
Рыбы открытого океана
| Вид материала | Документы |
- Казимир Малевич, я начало всего, 829.32kb.
- Конспект открытого урока биологии в 7 классе по теме: «класс хрящевые рыбы», 39.29kb.
- Урок-викторина по теме Животные. Класс рыбы, 75.59kb.
- Конспект открытого урока биологии в 7 классе по теме: «класс хрящевые рыбы», 48.22kb.
- Рельеф дна Мирового океана, 52.6kb.
- Урок биологии в 7 классе. Тема: «Внутреннее строение рыб», 112.94kb.
- Процессы и производственные линии замораживания продуктов растительного происхождения, 145.64kb.
- Тема урока Северный Ледовитый океан; особенности природы, хозяйственного использования,, 53.05kb.
- Магистерская специальность «Экономика ресурсов Мирового океана», 26.67kb.
- А. В. Клименко конкурсная документация по проведению открытого конкурс, 1840.42kb.
Большой алепизавр (Alepisaurus ferox) (рис. 25) имеет очень широкое распространение. Он встречается как в тропических, так и в умеренных водах в очень растянутом диапазоне глубин — от подповерхностных горизонтов до 1000 м, будучи особенно обычным в слое от 150 до 350 м, и часто ловится на крючки тунцеловных ярусов. Максимальная зарегистрированная длина этой рыбы 210 см (второй вид рода — короткорылый алепизавр А. brevirostris не превышает 90 см). Экземпляры длиной 1,5 м и массой около 5 кг (по соотношению этих величин видно, насколько прогонисто их тело) вполне обычны в уловах. Алепизавры — прожорливые хищники, не очень разборчивые в выборе добычи. В списках их жертв значатся десятки видов рыб, головоногих моллюсков, ракообразных, а также сальпы, гребневики, медузы, пелагические полихеты и другие объекты. Следует отметить, что переваривание пищи у алепизавров (как и у родственных им рыб) происходит только в кишечнике, а желудок используется лишь в качестве своего рода «провизионной кладовой» и всегда содержит абсолютно целые объекты питания (из этого «орудия лова» происходит немало голотипов мезопелагических животных). Общее число жертв в желудке может быть очень велико: у одного алепизавра, например, там были найдены 41 рыба, 3 кальмара, 1 амфипода и 1 полихета, а у другого сразу 83 экземпляра рыб-топориков. Трофологические исследования показывают, что именно алепизавры являются главными пищевыми конкурентами «ярусных» тупцов, но они же принадлежат к числу первостепенных кормовых объектов этих тунцов.
Среди мезопелагических хищников нельзя не упомянуть небольших акул из преимущественно бентопелаги-ческого семейства катрановых (Squalidae), представляющих довольно специфический элемент океанической ихтиофауны. Их насчитывается около 10 видов. Акула-карлик (Euprotomicrus bispinatus) встречается во всех океанах и приурочена преимущественно к центральным водным массам (в других районах открытого океана ее замещает близкий и также карликовый вид — акула-пигмей Squaliolus laticaudus). Акула-карлик принадлежит к числу никтоэпипелагических рыб, мигрирующих ночью к поверхности (акула-пигмей не поднимается, как правило, выше 200 м), а в дневное время опускается, как предполагают, на глубину порядка 1000—1500 м. Самцы этого вида созревают при длине 17—19 см и достигают длины 22 см, самки немногим крупнее — самая большая из них имела размеры 27 см. Размножается путем яйцеживорождения, принося на свет 8 детенышей длиной около 6 см. Характерной особенностью акулы-карлика является способность к произвольному свечению. При возбуждении вся брюшная поверхность ее тела и нижние края боков светятся ровным бледно-зеленым светом, ярко вспыхивая при резких движениях рыбы и затухая, когда она успокаивается. Органами свечения служат мелкие фотофоры диаметром 0,3—0,8 мм, густо усеивающие всю нижнюю часть туловища. Люминесценция может быть очень интенсивной: известен случай, когда такую акулу заметили ночью на расстоянии 15 м от борта.
Близко к поверхности океана изредка встречается ночью и сигарная акула Isistius brasiliensis, широко распространенная в тропиках, но она, как правило, не поднимается выше верхней границы основного термоклина. Впечатляющий размах ее вертикальных миграций (с 3500 до 85 м) облегчается практически нейтральной плавучестью тела, обеспечиваемой большим количеством жидкого жира в полости тела и огромной печени. Эта акула, достигающая 50 см в длину (самцы несколько мельче — до 40 см), — прекрасный пловец. Наряду с потреблением в пищу относительно небольших рыб и головоногих она, как и второй вид рода — большезубая акула I. plutodus, способна к чрезвычайно своеобразному способу питания, который, по сути дела, может быть определен как факультативный паразитизм (уникальное явление в классе хрящевых рыб). Оба вида могут нападать на самых крупных рыб и китообразных. В этом случае акулы сначала присасываются толстыми губами к боку своей жертвы и вонзают в нее свои непропорционально большие нижнечелюстные зубы (рис. 26), имеющие форму треугольных лезвий и образующие в своей совокупности подобие острой пилы. Затем они производят вращательные движения, позволяющие выпилить из кожи и мышц жертвы конический или цилиндрический кусок, который извлекается из раны шиловидными верхнечелюстными зубами и препровождается в широкую глотку. Следы таких атак — характерные кратеровидные шрамы — отмечаются не так уж редко. Они были обнаружены на телах марлинов, тунцов, ваху, корифен, большеротой акулы, дельфинов и китов. Интересно, что такие же отметины были обнаружены при осмотре внешнего резинового корпуса американских атомных подводных лодок. Оба вида обладают способностью к люминесценции.
