
Автореферат диссертации на соискание ученой степени
| Вид материала | Автореферат диссертации |
- Автореферат диссертации на соискание ученой степени, 378.33kb.
- Автореферат диссертации на соискание учёной степени, 846.35kb.
- Автореферат диссертации на соискание ученой степени, 267.76kb.
- Акинфиев Сергей Николаевич автореферат диссертации, 1335.17kb.
- L. в экосистемах баренцева моря >03. 02. 04 зоология 03. 02. 08 экология Автореферат, 302.63kb.
- Автореферат диссертации на соискание ученой степени, 645.65kb.
- Автореферат диссертации на соискание ученой степени, 678.39kb.
- Автореферат диссертации на соискание ученой степени, 331.91kb.
- Автореферат диссертации на соискание ученой степени, 298.92kb.
- Автореферат диссертации на соискание ученой степени, 500.38kb.
сосны обыкновенной
3.1. Характеристика изменчивости годичного радиального прироста деревьев
Длительность обобщенных древесно-кольцевых хронологий (ДКХ) по площадям варьирует от 124 лет до 163 лет (рис. 3). Средний возраст деревьев на площадках характеризуется сходными значениями от 108 до 119 лет. Сосны свыше 140–150 лет единично встречаются на контроле и ПП 12, наименьший возрастной состав модельных деревьев на ПП 5. В связи с разными возрастными характеристиками репрезентативный период с выборкой от 9–10 кернов по разным хронологиям представлен с 1896–1903 гг.
Средняя ширина годичного кольца (ШГК) у индивидуальных серий на контроле изменялась от 1.04 до 2.49 мм при 2.28 мм у обобщенной серии. Близкими к контролю размерами радиального годичного прироста характеризуются деревья ПП 12, их ШГК составила 1.47–2.41 мм у индивидуальных и 1.88 мм у обобщенной серий. Самый низкий средний прирост у деревьев ПП 5 – соответственно от 1.08 до 2.29 и 1.78 мм. Формы трендов обобщенных древесно-кольцевых хронологий по всем участкам идентичны и имеют классический вид, т.е. широкие годичные кольца формируются на начальном этапе роста дерева, затем их размеры постепенно снижаются. Период большого роста в среднем составляет в около 50 лет. Значимые отличия по форме возрастного тренда отдельных деревьев отсутствуют.
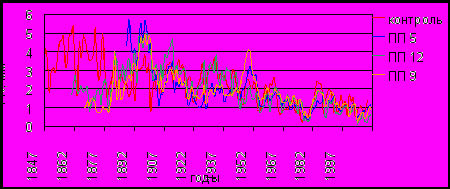
Рис. 3. Обобщенные древесно-кольцевые хронологии по абсолютным значениям прироста.
Таблица 2
Статистические характеристики обобщенных серий абсолютных (измеренных) древесно-кольцевых хронологий
| Статистические параметры | ПП 5 | ПП 9 | ПП 12 | ПП 11 (контроль) |
| Арифметическое среднее, мм | 1.78 | 1.86 | 1.88 | 2.28 |
| Ошибка среднего | 0.10 | 0.08 | 0.09 | 0.09 |
| Дисперсия выборки | 1.25 | 1.04 | 1.16 | 1.34 |
| Максимум, мм | 5.71 | 5.32 | 4.92 | 5.30 |
| Количество лет | 124 | 145 | 152 | 163 |
| t-критерий | 3.7* | 3.4* | 3.1* | |
Примечание.* – отличия с контролем достоверны при уровне значимости p < 0.05.
Поскольку абсолютные значения прироста содержат в себе информацию о возрастных изменениях, для их устранения используется процедура стандартизации или индексации (Битвинкас, 1974; Fritts, 1976; Ловелиус, 1979; Briffa et al., 1986; Cook, Kairiukstis, 1990; Ваганов, Шиятов, Мазепа, 1996; и др.). Индексы определяются делением абсолютных значений ширины кольца на значения индивидуально подобранной возрастной кривой по формуле:
It = wt/w–t,
где I – индекс прироста, w – фактическая ширина годичного кольца, w– – значение, предсказанное аппроксимирующей кривой, все для года t. Аппроксимация выполнена с помощью негативной экспоненты или линейной регрессии.
