранах хлоропластов, где выполняют две важнейших функции: светособирающую и фотозащитную. Поглощая свет в сине-фиолетовой области (350-500 нм), где хлорофиллы поглощают мало, они являются дополнительными сборщиками световой энергии [21]. Хорошо известно, что каротиноиды, в частности b-каротин, является наиболее распространенным метаболитом живых организмов, учавствующих в системе защиты клеток от воздействия неблагоприятных факторов среды [22]. Учитывая важную роль каротиноидов в устойчивости клеток к экстремальным воздействиям представлялось важным изучение их динамики в зависимости от сезонности у хвойных растений.
Годичная динамика желтых пигментов-каротиноидов представленная на рис. 3 не имеет той же закономерности изменения, как у зеленых пигментов. Результаты полученные в нашей работе говорят о том, что количество каротиноидов в течении года изменяется в меньшей степени (в пределах 0,35–0,45 мг/г сырого веса). Хотя, как видно из рис.3, наблюдается некоторое уменьшение содержания каротиноидов в летние месяцы и нарастание его количества в зимние месяцы, как у хвои 1-го так и 2-го года. Результаты наших исследований частично согласуются с различными мнениями относительно динамики желтых пигментов в зимний период. По данным Э.В. Ходасевич [18 ], количество каротиноидов в хвое в зимний период меняется мало. Г.А. Ширяева (1967), изучавшая годичную динамику каротиноидов в хвое сосны, считает, что в условиях Ленинградской области отмечается максимум содержания каротиноидов в октябре–ноябре, а минимум летом, в период роста молодой хвои. По данным Ю.Е. Новицкой и П.Ф. Чикиной, в условиях Карелии у молодой хвои идет накопление пигментов с момента формирования до декабря, тогда как у хвои 2-го года вегетации количество каротиноидов относительно стабильно. Это объясняется тем, что каротиноиды помимо светособирающей функции выполняют еще и фотозащитную [17].
Роль каротиноидов в фотосинтезе обсуждается уже много лет, но однозначного решения этой проблемы, как оказалось, не существует. Всегда остается открытым вопрос: зачем фотосинтетической системе такой богатый набор каротиноидов? Одна функция каротинов – защитная, т.е. предохранение хлорофиллов светособирающей антенны ФС 11 от фотодеструкции, была известна еще на первых этапах исследования фотосинтеза. каротиноиды выполняют также роль структурного компонента кислородвыделяющей системы, соединяющей Р680 и марганцевый комплекс. Этот же каротиноид служит проводником электронов от выше указанного комплекса к Р680. Участие в переносе электрона и определенная локализация в кислородвыделяющей системе требует высокой степени ориентации каротиноидов в структуре хлоропластов. По данным работ Д.О. Сапожникова каротиноиды преобразуют энергию триплетного возбужденного состояния в теплоту.
Таким образом, полученные нами данные позволяют заключить, что динамика зеленых и желтых пигментов в зависимости от сезонности имеет противоположную направленность. По мере нарастания положительной температуры содержание обоих форм хлорофилла резко возрастает, а каротиноидов снижается. При нарастании отрицательных температур значительная часть хлорофиллов а и в разрушается, а содержание каротиноидов увеличивается. Сопоставление и анализ этих результатов и литературных данных позволяет предположить, что каротиноиды в зимний период выполняют не столько фотосинтетическую функцию, сколько защитную – переводя энергию квантов сине-фиолетовых лучей в теплоту. Желтые пигменты, по-видимому, наряду с белками холодового шока являются элементами термогенеза в растительных клетках, что обеспечивает повышение температуры хвои на 10–140 С выше температуры окружающей среды и осуществления фотосинтеза в зимних условиях.