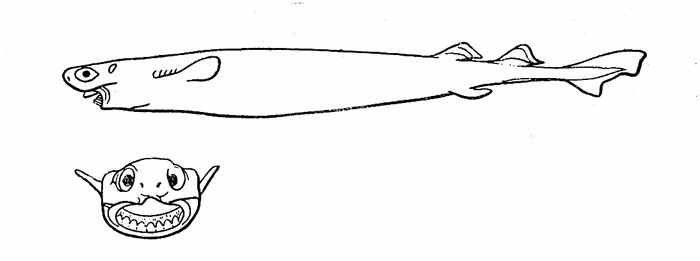
Рис. 26. Большезубая акула — факультативный паразит tspyniibix рыб и китообразных.
После лова в мезопелагиали сети и тралы иной раз приходят на борт с характерными дырами, происхождение которых долго оставалось непонятным — ведь в толще воды нет, как известно, предметов, за которые можно зацепиться. Оказалось, что эти разрезы - своеобразные автографы сигарных акул, побывавших в трале и благополучно ушедших на свободу. Лишь в крайне редких случаях, видимо, будучи пойманными в самых верхних горизонтах, они разрешают извлечь себя из воды, но и в этом случае чаще всего не ждут, пока их освободят из сети, а сами прорезают ее острыми, как бритвы, зубами, снова возвращаясь в море или, гораздо реже, падая па палубу.
Живые акулы (брать их в руки, как показывает мой опыт, надо с большой осторожностью), помещенные в аквариум, интенсивно светятся в темноте таинственным зеленоватым светом, и я видел эту иллюминацию несколько раз. Впервые такое свечение наблюдалось в начале прошлого века натуралистом старого закала Ф. Беппеттом, который оставил его прекрасное описание, приводимое здесь с некоторыми купюрами. «В темном помещении экземпляр представлял собой необычное зрелище, Вся нижняя поверхность тела и головы ярко излучала зеленоватый фосфоресцирующий свет, что поистине придавало животному страшный и пугающий облик. Люминесценция была постоянной и заметно не усиливалась при раздражении... Когда акула погибла (после трех часов наблюдений) люминесценция полностью исчезла... Моим первым впечатлением было, что рыба случайно получила некоторое количество светящегося вещества из моря, но более тщательное изучение не подтвердило этого подозрения, потому что постоянство, с которым свечение занимало определенные части тела и плавников, его неизменность при жизни и прекращение после смерти не оставляют сомнения в том, что это был жизненно необходимый процесс...»
Самыми крупными мезопелагическими хищниками являются, по-видимому, гемпиловые рыбы — змеиная макрель максимальной длиной около 1 м (как интерзональ-пый мигрант, она была рассмотрена в предыдущей главе), эсколяр (Lepidocybium flavobrunneum) длиной до 2 м (обычно 1—1,2 м), достигающая иногда еще больших размеров руветта (Ruvettus pretiosus). Оба последних вида более обычны, впрочем, над материковым склоном и подводными поднятиями и, может быть, должны считаться бенто-пелагическими. В своем вертикальном распространении они тяготеют к верхней части основного термоклина. Истинно мезопелагические темпилиды из родов Diplospinus, Paradiplospinus и Nealotus не превышают 25—45 см в длину.
В целом рыбы, входящие в состав глубоководных пелагических сообществ, характеризуются значительно меньшими размерами, чем рыбы приповерхностных слоев. Как показывают ориентировочные расчеты (в качестве примера проанализированы данные по максимальной длине приблизительно 400 видов, известных, согласно новейшей сводке, из вод Южной Африки), длина «среднестатического» глубоководного вида равна примерно 12 см, что на целый порядок меньше длипы «среднего» эпипелагического вида, которая достигает 102 см (рис. 27). При этом наибольшая длина двух третей (64%) мезобатипелагических видов не превышает 20 см, а преобладающей их части (85%) —50 см, тогда как абсолютное большинство (93%) эпипелагических видов может достигать Длины более 20 см, а почти три четверти (72%) — и более 50 см.
Неудивительно поэтому, что подавляющее большинство глубинных видов относится к пассивной жизненной форме (т. е. к планктону в широком понимании термина). К ней принадлежат и многие из крупных рыб (например, полутораметровый удильщик церация). Настоящие нек-тонные виды характерны только для самых верхних горизонтов мезопелагиали, а в холодных глубинах с ничтожными кормовыми ресурсами отсутствуют даже микронектонные рыбы: по-видимому, активный поиск и преследование добычи энергетически не оправдывают себя в этих специфических условиях.