В результате расчета индексов получены стандартизированные обобщенные ДКХ (рис.4).
Высокие межсериальные коэффициенты корреляции (от r = 0.51 (ПП 5) до r = 0.8 (контроль)) свидетельствуют о синхронной реакции деревьев на влияние внешних условий. Наибольшие значения коэффициентов между сериями годичных проростов у отдельных деревьев контрольного участка, наименьшие – между деревьями на площади 5 с экстремальным значением СПЗ. Стандартное отклонение, которое характеризует амплитуду изменчивости прироста от условий среды, у обобщенных хронологий с разных площадей имеет близкие значения – 0.33–0.35, а средняя чувствительность (оценивает степень воздействия внешних факторов природной среды по относительным различиям размеров соседних годичных колец индивидуальных хронологий) сосен контрольной и ПП 12 выше, чем у обобщенных серий площадей 5 и 9 (0.33 и 0.33; 0.22 и 0.23 соответственно). Так как рассчитанные статистические показатели выше пороговых (> 0.2), хронологии со всех площадей можно охарактеризовать как чувствительные к внешним воздействиям среды.
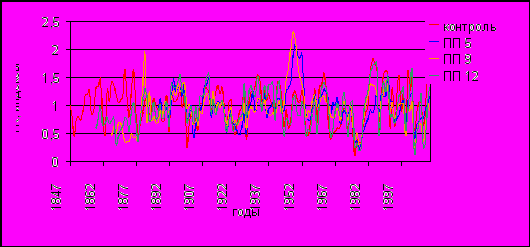
Рис. 4. Обобщенные стандартные древесно-кольцевые хронологии по пробным площадям.
Все ДКХ имеют высокую автокорреляцию -го порядка. Это говорит о том, что размеры прироста исследуемых деревьев испытывают значительное влияние условий роста предыдущего года. Средняя автокорреляция у обобщенных серий стандартных хронологий изменяется от 0.40 на контроле до 0.71 на ПП 5. У индивидуальных серий автокорреляция имеет разброс значений от 0.32 (ПП 12) до 0.83 (ПП 5), при этом средние показатели индивидуальных серий в среднем на 9 % (от 0.4 до 14 % по участкам) выше обобщенных.
Выполнен анализ цикличности и лет с экстремальными приростами по остаточным ДКХ, так как в них максимально выражена погодичная изменчивость, а годы экстремумов выделяются более четко. Остаточные хронологии получены с помощью «выбеливания» стандартизированных рядов (устранение влияния прироста предшествующих лет на прирост последующего).
Высокочастотные значимые циклы длительностью 5–7 лет выявлены только для индексированных хронологий на контроле и ПП 12. При 5-летнем сглаживании индексов прироста максимальный пик спектральной плотности у разных хронологий отмечен на частотах, соответствующих циклам в 10–13 лет, близким к 11-летним циклам солнечной активности. Такой сигнал является общим для всех исследованных древесно-кольцевых хронологий. В динамике прироста деревьев со всех площадей имеют место также циклы средней продолжительностью в 17–18 лет. При 10- и 15-летнем сглаживании выявляются циклы, средняя длительность которых колеблется в интервалах 30–41 год. Пик спектральной плотности на частоте, соответствующей циклу в 62 года, отмечен у древесно-кольцевой хронологии ПП 5. Выделить более длительные циклы не позволяет возраст деревьев.
В динамике ШГК за весь период исследования установлены надежные реперные годы с минимальным приростом: 1916, 1918, 1927, 1931, 1950, 1964, 1967–1968, 1970–1972, 1978, 1987, 1990, 1994, 1999, 2003. В эти годы прирост снижался у деревьев на всех ПП. Наиболее значительные падения прироста приходятся на 1909–1920, 1960–е–1970-е и 1994–2003 годы.