3.2. Динамика содержания марганца в хвое сосны обыкновенной в зависимости от сезонности
Активность работы фотосинтетического аппарата зависит не только от содержания зеленых и желтых пигментов, но и от присутствия в его структурных компонентах элементов минерального питания. Минеральное питание – столь же уникальный процесс для растений, как и фотосинтез. Именно эти две функции лежат в основе автотрофности растительного организма. Кроме воды, поглощаемой из почвы и органических веществ, образующихся в процессе фотосинтеза, земному растению необходимо множество минеральных веществ. В клетках эти вещества выполняют целый ряд структурных функций, а также вовлекаются в состав специфических ферментов, регулирующих важные аспекты метаболизма клетки. Некоторые элементы минерального питания оказывают влияние на обмен веществ и устойчивость к неблагоприятным факторам окружающей среды. Хорошо известно, что содержание пигментов и активность работы фотосинтетического аппарата во многом обусловлена накоплением целого ряда микроэлементов. Среди них большое значение имеет марганец.
Необходимость минимальных количеств марганца для всех растений и в то же время его вредность в количествах, превосходящих определенный оптимум, доказана многочисленными опытами с водными культурами и в полевых условиях. Работы посвященные выяснению биохимической роли марганца, как правило, проводятся на растениях, поглощающих его в сравнительно небольших количествах [5]. Единственный автор, который пытался биохимически обосновать существование растений манганофилов был Успенский (1915). Он не выделял отдельно растений манганофилов, а применял к ним уже приведенное, действительное для всех растений положение, что марганец в малых количествах ускоряет развитие растений, но при повышенной концентрации угнетает и отравляет их. Успенский установил, что некоторые растения, такие как болотные растения, хвойные и др., могут переносить большой избыток марганца. Причиной этого вероятно является избыток органических кислот. Этот вопрос остается открытым и требует дальнейших исследований
Результаты наших исследований по изучению содержания марганца в хвое сосны обыкновенной в зависимости от сезонности представлены в таблице 2–3 и на рис.4. Изучение динамики накопления марганца в хвое сосны обыкновенной в различные месяцы года показало, что его количество в расчете на абсолютно сухой вес колеблется в зависимости от сезонности. Так, начиная с сентября содержание марганца резко увеличивается в хвое сосны обыкновенной II года и постепенно в хвое I года. Это согласуется с литературными данными по которым марганец повышает устойчивость фотосинтетического аппарата, где он наряду с каротиноидами играет ключевую роль в фотоокислении воды и выделении кислорода. Наряду с этим марганец способствует оттоку сахаров из листьев.
Увеличение содержания марганца в зимний период, с декабря по февраль, (период низких отрицательных температур) говорит о том, что повышенное его содержание может быть одной из причин адаптации растений к низкотемпературным условиям. Адаптация растений к воздействию различных повреждающих факторов идет по пути аккумуляции низкомолекулярных протекторных соединений (свободной аминокислоты, полиаминов и бетаинов), новообразования белковых макромолекул, а также формированию защитной системы против окислительного стресса. По мнению Л.Я. Леванидова высокая концентрация марганца, дающего соединения с высоким окислительным потенциалом, нужна растению для синтеза именно таких органических веществ с высоким восстановительным потенциалом. [13].
Следует отметить, что у всех механизмов адаптации имеется одна общая черта: участие белков-ферментов. Вовлечение белков-ферментов в процессы адаптации растений к холоду связано с изменением их синтеза и активности, что в свою очередь связано с изменением экспрессии соответствующих генов. Наконец изменение под действием низкой температуры экспрессии генов включает в себя ингибирование синтеза большинства экспрессирующихся в нормальных температурных условиях конститутивных белков и в тоже время происходит резкое усиление синтеза ряда стрессовых белков. В последние годы различными исследователями установлено, что эти белки играют важную роль в приобретении растением устойчивости к действию неблагоприятного температурного фактора, участвуя в различных защитных механизмах растительной клетки (Колесниченко и др., 2000). В ранних работах Шенье и Мартина указывалось на то, что функционально активный марганец в фотосинтетической мембране должен быть связан с белками с образованием хелатных соединений (Чернавская, 1988),и вероятно именно со стрессовыми белками.