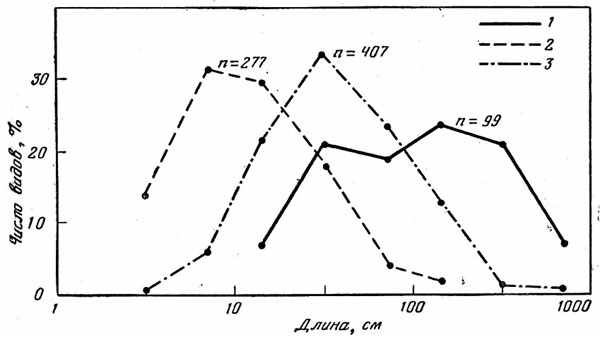
Рис. 27. Распределение видов эпипелагических (J), глубоководных пелагических (3) и глубоководных донно-придонных (3) рыб по максимальной длине особей. Среднегеометрическая длина вида составляет 102 см в эпипелагиали, 12 см в глубинной толще воды и 37 см у дна и над ним.
Рассматриваемые в этой главе глубоководные пелагические рыбы входят в состав сообществ четырех крупномасштабных биотопов — мезо-, бати-, абиссо- и хадопелагиали, однако комплексы видов, населяющие эти биотопы, далеко не равнозначны по объему и разнообразию (табл. 3). Наиболее богатый видами (около 950) мезопелагический ихтиоцен достаточно хорошо обособлен от батипелагического (в нем около 200 видов), но с дальнейшим увеличением глубины происходит лишь общее обеднение фауны. Специфических абиссопелагических и тем более хадопелагических рыб, насколько можно судить по имеющимся сейчас данным, не существует, и поэтому ихтиоцены трех нижних зон водной толщи можно рассматривать совместно. Общее число видов в глубоководной, по-видимому, близко к 1000 (все цифры в табл. 3 следует рассматривать впрочем, как приблизительные).
Мезопелагический биотоп занимает, как уже указывалось, воды основного пикноклина, как правило, совпадающего с основным термоклином. Положение и четкость его верхней границы определяются в первую очередь температурным фактором. Именно поэтому в высоких широтах, где в зимне-весеннее время температура поверхностных и промежуточных вод становится практически одинаковой, нередко наблюдаются выходы глубоководных рыб даже к самой поверхности. В тропиках граница между мезо-и эпипелагиалью полностью проницаема только для никтоэпипелагических мигрантов. Эта граница, или, точнее говоря, пограничная зона, приуроченная к наиболее градиентным по температуре слоям и имеющая толщину порядка 100—150 м, должна рассматриваться как экотон — зона смешения видов из обоих контактирующих сообществ. Одни из этих видов проводят в нем определенную часть жизненного цикла, другие встречаются там постоянно или почти постоянно.
Для эпимезопелагического экотона особенно характерны крупные нектопланктонные и нектонные рыбы, находящие в этой зоне оптимальные условия существования. Некоторые из них — алепизавры, ряд видов веретенников и гемпилид — более тяготеют к верхней мезопелагиали, другие — упоминавшиеся в предыдущей главе сельдяные короли и их родичи, морские лещи, центролофы — к нижней эпипелагиали, но это разделение представляется весьма условным. Здесь же нагуливается во взрослом состоянии большинство крупных тунцов, мечерылых и акул, служащих объектами ярусного рыболовства. Основной источник пищи всех таких хищников — макропланктонные организмы (рыбы, кальмары, ракообразные), мигрирующие к верхней границе термоклипа ночью, однако те хищники, которые питаются преимущественно на утренних и вечерних зорях (алепизавры, тунцы и др.), могут использовать этот источник пищи только опосредованно — через постоянно живущих в мезопелагиали менее крупных хищных нектеров (это паралепидиды, гемпилпды и хиазмодонтиды из рыб и, возможно, некоторые кальмары).
Преобладающие в мезопелагиали макропланктонные и микронектонпые рыбы делятся па две основные экологические группы. Одну нз них (большую) образуют виды, совершающие регулярные суточные вертикальные миграции с ночным подъемом и дневным опусканием, другую — виды, сохраняющие во взрослом состоянии более или менее постоянный горизонт обитания.
Следует отметить, однако, что большинству представителей обеих групп (а также рыбам бати- и абиссопелагиа-ли) свойственны онтогенетические миграции: их икринки, личинки и мальки проходят развитие в слоях воды, расположенных ближе к поверхности, чем горизонты обитания подрастающих и взрослых особей. Проявлением одномоментного состояния онтогенетической миграции многих видов служит прямая связь размера рыб с глубиной обитания — в пределах вертикального диапазона, характерного для данного вида, более крупные особи распределены на большей глубине, чем мелкие. По мнению М. Е. Виноградова, основной смысл онтогенетических перемещений, которые хорошо выражены и у многих беспозвоночных, состоит в том, что растущие личинки и молодь могут откармливаться в более кормных верхних слоях воды.
Суточные миграции рыб (как и других пелагических животных) в самой общей форме обычно объясняют с трофических позиций, указывая, что вертикальные мигранты, поднимаясь в ночное время на меньшие глубины (вплоть до поверхности океана), используют в качестве пищи относительно более богатые ресурсы мезоплапктона этой зоны или поедают животных, перемещающихся вместе с ними. Такое объяснение ничего не дает, впрочем, для понимания обратного процесса — перемещения мигрантов на глубину после откорма. Для обоснования этого опускания было предложено несколько гипотез.