3.2. Анализ динамики годичного радиального прироста деревьев в связи с воздействием природно-антропогенных факторов
Анализ отклика обобщенных стандартных ДКХ на метеопараметры (температура воздуха, атмосферные осадки и производный от них гидротермический индекс увлажнения) выполнен с помощью корреляционного анализа по самой короткой хронологии с 1903 по 2007 г., представленной достаточным количеством кернов. В расчеты включены значения климатических переменных за все месяцы текущего года и с сентября по декабрь года предшествующего. Анализ функций отклика показал, что доминирующее влияние на радиальный прирост сосны оказывает годовая сумма осадков. Значения коэффициента корреляции составили от 0.26 (ПП 5) до 0.45 (ПП 9) при доверительном интервале за пределами значений ±0.2 (при р < 0.05). Осадки сентября предшествующего года также оказывают положительное влияние по всем хронологиям, что свидетельствует о лимитирующем значении запаса влаги, содержащейся в почве к моменту начала камбиальной активности. Значимые коэффициенты корреляции отмечаются в динамике прироста деревьев контрольного участка с суммой осадков за отдельные месяцы теплого периода: май (r = 0.27), июнь (r = 0.4) и сентябрь текущего года (r = 0.23). На ПП 12 сосновые деревья характеризуются положительной реакцией ШГК на осадки мая и июня (r = 0.3 и 0.46 соответственно) при одновременном отрицательном воздействии июньских температур воздуха (r = –0.29). У хронологий ПП 5 и ПП 9 достоверной связи с осадками за отдельные месяцы весенне-летнего периода не обнаружено. В других случаях влияние температуры корреляционным анализом не выявлено. Важным показателем влагообеспеченности является гидротермический коэффициент увлажнения (ГТИ), который рассчитывался нами как отношение суммы осадков сентября предшествующего года и июня-сентября года вегетации к средней температуре тех же месяцев. Установлена положительная корреляция у всех ДКХ со средним значением ГТИ за период с мая по сентябрь текущего года прироста. При этом значения r были выше, чем с осадками, и составляли от 0.43 на ПП 5 и до 0.54 на контроле. Значимые связи с коэффициентом увлажнения августа для ДКХ со всех участков отсутствуют.
В дополнение к корреляционному анализу для выявления лимитирующих факторов в годы с экстремальными (минимальными и максимальными) величинами индексов прироста был использован анализ климаграмм локальных экстремумов прироста (Ловелиус, 1979, 2003; Румянцев, Николаев, 2006; Николаев, 2010). Индексы для анализа вычислялись как отклонение значений прироста данного года от среднего за 5 лет, как это принято в лесоводстве и лесной таксации, по формуле (Румянцев, 2010):
It = Wt / Yt,
где I – индекс прироста, W – фактическая ширина годичного кольца, Y – средняя ширина годичного кольца за 5 лет, все для года t.
Климаграммы построены по средним месячным суммам атмосферных осадков (мм) и температур воздуха (ºС) за период вегетации и относительного покоя деревьев. Анализ их показал, что разницы в реакции на температурный режим в течение года между группами с разной интенсивностью прироста не прослеживается, в то время как благоприятными для роста сосны по всем участкам являются осадки. Согласно t-критерию существуют достоверные отличия по суммам осадков мая, июня и июля текущего, а также сентября предшествующего года между двумя средними по экстремальным годам значениями прироста для всех ДКХ с превышением в 1.2–1.7 раза. При этом, наибольшее расхождение по количеству осадков отмечено для мая. Для августа, который наряду с июлем наиболее обилен атмосферными осадками, по t-критерию отличия недостоверны, что указывает на формирование достаточного для роста дерева запасов почвенной влаги в предшествующие месяцы.
Анализ климаграмм подтверждает результаты корреляционного анализа и дает основание заключить, что внутригодовое распределение осадков в теплый период имеет наибольшее значение в годы с аномальным приростом деревьев. По всей видимости, именно водный стресс в начале периода вегетации (в мае) имеет определяющее значение для размеров годичного радиального прироста сосны в исследуемых условиях.