На рис. 4 показано, что наибольшее количество марганца наблюдается в зимние месяцы (декабрь–февраль), т.е. в период глубокого покоя. В то же самое время (декабрь–февраль) содержание пигментов самое наименьшее (рис. 1–3). Результаты, полученные в нашей работе не согласуются с данными, в которых показано, что содержание марганца в листьях находятся в прямом соотношении с количеством хлорофилла. Так, в этиолированных листьях одуванчика находится 1,05 мг/100г с.в., в зеленых – 2,95 мг; в наружных (зеленых) листьях капусты – 2,55 мг/100г/с.в., тогда как во внутренних (белых) – только 0,46 (Липская, Годнева, 1963). По видимому следует оговориться, что сравнивать содержание марганца в листьях цветковых и хвойных растений не совсем корректно. Однако других литературных данных касающихся этого вопроса не было найдено.
Прямое влияние марганца на синтез хлорофилла связано, прежде всего, с тем, что он входит в состав флавиновых ферментов, участвующих в синтезе порфобиллиногена. Вместе с тем, данный микроэлемент участвует в упрочнении связи хлорофилл–белок, предохраняя пигмент от прежде временного разрушения. Этим можно объяснить повышение содержания марганца в зимние месяцы. При этом учитывается, что изменения активности фотосинтетического аппарата, происходящие в зимний период, обратимы, поскольку деревья хвойных пород приспособлены к переживанию морозов и сохраняют иглы в течение нескольких лет (Барская, 1967).
Как видно из рис. 4 содержание данного микроэлемента больше в хвое II года в зимнее время, чем в хвое I года в этот же период. Затем количество марганца в хвое обоих лет выравнивается. По-видимому, в весенние месяцы энергично синтезируются вещества, идущие на увеличение собственного веса хвои, а в дальнейшем, когда лист достигает нормальных размеров, идет энергичное накопление марганца, а, следовательно, вероятно и активное накопление стрессорных белков. Поэтому хвоя первого года (молодая) чаще подвергается разрушающему действию низких отрицательных температур в отличие от хвои второго года. Полученные результаты совпадают с данными Л.Я. Леванидова о накоплении марганца растениями в различные месяцы года. [5].
Таким образом, динамика содержания марганца в хвое сосны обыкновенной имеет прямую корреляцию с динамикой отрицательных температур, то есть чем ниже температуры окружающей среды, тем выше концентрация данного микроэлемента в изучаемом объекте.
Представляется интересным сравнение этих данных с результатами, полученными ранее нами, которые свидетельствуют, что подобная закономерность отмечается и в динамике каротиноидов. Динамика зеленых пигментов в те же годы имеет противоположную направленность. Сходная динамика марганца и каротиноидов, по-видимому, не случайна, т.к. известно, что данный микроэлемент повышает содержание каротиноидов. Особое внимание заслуживает тот факт, что марганец способствует увеличению содержания углеводов в иглах хвойных растений. Роль последних в устойчивости клеток к отрицательным температурам общеизвестна.
На основании полученных результатов и литературных данных можно констатировать, что участие марганца в упрочнении связи хлорофилл-белок и предохранение его от разрушения может быть одной из причин устойчивости хвойных к низким температурам и их способности осуществлять фотосинтез даже зимой. Марганец необходим для нормального протекания фотосинтеза, он участвует в процессе на этапе разложения воды и выделения кислорода (фотолиз воды), а также восстановления углекислого газа. Другой причиной устойчивости хвойных растений к низким отрицательным температурам может быть то, что функционально активный марганец в фотосинтетической мембране связан с белками, образуя хелатные соединения. Это дает основание для предположения, что некоторые из этих металлоорганических соединений играют роль в адаптации хвойных растений к низкотемпературному стрессу.