Так, в 50-е годы советский ихтиолог Б. П. Маитейфель, который был в пашей стране одним из основоположников изучения поведения рыб, высказал предположение, что сельди (а также другие рыбы из числа зрительных планктофагов) активно уходят вниз, спасаясь от «пресса хищников» поверхностного слоя, и придавал этому уходу прямое адаптивное значение. Такое объяснение не может быть принято: как ни трудно количественно оценить упомянутый «пресс», он выражен в мезопелагиали никак не меньше, чем в эпипелагиали (это особенно очевидно в тепловодной зоне, где наблюдаются самые масштабные вертикальные миграции миктофид и других макропланктонных рыб). Другая теория, также исходящая из адаптивности миграций, предполагает активное опускание вниз с целью уменьшения энергетических затрат мигрантов в том случае, если они питаются в более теплой воде, чем та, в которой происходит переваривание их пищи. Эта гипотеза, приложимая в отличие от предыдущей только к тропическим водам, не проверялась балансовыми расчетами и вряд ли выдержит такую проверку в подавляющем большинстве случаев.
По мнению Ю. А. Рудякова, ритмические вертикальные перемещения пелагических животных могут быть следствием изменения их двигательной активности в течение суток, причем опускание на глубину происходит в те часы, когда уровень этой активности оказывается недостаточным для компенсации пассивного погружения под действием силы тяжести. Для вертикально мигрирующих рыб наиболее правдоподобно выглядит следующая схема суточных перемещений: активный подъем вечером с целью встречи с главными кормовыми объектами, ночной откорм в слое обитания этих объектов (иногда с перерывом в полуночные часы), более или менее пассивное утреннее опускание на глубину (возможно, соответствующую переходу мигрантов в состояние нейтральной плавучести) и дневной пассивный отдых («сон») на этой глубине. В связи с этим нужно отметить, что большая часть мигрирующих рыб имеет плавательный пузырь — орган, довольно эффективно, хотя и не быстро регулирующий изменение удельной массы тела. У многих рыб вертикальные миграции начинают формироваться еще на личиночной стадии развития. Известно, например, что личинки многих массовых миктофид встречаются у поверхности воды только днем (это пример так называемой инвертированной, или обращенной, миграции), что прямо связывается с существованием у них суточного хода двигательной активности, обусловленной ритмом питания.
Общепризнано, что миграции контролируются суточными изменениями освещенности. Этот фактор может действовать как ориентирующий, сигнальный, определяющий ритм питания или синхронизирующий эндогенные (циркадные) ритмы отдельных особей.
Размах вертикальных перемещений может сильно варьировать. Наибольшей протяженности — с глубины 400— 800 (1000) м до поверхности — они достигают у никтоэпипелагических рыб, уже упоминавшихся выше. К ним относятся около 40 видов светящихся анчоусов из 5 родов, акула-карлик, змеиная макрель, которые, по-видимому, поднимаются к поверхности каждую ночь, а также отдельные виды винцигуерий и астронестов (например, Aslronesthes aiger), у которых такал миграция не имеет массового характера, и значительная часть популяции остается ночью в подповерхностных слоях,
У преобладающего большинства тропических мезопе-лагических рыб вертикальные миграции полностью укладываются в пределы своего биотопа — основного термоклина (т. е. между нижней границей поверхностного изотермического слоя и верхней границей глубинных однородных вод). При этом положение верхней границы ночного подъема разных видов довольно сильно варьирует: по-видимому, оно контролируется в основном температурой воды (может быть, также положением слоя концентрации пищевых объектов). Известную роль в размахе вертикальной миграции играет и освещенность (многие миктофиды, например, поднимаются в новолуние на 30— 50 м выше, чем в полнолуние). Имеются различия и в батиметрическом распределении разных размерно-возрастных групп одного вида, У некоторых видов (из родов Diaphus, Ceratoscopelus и др.) в нижние слои эпипелагиали проникают, как правило, только неполовозрелые особи.
Мигрирующие по вертикали рыбы перемещаются в толще воды в многовидовых ассоциациях — звукорассеивающих слоях, образующих особые сообщества неполного состава (без автотрофного звена), которые можно называть схизоценами. Эти слои объединяют рыб, креветок, эвфаузиид, кальмаров, киленогих и крылоногих моллюсков, сифонофор, сальп и других животных. Суточные миграции ЗРС (как и другие виды вертикальных перемещений) связывают мезопелагических консументов с пищевыми ресурсами вышележащих горизонтов и имеют существенное значение в перераспределении органического вещества в океане. Эти слои имеют очень широкое распространение и наблюдаются почти повсеместно. В продуктивных районах часто формируются сразу несколько звукорассеивающих слоев, перемещающихся по-разному, хотя иногда и сливающихся вместе ночью или днем. Основную роль в мигрирующих ЗРС играют макроплаыктонные рыбы-планктофаги, среди которых резко преобладают миктофиды из родов Diaphus, Ceratoscopelus, Lampanyctus, Hygophum и др., составляющие до 80—90% общей численности рыб в схизоцене. В некоторых участках океана к доминирующим формам относятся также винцигуерии и батилагиды. Хищные рыбы представлены в сообществах ЗРС хаулиодами, стомиями, гоностомами, астронестами, идиакантами и другими относительно крупными видами отряда Stomiiformes, питающимися только в пределах самого слоя. Глубины дневного и ночного положения звуко-рассеивающих слоев колеблются в довольно значительных пределах — у светящихся анчоусов, например, амплитуда суточных перемещений варьирует от 200 м (Diaphus regani) до 700—800 м (Ceratoscopelus warming!),— но почти всегда остаются в границах мезопелагиали, т. е. зоны, занятой промежуточными водами. В светлое время суток мигрирующие рыбы живут на глубине 300—1000 м (обычно глубже 400—500 м), а ночью концентрируются в слое от 50-100 до 300 м.