Выявления реакции радиального прироста на воздействие техногенного загрязнения базировалось на сравнительной оценке ШГК по ряду параметров за 2 периода времени – с 1934 по 1970 г. и с 1971 по 2007 г. (37 лет до предполагаемой ответной реакции и 37 лет после). Сопоставление статистических характеристик изменения ширины колец между хронологиями разных площадей показывает, что с 1971 г. на ПП, попадающих под загрязнение разного уровня, отмечается достоверное снижение средних размеров ШГК. Отмечено, что большие размеры годичных колец на контрольной площади связаны с более высокими максимальными значениями, тогда как минимальные сопоставимы с другими площадями. Помимо этого, данная хронология характеризуется увеличением дисперсии по сравнению с остальными абсолютными ДКХ, что может объясняться повышенной реактивностью здоровых насаждений, позволяющей им четко реагировать на изменение климатических факторов (Ловелиус, 1979; Демаков, Медведкова, 2009). Таким образом, при загрязнении темпы прироста сосны в благоприятные по климатическим условиям годы по сравнению с контролем снижаются.
Имеет место увеличение корреляции величины приростов между площадями по периодам от 0.34–0.87 (1934–1970 гг.) до 0.78–0.92 (1971–2007 гг.). Это объясняется совпадением выделенного нами антропогенного периода (1971–2007 гг.) с климатическими изменениями, характеризующимися ростом температур воздуха и засушливой фазой в выпадении атмосферных осадков. По стандартизированным сериям в целом прослеживаются сходные тенденции, но отмечается сглаживание негативного линейного тренда, который наблюдается у обобщенных ДКХ по абсолютным значениям, где он, по всей видимости, связан с влиянием возрастных особенностей деревьев.
Анализ цикличности ДКХ в зависимости от уровня СПЗ показал, что на контроле характерно проявление более высокочастотных циклов, которые наибольшим образом соответствуют периодичности в режиме выпадения атмосферных осадков. В частности, пики спектральной плотности для приростов с 1903 года отмечаются на 5.4 года и 10.8 лет, такие же цифры получены для атмосферных осадков за тот же период времени. Аналогичные высокочастотные циклы выделены для обобщённой хронологии ПП 12, но показатели их спектральной плотности ниже. Для хронологий ПП 9 и ПП 5 слабый сигнал спектральной плотности начинает фиксироваться со значений в 11.3 лет и 13.1 лет, достоверные значения отмечены на пиках в 17.7 лет и 18.0 лет соответственно.
Анализ климатического отклика на площадях с разной интенсивностью техногенной нагрузки показал, что в большинстве случаев второй период характеризуется более интенсивным откликом радиального прироста на влияние климатических параметров. Показательно, что в этот период возрастает положительное влияние атмосферных осадков на прирост сосны. По всем участкам отмечаются значения коэффициентов корреляции от 0.41 до 0.60 (доверительный интервал за пределами ±0.33 при р < 0.05) с суммой атмосферных осадков за год. За весь период наблюдений ДКХ на контрольной ПП характеризуется более высокими значениями отклика прироста на климатические параметры. Наименьшая реакция на метеопараметры у древесно-кольцевой хронологии ПП 5, где уровень загрязнения максимальный. Другие закономерности в откликах деревьев на природно-техногенные условия не выявляются.
ГЛАВА 4. Морфобиологические параметры вегетативных и генеративных органов
4.1. Морфологическая характеристика вегетативных органов
Отбор хвои на пробных площадях проводился в 2007 г. и 2008 г. с генеративных побегов высоких порядков ветвления в нижней освещенной (преимущественно южной) части кроны, для исследования использовалась хвоя второго года вегетации. Средние размеры хвои изменялись в пределах от 44.5±0.13 до 67.8±0.21 мм (табл. 2). Как минимальная (40 мм), так и максимальная (82 мм) длина отмечена на ПП 2 для хвои 2007 г. Какие-либо закономерности в изменении длины хвои в зависимости от интенсивности загрязнения по площадям не выявляются. Коэффициенты вариации исследуемого признака соответствует очень низкому и низкому уровню изменчивости, что свидетельствует о его стабильности.
Из сравнения длины хвои с метеопараметрами за период вегетации установлена следующая зависимость. Вегетационный период в 2007 году характеризовался наименьшей суммой осадков (129 мм) по сравнению с 2006 г. (221 мм) и 2008 г. (362 мм), тогда как сумма среднемесячных температур была наибольшей – соответственно 80.8; 69.7 и 71.7 ºС. Хвоя, сформированная в этом году, имела меньшую среднюю длину по всем пробным площадям в сравнении с хвоей предыдущего года формирования, когда погодные условия были более благоприятными. Различие между длиной хвои исследуемых лет достоверное – tф = 4.9 при tst = 2.31 (р < 0.05). Очевидно, что осадки и температуры воздуха в первый год вегетации оказывают определяющее влияние на длину хвои.