ВЫВОДЫ
Содержание зеленых и желтых пигментов не остается постоянным в течение года. Минимальное содержание хлорофиллов отмечается в зимний период, но полного разрушения хлорофиллов в этот период не наблюдается.
- Одна из причин относительно высокой устойчивости хвойных к низким отрицательным температурам, в частности сосны обыкновенной, может быть обусловлена повышением содержания каротиноидов в зимние месяцы.
- Повышенное содержание каротиноидов в зимний период обеспечивает повышение температуры хвои на 10–140 С по сравнению с температурой окружающей среды, что способствует осуществлению фотосинтеза в период низких отрицательных температур.
- Устойчивость хвойных вечнозеленых пород деревьев (сосна обыкновенная) к действию низких отрицательных температур обусловлена не только повышенным содержанием желтых пигментов, но и марганца, содержание которого в зимний период года также наибольшее.
- Высокое содержание активного марганца, входящего в состав фотосинтетической мембраны и связанного с белками (вероятно стрессовыми белками), по-видимому, способствует адаптации сосны обыкновенной к низким отрицательным температурам.
- В эксперименте четко прослеживается прямая корреляция между отрицательными температурами и содержанием каротиноидов и марганца. Это указывает на то, что они совместно принимают участие в адаптации хвойных к низким отрицательным температурам.
Литература
Полевой В.В. Физиология растений: Учеб. для биол. спец. вузов - М.: высш. шк., 1989 – 464 с.
- Селье Г.Очерки об адаптационном синдроме. Пер. с англ. В.И. Кандорова и А.А. Рогова. – М.: Медгиз, 1960. – С. 254.
- Болондинский В.К. Экофизиологические исследования динамики фотосинтеза у сосны обыкновенной// Эколого-физиологические исследования фотосинтеза и водного режима растений в полевых условиях- Иркутск: СиФИБР СО АИ СССР, 1983.
- Кутюрин В.М. Разложение воды при фотосинтезе как процесс биологического окисления. - В сб. Механизм дыхания, фотосинтеза и фиксации азота. М.: «Наука», 1967.
- Леванидов Л.Я., Давыдов С.Т. Марганец как микроэлемент в связи с биохимией и свойствами таннидов., Челябинское книжное издательство, 1961.
- Сарапульчев Б.И., Гераськин С.А..Генетические основы радиорезистентности и эволюция. – М..: Энергоатомиздат,1993. – С. 208.
- Гродзинский Д.М. Надежность растительных систем. – Киев, 1983. – 366 с.
- Физиология сосны обыкновенной /Судачкова Н.Е., Гирс Г.И., Прокушкин С.Г. и др.- Новосибирск: Наука. Сибирское отделение, 1990. – 248 с.
- Лир Х. Физиология древесных растений- М.: Лесная промышленность, 1974. – 420 с
- Молчанов А.А. Сосновый лес и влага. М.: Наука, 1983. – 139 с.
- Физиолого-биохимические механизмы роста хвойных. Ответв. редакторы канд. с. - х. наук Г.И. Гирс, канд. биол. наук Н.Е. Судачкова. Изд-во «Наука» Сибирское отделение. Новосибирск, 1978.
- Новицкая Ю.Е. Адаптация сосны к экстремальным факторам среды // Физиолого-биохимические основы роста и адаптации сосны на Севере. - Л.: Наука. Ленинградское отделение, 1985. – С. 113–138.
- Полухина А.Я., Леванидов Л.Я. Окисление двухвалентного марганца перекисью водорода в присутствии цитрат-иона /В сб. Биохимическая роль марганца в растениях. Челябинск, 1967.
- Рубин Б.А., Гавриленко В.Ф. Биохимия и физиология фотосинтеза. Издательство Московского университета 1977.
- Микроэлементы в жизни растений. – Л.: изд-во «Наука», Ленингр. отд. – 324 с.