В мезопелагиали, как уже отмечалось, имеются и слабо мигрирующие рыбы, у которых размах вертикальных перемещений не превышает 50—100 м, т. е. настолько ничтожен, что им можно пренебречь (у некоторых, например у топориков рода Argyropelecus, ночью поднимается только верхняя граница). Есть там и рыбы, вовсе не мигрирующие на протяжении суток. Глубина обитания этих постоянно живущих на одном горизонте видов (их еще называют «стационарными») также не одинакова. Грубо говоря, на средних глубинах можно выделить два комплекса видов, разделение которых особенно четко выражено в тропических водах с развитым главным термоклином. Там, на глубине 400—500 м, температурный градиент сильно уменьшается и понижение температуры становится гораздо более медленным, чем в самом слое скачка, и это приводит к обособлению (конечно, не абсолютному) двух среднеглубинных группировок — верхне- и нижнемезопе-лагической. К первой относятся, в частности, рыбы-топорики рода Argyropelecus, эверманнеллиды, жемчужноглаз-ки, светящиеся живоглоты и др. Многие из них, можно повторить еще раз, входят в число основных кормовых объектов тунцов рода Thunnus и других крупных хищников. В нижней мезопелагиали постоянно живут «светлые» Циклотоны (например, Cyclothone alba), представители ро-Дов Melamphaes, Valenciennellus, Sternoptyx и др., и там же сосредоточиваются днем все рыбы, предпринимающие вертикальные миграции. На глубине порядка 300—400 м находится и нижняя граница ночного подъема наиболее слабо мигрирующих ЗРС. В результате верхние горизонты мезопелагиали — выше 300—500 м — в светлое время имеют очень скудное макропланктонное население, а макро-планктонные рыбы (если не считать личиночно-мальковых стадий развития) в этом слое днем отсутствуют.
Мезопелагический ихтиоцен, как уже говорилось, исключительно разнообразен по видовому составу. Он содержит около 960 видов, из которых две трети специфичны для среднеглубинных горизонтов. Наибольшее число мезопелагических видов содержат семейства Myctophidae (почти 200) и Melanostomiidae (примерно 180), а также Paralepididae (около 50), Melamphaidae (около 35), Astro-nesthidae (около 35), Gonostomatidae (около 30) и др. Всего в этой зоне океанской пелагиали представлено более 75 семейств, из которых около 20 являются голомезопелагическими, т. е. не представлены ни в прибрежной зоне, ни в других биотопах океанской пелагиали.
Видовое разнообразие батиабиссопелагического ихтио-цена значительно меньше, чем мезопелагического. Здесь отмечено, впрочем, не так уж мало видов — почти 200, которые, как уже указывалось, все относятся к макропланктону. Для этой зоны особенно характерны рыбы-удильщики подотряда Ceratioidei (около 100 видов, т. е. половина видового состава ихтиоцена). Типичными обитателями глубинных однородных вод являются также «черные» цикло-тоны, миктофиды рода Taaningichthys, китовидки, глубоководные угреобразные.
На всех горизонтах океанской пелагиали видовой состав рыб в тропических и субтропических водах значительно разнообразнее, чем в субполярных. В антарктической ме-зопелагиали, например, обитает чуть больше десятка видов (все эндемичные), причем более чем одним видом представлено только четыре рода — Bathylagus, Protomyctophum, Electrona, Gymnoscopelus. Это различие отчасти объясняется отсутствием термосферы в высоких широтах, однако такое объяснение неприложимо к бати- и абиссопелагиали в связи с полнейшим однообразием глубинных вод. Общая причина видовой бедности всех высокоширотных пелагических ихтиоценов состоит в том, что ранний онтогенез всех глубоководных рыб проходит в верхних горизонтах пелагиали — там, где условия среды в разных широтах различаются коренным образом. Поэтому-то адаптации большинства видов к развитию икры и личинок при высокой температуре препятствуют освоению ими субполярных районов.
Количественное распределение рыб в глубоководной пелагиали зависит от продуктивности поверхностных вод, так как первичное продуцирование органического вещества идет только в эпипелагиали и количество пищи, попадающей на глубины, определяется в конечном итоге именно этим переносом. Как и в эпипелагиали, наименьшая биомасса рыб наблюдается в центральных частях субтропических круговоротов, а наибольшая — в экваториальных и умеренно теплых водах, а также в нерито-океа-ннческих участках. Иногда наблюдаются некоторые отклонения от этого правила, которые всегда объясняются подтоком глубинных вод из районов, отличающихся иной продуктивностью поверхностного планктона. Биомасса пелагических рыб в целом уменьшается с глубиной, но в тропической зоне пик концентрации макропланктона и нектона находится не в эпипелагиали, а в верхней мезопелагиали.