Таблица 2.
Длина двухлетней хвои (мм) у деревьев сосны за два периода вегетации
| № ПП | Вегетация 2006–2007 гг. | Вегетация 2007–2008 гг. | ||||||
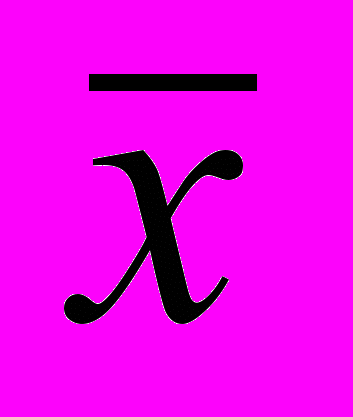 ±m ±m | Сv, % | X min | X max | ±m | Сv, % | X min | X max | |
| 1 | 56.7±0.17 | 10 | 42 | 72 | 50.3±0.16 | 10 | 35 | 63 |
| 2 | 62.9±0.33 | 17 | 40 | 82 | 56.8±0.17 | 9 | 42 | 78 |
| 3 | 58.2±0.15 | 8 | 48 | 74 | 57±0.29 | 15 | 35 | 70 |
| 4 | 63.2±0.24 | 12 | 45 | 80 | 50.7±0.19 | 12 | 35 | 63 |
| 5 | 67±0.18 | 9 | 54 | 81 | 59.9±0.18 | 9 | 43 | 70 |
| 7 | 50.3±0.17 | 10 | 41 | 61 | 44.5±0.13 | 9 | 39 | 52 |
| 8 | 52.0±0.08 | 5 | 45 | 59 | 51.9±0.2 | 12 | 44 | 64 |
| 9 | 67.8±0.21 | 10 | 55 | 85 | 61.5±0.11 | 6 | 53 | 73 |
| контроль | 63.0±0.19 | 10 | 48 | 76 | 55.5±0.25 | 14 | 35 | 75 |
Примечание.
±m — среднее значение и его ошибка, Сv — коэффициент вариации, X min – X max – лимиты признака.Для сравнительной характеристики линейного прироста побегов по пробным площадям принято использовать величину прироста второго года. В фоновых условиях по годам она варьировала от 1.4 см до 5.9 см при среднем значении 3.2±0.38 см и 3.9±0.39 см. По остальным площадкам максимальные приросты на ПП 1 (10.8 см) и ПП 2 (11 см), наименьшие – на ПП 4 (1.4 см). Средние значения на площадках с различным уровнем загрязнения варьируют в пределах от 3.2±0.42 см (ПП 4) см до 6.1±0.69 см (ПП 1). Какой-либо закономерности в изменении длины побега второго года в зависимости от интенсивности загрязнения по площадям установить не удалось.
Для выявления зависимости линейного прироста побегов от температуры воздуха и атмосферных осадков проанализированы размеры прироста за 5-летний период (с 2003 г. по 2007 г.) (рис. 5). Высокие значения приростов наблюдались в 2003, 2005 и 2006 годах, а низкие пришлись на 2004 и 2007 гг. Поскольку все участки обладают высокой синхронностью годичных изменений и дифференциации от степени загрязнения не отмечается, это позволяет заключить, что определяющим фактором линейных размеров побега являются климатические условия. Корреляционный анализ климатических показателей с усредненными по всем площадям размерами прироста за пятилетний период показал, что доминирующее влияние на прирост побегов оказывает сумма осадков периода вегетации. Немаловажное значение имеет и отношение сумм осадков к суммам температур вегетативного периода. В годы большего прироста оно равнялось 3.3 (2003 г.), 5.1 (2005 г.), 3.2 (2006 г.), а в годы низкого прироста снижалось до 2.0 (2004 г.) и 1.6 (2007 г.).