- Гавриленко В.Ф., Ладыгина М.Е., Хандобина Л.М. Большой практикум по физиологии растений. Фотосинтез. Дыхание. Учебное пособие. М., «Высшая школа», 1975. – 392 с.
- Новицкая Ю.Е., Чикина П.Ф. Азотный обмен у сосны на Севере. - Л.: Наука. Ленинградское отделение, 1980. – 166 с.
- Ходасевич Э.В. Фотосинтетический аппарат хвойных: Онтогенетический аспект. - Мн: Наука и техника, 1982. – 199 с.
- Годнев Т.Н. Хлорофилл и его роль в природе (конспект лекций по избранным главам физиологии растений). – Минск.: изд-во Белгосуниверситета им. В.И. Ленина, 1955.
- Возилова Л.Д. Особенности биосинтеза хлорофилла в хвое проростков сосны обыкновенной (автореферат) Томск, 1971.
- Карнаухов В.Н. Биологические функции каротиноидов. М.: Наука, 1988. –240 с.
- Дубров А.П. Генетические и физиологические эффекты действия ультрафиолетовой радиации на высшие растения. М.: Наука, 1968. – 250 с.
- Похлебаев С.М., Похлебаев В.С. Физиолого-биохимические основы адаптации сосны обыкновенной к экстремальным абиотическим факторам среды обитания. / Экологический вестник Челябинской области. Издательство «СОХА», № 5-6 (16), 2003. – С. 97–109.
- Пол Д. Крамер, Теодор Т. Козловский. Физиология древесных растений / Documents and Settings\Дом.1-ABA9857062AF4, 7 января, 2003.
- Светлана А. Н., Д. А. Савчук. Климатогенная реакция деревьев сосны на юге Томской области /Журнал Сибирского Федерального университета: Серия «Биология», т. 1, № 4. – 2008.
ПРИЛОЖЕНИЕ
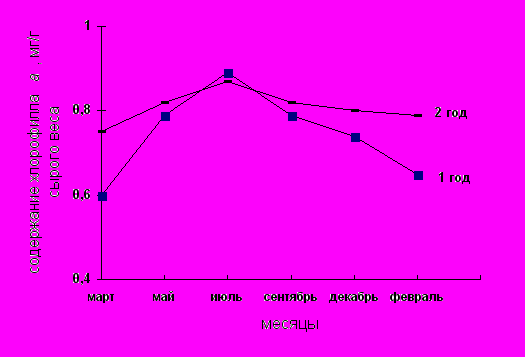
Рис. 1. Содержание хлорофилла а в хвое сосны обыкновенной первого и второго года
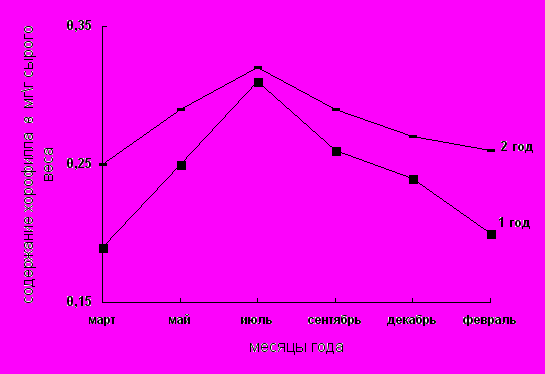
Рис. 2. Содержание хлорофилла в в хвое сосны обыкновенной первого и второго года
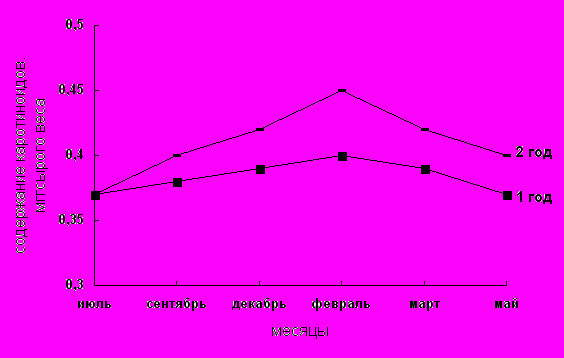
Рис.3. Содержание каротиноидов в хвое сосны обыкновенной первого и второго года