Количественно оценить обилие рыб в пелагиали совсем не просто, даже если ограничить эту задачу только некрупными — макропланктонными и микронектонными объектами. К этой цели ведут разные пути (ловы разноглубинными тралами, гидроакустические съемки, прямые наблюдения из подводных аппаратов, телеучеты), но в любом случае очень важно, чтобы способы оценки были единообразными во всех странах и учреждениях, так как только в этом случае обеспечивается полная сравнимость данных. Для обсуждения этой проблемы и выработки соответствующих рекомендаций Научный совет по океаническим исследованиям (сокращенно СКОР) при ЮНЕСКО образовал в 1977 г. международную рабочую группу, в состав которой входил и я. Результатом нашей деятельности были предложения по унификации сетных орудий лова и акустических методов, однако для полного внедрения подготовленных рекомендаций в широкую практику нужно еще преодолеть немало препятствий.
С участием в этой рабочей группе у меня связано курьезное воспоминание об одной поездке в США. Я прилетел в Сан-Диего, не имея представления, как добираться оттуда до небольшого городка Айдилвилл (именно там мы должны были заседать), но будучи твердо уверенным, что меня, как было обещано, встретят в аэропорту. Однако в небольшой группе ожидавших у входа в здание я не увидел никого из известных мне ихтиологов. Не было там и плакатика с моей фамилией — так коллеги обычно встречают незнакомого человека, и я с некоторым недоумением прошел в почти пустой (дело было поздним вечером) зал. Что же делать? Вернулся назад, потоптался у входа, пока не иссяк поток пассажиров. Опять ушел в зал, отыскал в справочной книге около телефона-автомата номер знакомого ученого, но, прежде чем позволить, решил еще раз показаться на входе. Теперь там стоял только один представительный мужчина с седой бородкой клинышком, и вид у него был довольно растерянный. Может быть, этот человек встречал меня? Я демонстративно встал перед ним, поймал его отрешенный взгляд... Несколько секунд мы в упор глядели друг на друга, и вдруг его глаза ожили. «Вы не доктор Парии?» — спросил он без большой, впрочем, надежды. «Да». Не могу сказать, кто из нас — Пол Смит (так его .звали) или я — был больше рад благополучному исходу дела. Потом он признался, что, действительно, был очень растерян, так как считал, что узнает русского с первого взгляда, а среди пассажиров не оказалось никого, отличающегося от обычных людей. Я не стал уточнять, какими именно Пол представлял себе наши отличительные особенности, но догадываюсь, что специфический образ «русского» сложился в его представлении не без влияния местных средств информации.
Экология рыб глубоководной пелагиали изучеыа крайне слабо. Сведения об их питании и трофических связях, о размножении и росте, об образе жизни и поведении явно недостаточны, хотя результаты отдельных исследований уже начали публиковаться в последние годы. Это дает возможность сделать некоторые предварительные обобщения.
Размножение. Икра и ранние личинки самых разных рыб глубоководной пелагиали — светящихся анчоусов, цикдотон, удильщиков — проходят развитие в поверхностных горизонтах, но размножающихся особей в эпипелагиали никогда не наблюдали. Логично заключить, что нерест происходит на глубине обычного обитания взрослых рыб, откуда оплодотворенные икринки, обладающие положительной плавучестью, быстро всплывают. Вылупившиеся личинки по мере развития и роста постепенно опускаются все глубже и глубже.
Насколько сейчас известно, почти все глубоководные пелагические рыбы относятся к порционно нерестующим. Число икринок в порции подвержено довольно сильной изменчивости. В районе Гавайских островов оно варьирует, по данным Т. Кларка, у миктофид от 30—150 (крохотный Notolychnus valdiviae длиной не более 2,5 см) до 2,6— 9,5 тыс., а у других мезопелагических рыб от 70—250 до 27—61 тыс. ооцитов (относительно крупная Gonostoma elongatum длиной до 27,5 см), причем общее число порций составляет 10—20. Антарктический батилаг (Bathylagus autarcticus) выметывает от 250 до 1300 икринок в 3—4 порциях, а североатлантический светящийся анчоус (Benthosema glaciale) — 190—470 икринок, возможно, при разовом нересте. В яичниках «светлых» циклотон насчитывали 0,1—0,9 тыс. ооцитов, «черных» циклотон — около 2— 10 тыс., угря Derichthys serpentinus — 4,1 тыс., рыб-удильщиков, как правило, 7,5—10,8 тыс., но у очень крупной (65 см) самки цератии (Ceratias holboelli) — почти 5 млн. ооцитов. Суммарная абсолютная плодовитость у рыб мезо- и батипелагиали в общем невелика и заметно уступает плодовитости эпипелагических рыб, которые, впрочем, имеют гораздо более крупные средние размеры. Относительная плодовитость (число икринок на 1 г массы тела) у глубоководных видов также достоверно меньше, чем у приповерхностных.