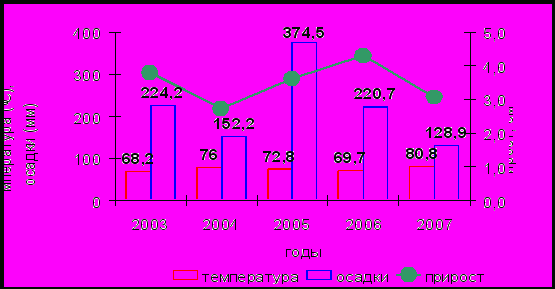
Рис. 5. Зависимость среднего по площадям линейного прироста (см) побегов от сумм среднемесячных температур и осадков за период вегетации.
4.2. Морфобиологическая характеристика генеративных органов
Сбор зрелых макростробилов проводился в 2008 и 2009 гг. во второй декаде марта до начала естественного выпадения семян, с нижней освещенной части кроны. По каждой площадке формировался смешанный образец из 35–50 штук.
Окраска макростробилов варьировала по площадям от серо-зеленой (преобладающая) до темно-коричневой. Данный признак не связан с влиянием внешних факторов, а является результатом эндогенной изменчивости (Мамаев, 1972).
Высота макростробилов на фоновой площади в среднем составляла 3.1±0.08 см при первом сборе и 1.5±0.03 см при втором. По остальным площадям она соответственно изменялась от 2.9±0.05 до 3.9±0.14 см и от 1.2±0.02 до 1.8±0.03 см. Характерно, что на ПП 9, которая попадает в контур очень высокого уровня загрязнения (СПЗ – 257–768), за оба исследуемых периода высота шишек была наибольшей (рис. 6). Явные различия по годам по всем площадкам с разницей в 2–2.6 раза связаны с ухудшением температурного режима и условий увлажнения. Закладка шишек урожая 2009 года происходила в 2007 году, когда климатические условия характеризовались низкой суммой осадков и высокими суммами температур вегетационного периода (Р = 128.9 мм, Т = 80.8 ºС), в то время как шишки урожая 2008 года закладывались в 2006 году, когда условия для роста были более благоприятны (Р = 221 мм, Т = 70 ºС).
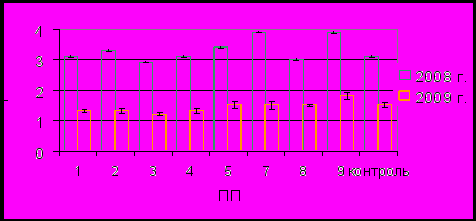
Рис. 6. Средняя высота макростробила по площадям.
Помимо этого макросторобилы урожая 2009 г. характеризуются снижением выхода семян, приходящихся на одну шишку, тогда как масса и размеры семян, напротив, были больше.
Масса 1000 зрелых семян урожая 2008 г. составляла 4.1–6.4 г, а 2009 г. – от 4.8 до 8.9 г. Достоверных различий массы семян от уровня загрязнения не обнаружено.
Размеры семян и крылаток. Для урожая 2008 г. минимальная длина семени была 2.3 мм, максимальная – 6.5 мм при средних величинах 3.1–4.5 мм. В 2009 г. разброс меньше – от 2.1 до 5.2 мм при средних по площадям 3.1–4.2 мм. Другие линейные характеристики семян и крылаток имели тенденцию к увеличению у урожая 2009 г. Отмечалось наличие в выборках с каждой площади от 1 до 3 % нормально развитых летучек при отсутствии сформировавшихся семян. Данные измерений семян и крылаток показывают, что, несмотря на некоторое увеличение на контроле, их линейные размеры не обнаруживают взаимосвязи с зонами загрязнения и, скорее всего, отражают индивидуальные особенности отдельных деревьев и экологические условия местообитания.
Семена со всех площадок имели преимущественно светлую серую и серо-коричневую окраску кожуры, что подтверждает вывод В.Л. Черепнина (1980) о связи светлоокрашенных семян с малым количеством годовых осадков и низким классом бонитета (не выше III).
Форма семян и крылаток по площадям сохранялась от года к году, что подтверждает вывод других исследователей о генотипической заложенности этого признака.