Таблица 1. Содержанию зеленых и желтых пигментов (мг/г сырого веса) в хвое I и II года сосны обыкновенной в зависимости от сезонности
| | март | май | июль | сентябрь | декабрь | февраль | ||||||
| | I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. | I г. | II г. |
| содержание хл.а | 0,6 | 0,75 | 0,79 | 0,82 | 0,89 | 0,87 | 0,79 | 0,82 | 0,74 | 0,8 | 0,65 | 0,79 |
| содержание хл в | 0,19 | 0,25 | 0,25 | 0,29 | 0,31 | 0,32 | 0,26 | 0,29 | 0,24 | 0,27 | 0,2 | 0,26 |
| содержание каротин-ов | 0,39 | 0,42 | 0,37 | 0,4 | 0,37 | 0,37 | 0,38 | 0,4 | 0,39 | 0,42 | 0,40 | 0,45 |
| отношение хл а/хл в | 3,16 | 3 | 3,16 | 2,8 | 2,9 | 2,7 | 3 | 2,8 | 3,1 | 3 | 3,25 | 3,16 |
Таблица 2. Содержание марганца в хвое I года сосны обыкновенной в зависимости от сезонности
| № п/п | месяцы | масса хвои, г | масса золы хвои, г | оптическая плотность, нм | масса Мn, мг/кг а.с.в. |
| 1 | август | 2,90 | 0,09 | 0,011 | 62,09 |
| 2 | октябрь | 1,03 | 0,03 | 0,009 | 142,8 |
| 3 | декабрь | 1,73 | 0,04 | 0,016 | 151,2 |
| 4 | февраль | 1,21 | 0,04 | 0,011 | 148,9 |
| 5 | апрель | 1,86 | 0,06 | 0,014 | 123,4 |
| 6 | май | 3,65 | 0,12 | 0,014 | 64,16 |
Таблица 3. Содержание марганца в хвое II года сосны обыкновенной в зависимости от сезонности
| № п/п | месяцы | масса хвои, г | масса золы хвои, г | оптическая плотность, нм | масса Мn, мг/кг а.с.в. |
| 1 | август | 2,80 | 0,07 | 0,010 | 66,3 |
| 2 | октябрь | 1,40 | 0,04 | 0,014 | 164,1 |
| 3 | декабрь | 1,20 | 0,03 | 0,017 | 232,4 |
| 4 | февраль | 1,84 | 0,04 | 0,026 | 231,4 |
| 5 | апрель | 1,56 | 0,05 | 0,014 | 147,3 |
| 6 | май | 3,28 | 0,11 | 0,014 | 70,1 |
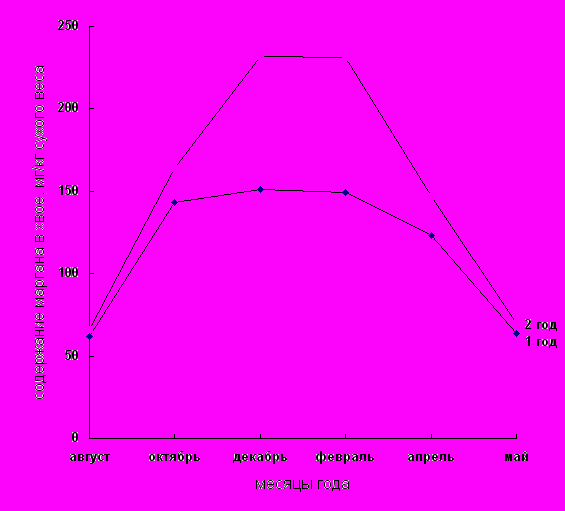
Рис. 4. Содержание марганца в хвое сосны обыкновенной первого и второго года в зависимости от сезонности