Размножение глубоководных пелагических рыб в тропических водах, по всей вероятности, продолжается в течение всего года, но у краев тепловодной зоны либо имеет пик в летне-осеннее время, либо вообще ограничено этим сезоном. В то же время в субполярных водах существуют как зимненерестующие, так и летненерестующие виды (в Северной Пацифике, например, к числу первых принадлежит миктофида Stenobrachius leucopsarus).
Некоторые из упоминавшихся специфических особенностей батипелагических рыб, связанные с их размножением, в частности, половой диморфизм по размерам (и крайнее его проявление — самцовый паразитизм) и по строению органов обоняния, а также синхронный гермафродитизм, обеспечивающий возможность нереста любой пары случайно встретившихся производителей, несомненно, объясняются трудностями отыскания партнеров в условиях разреженного существования.
Возраст. Данных по возрасту глубоководных рыб пока совсем немного, и почти все цифры, приводимые здесь, получены Дж. Фитчем и Р. Лавенбергом при исследовании отолитов отдельных экземпляров из вод, прилегающих к берегам Калифорнии. Продолжительность жизни этих рыб в общем невелика. Скорее всего созревают (еще до наступления годовалого возраста) и меньше всего живут (1,5—2 года) мигрирующие в эпипелагиаль тропические миктофиды и винцигуерии: до наступления половой зрелости они быстро растут, а затем почти беспрерывно нерестятся вплоть до самой смерти. Так, Benthosema pterotum, наиболее массовый вид миктофид Аравийского моря, становится половозрелой к 6—7 месяцам жизни при длине порядка 40 мм (максимальная длина этого вида 50 мм) и гибнет после окончания нерестового периода в возрасте чуть больше 1 года. Созревание слабо мигрирующих и постоянно остающихся на глубинах, а также обитающих в высоких и умеренных широтах рыб наступает позже (у батипела-гического угря Serrivomer sector, например, в возрасте около 4, у северного светящегося анчоуса Benthosema glaciale — в 2—3 года). Продолжительность их жизни составляет от 3—5 (циклотоны, рыбы-топорики, батилаги, умеренно-холодноводные и глубинные миктофиды, веретенники рода Lestidium и др.) до 6—8 лет (хаулиоды, идиакан-ты, кинжалозуб, китовидки, живоглоты) и даже 10—18 лет (представители родов Poromitra, Nemichthys, Gigantactis). Это свидетельствует о более высоком уровне энергетических процессов у мигрирующих по вертикали видов по сравнению с ограниченно мигрирующими и немигрирующими.
Питание. В каждом биотопе глубоководной пелагиали есть и планктоядные и хищные рыбы. Среди мезопелаги-ческих планктофагов доминируют, как уже отмечалось, мигрирующие по вертикали миктофиды и винцигуерии. Исследования, выполненные в последние годы Т. А. Гореловой, Т. Кларком и другими специалистами, показывают, что те из них, которые достигают ночью поверхности, питаются в основном приповерхностными беспозвоночными (копеподы понтеллиды и др.), а перемещающиеся в составе ЗРС без выхода в верхний однородный слой — мигрирующим вместе с ними интерзональным мезопланктоном (главным образом также ракообразными — копепо-дами, остракодами, амфиподами, эвфаузиидами) и являются основными потребителями этого планктона. Рыбы-мигранты питаются по преимуществу ночью или в вечерние и утренние часы (т. е. на закате и восходе солнца) и, подобно всем прочим зоофагам океанской толщи, захватывают обнаруженную ими (как правило, визуально) добычу, какой бы размер она ни имела, поодиночке (единственным фильтратором среди глубоководных рыб является, насколько сейчас известно, большеротая акула мегахазма). Об этом свидетельствует, в частности, явное преобладание в их желудках непрозрачных, более или менее контрастных по окраске объектов. Несмотря на очень широкие пищевые спектры и их большое сходство у разных видов тропических миктофид, между ними, по меньшей мере в некоторых районах, существуют определенные различия в составе пищи, которые, по всей вероятности, связаны с различиями в длине рыб (корреляция между размерами потребителя и жертвы часто выражена довольно явно) и в видовом составе планктона на горизонтах их ночного подъема. Это, по-видимому, способствует снижению межвидовой конкуренции за пищу в условиях ограниченных трофических ресурсов, поскольку у миктофид, живущих в более продуктивных по планктону умеренных и холодных водах, различий в составе пищи фактически не обнаружено. В олиготрофных участках океана миктофиды демонстрируют исключительно высокую кормовую пластичность: Ceratoscopelus warmingi, например, может использовать в пищу не только излюбленных копепод, но также хетогнат, оболочников, сифонофор, личинок рыб и даже фитопланктон.
Немигрирующие планктофаги — циклотоны, гоностомы, рыбы-топорики, меламфаи, батилаги (кроме совершающего суточные миграции Bathylagoides nigrigenys) — питаются независимо от времени суток или, что представляется характерным только для мезопелагических форм — жителей сумеречной зоны (Sternoptychidae и др.), в дневные часы. Основной пищей топориков, гоностомовых и меламфаев служат ракообразные (у более мелких — копеподы, у крупных — можно напомнить, что длина Gonosto-ma elongatum превышает 25 см,— эвфаузииды). Батилаги предпочитают планктеров с мягкими покровами — крылоногих моллюсков, личинок головоногих, аппендикулярий и самых мелких рачков. Средние индексы наполнения желудков у пелагических рыб уменьшаются с глубиной в соответствии с разреженным распределением глубинного плактона, и батипелагические циклотоны (в частности, Cyclothone pallida) влачат, казалось бы, самое жалкое существование (для них, впрочем, совершенно нормальное), поглощая в среднем только одного маленького рачка раз в 10 дней. Рассчитано, что суточные рационы немигрирующих рыб составляют примерно 1% сырой массы их тела и заметно уступают рациоцам мигрирующих видов, у которых они достигают 3—5, а у быстро растущей молоди даже 10—15% сырой массы тела.