Результаты проращивания семян показали, что почти все здоровые семена сбора 2008 г. проросли в течение 5–7 суток, в оставшиеся дни учета проростки образовали только единицы (рис. 7). Несколько сниженной скоростью прорастания за весь период эксперимента отличались семена, отобранные с деревьев ПП 5, попадающей в зону с экстремальным уровнем загрязнения (СПЗ выше 769 ед.) и ПП 7, где СПЗ составил 65–128 ед. (загрязнение среднее). Результаты по ПП 5 по дням характеризовались 74.7; 5.7; 3.7 и 0.0 % прорастаний, а по ПП 7 – 78.5; 8.0; 2.0 и 0.0 % на 5-е, 7-е, 10-е и 15-е сутки соответственно. Энергия прорастания по площадкам у семян урожая 2008 г. составила от 80 до 96 % при 87 % на контроле (табл. 3). В связи с тем, что у основного количества семян нормальные проростки были образованы уже в первые дни эксперимента, энергия прорастания и всхожесть характеризуются сходными значениями, расхождения не более 2–5 % наблюдаются только по семенам с пробных площадей 2. 5 и 7. Расчет t-критерия Стьюдента показал достоверные различия между контрольной и остальными площадями.
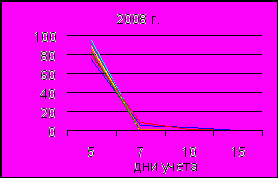 | 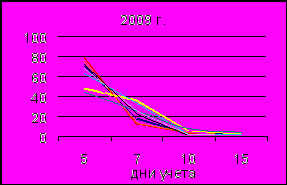 |
Рис. 7. Прорастание семян по дням учета в 2008 г. и 2009 г.
Иная картина получена при определении качества семян, собранных в 2009 г. Образование проростков отличалось большей временной растянутостью. В первый день учета появилось в среднем на 15–52 % (площади 2 и 8) меньше проростков, чем в опыте по предыдущему году. Энергия прорастания варьировала в пределах 69–96 % (площади 8 и 7) при 94 % на контроле, а всхожесть от 80 % (ПП 8) до 95 % (ПП 9 и контроль). В связи с удлинением срока прорастания разница между этими двумя показателями составила от 3 % (ПП 2 и 7) до 16 % (ПП 8). Исключением были площади 9, по которой все семена проросли уже на 2 учетный день, и контрольная, где разница не превышала 1 %. Для семян урожая 2009 г. выявлены достоверные расхождения с контролем: по энергии прорастания – площади 1, 3, 4, 5 и 8, а по всхожести – площади 5, 7 и 8.
Ухудшение показателей, характеризующих качество семян, созревших в 2009 году, связано, по всей видимости, с влиянием погодно-климатических условий в год закладки семян. Отмечается, что наряду с различием по годам, у семян этого урожая выявляется дифференциация выборок в зависимости от уровня загрязнения площадок. По всей видимости, негативное влияние климатических факторов в год закладки семян повлекло за собой ослабление их устойчивости к таким внешним факторам, как техногенное загрязнение. Наибольшей чувствительностью к влиянию внешних пессимальных условий среды характеризуется энергия прорастания семян.
Таблица 3
Оценка достоверности расхождения с контролем показателей проращивания семян
| № ПП | Энергия прорастания, % | Всхожесть, % | ||||||
| 2008 г. | t-критерий | 2009 г. | t-критерий | 2008 г. | t-критерий | 2009 г. | t-критерий | |
| 1 | 95 | 1.5 | 88 | -4.0* | 95 | 1.6 | 93 | -1.1 |
| 2 | 96 | 2.0 | 91 | -1.7 | 98 | 2.3 | 94 | -0.3 |
| 3 | 89 | 0.2 | 84 | -7.8* | 89 | 0.2 | 91 | -1.2 |
| 4 | 95 | 1.6 | 88 | -5.2* | 95 | 1.6 | 92 | -1.5 |
| 5 | 80 | -1.4 | 79 | -7.1* | 84 | -0.8 | 84 | -4.5* |
| 7 | 86 | -0.3 | 96 | 0.9 | 88 | 0.2 | 99 | 3.0* |
| 8 | 93 | 1.1 | 69 | -14.1* | 93 | 1.1 | 80 | -6.2* |
| 9 | 83 | -1.0 | 93 | 0.5 | 83 | -1.0 | 95 | 0.0 |
| контроль | 87 | | 94 | | 87 | | 95 | |
Примечание. * – достоверные по t-критерию различия с контролем (при p < 0.05).