Среди рыб, питающихся макроплаиктониыми объектами, особое место занимают специализированные потребители относительно крупных ракообразных — больших глубоководных эвфаузиид и креветок. Это все виды бекасорылых, или нитехвостых, угрей (семейство Nemichthyidae), желудки которых никогда не содержат другой пищи, а также, возможно, еще два семейства угрей — пилосошниковые (в питании Stemonidium hypomelas отмечены только ракообразные, но виды рода Serrivomer наряду с ними поедают рыб и кальмаров) и пессарамфовые (для представителей обоих родов — Nessarhamphus и Derichthys — ракообразные указываются в качестве основ-ной пищи). Предположительно такая специализация связана с какой-то особенностью поведения: можно думать, что все эти угри «знают секрет» приманивания крупных ракообразных, так как активное выборочное преследование именно этих жертв хищникам явно не по силам. Интересно, что узкоспециализированные ракоеды (карцинофаги) есть и среди стомиеобразных рыб (Tactostoma macropus, виды рода Photostomias), которые не имеют ] ничего общего с глубоководными угрями.
Обычно считается, что остальные хищные рыбы не имеют особых пищевых предпочтений, но это, по-видимому, не совсем правильно. Так, стомиеобразные хищники — хаулиоды, астронестовые, меланостомиевые и другие, входящие в сообщества ЗРС, питаются только рыбами и беспозвоночными, также мигрирующими в составе этих слоев. Мало того, недавно выяснилось (эти данные получены Т. Кларком), что хищники, обладающие длинными подбородочными усиками со светящимися органами на конце (представители родов Stomias, Eustomias, Mela-nostomias и др.) или удлиненным лучом спинного плавника, также оснащенным люминесцентной железкой у конца (виды рода Chauliodus), питаются главным образом рыбами — миктофидами и др., которые привлекаются светом этих желез. При этом в тех случаях, когда представители разных родов или разных видов одного рода (например, рода Enstomias) сильно различаются по строению светящихся приманок (рис. 28), между ними есть и более или менее существенные различия в составе кормовых объектов — у некоторых в желудках преобладают такие жертвы, доля которых в ихтиоцене, судя по траловым ловам, совсем незначительна. Преимущественными ихтиофагами являются и батиабиссопелагические Удильщики, которые также подманивают добычу светящимися эскальными приманками и в некоторых случаях светящимися придатками усиков.

Рис. 28. Концевые части подбородочных усиков у разных видов эвстомий (род Eustomias)
Виды стомиевидных, у которых усик отсутствует или развит слабо (как упомянутые выше тактостома и фотостомия), питаются только ракообразными или преимущественно ими и, по всей вероятности, активно ловят свою Добычу. В некоторых случаях хищник (например, Thysa-nactis dentex) может использовать оба способа питания — подманивать рыб с помощью светящейся «удочки» и визуально выискивать зоопланктон (включая и довольно крупных раков). Самый глубоководный представитель этой группы — Malacosteus niger, немигрирующий обитатель батипелагиали, имеет широкий спектр питания — от мелких копепод до рыб.
Немигрирующие мезопелагические хищники сильно различаются по размерам — мелкие эверманнеллиды, жемчужноглазки и светящиеся живоглоты (Pseudoscopelus) имеют всего 6—7 см в длину, тогда как крупные веретенники (Sudis hyalina), кинжалозуб, алепизавры могут достигать 1—2 м. Известно, что их пища достаточно разнообразна и состоит в основном из рыб, головоногих и крупных ракообразных, однако специальные исследования касались только питания двух видов алепизавров. Исключительно широкий кормовой спектр этих рыб имеет, пожалуй, лишь одно ограничение — в нем практически отсутствуют все организмы, ассоциированные с ЗРС, в частности основной их компонент — миктофиды. Мигрирующие формы (диафусы, винцигуерии, креветки сергестиды, кальмары кранхииды) входят вместе со стационарными обитателями нижней мезопелагиали (светлые циклотоны и др.) в рацион эверманнелловых, жемчужноглазковых и веретенниковых рыб. Это различие можно объяснить тем, что алепизавры (как и тунцы) добывают пищу на утренней и вечерней заре, когда ЗРС находятся выше горизонта их охоты, а другие хищники — преимущественно днем, т. е. в период нахождения ЗРС на глубине. Получается, таким образом, что очень мощная трофическая система мигрирующих слоев, которая сохраняет автономию по меньшей мере на трех уровнях консументов и включает преобладающую часть биомассы мелких средне-глубинных рыб (миктофиды и пр.), кормит на выходе только стационарных нижнемезопелагических хищников. Их численность не поддается пока прямой оценке (особенно численность нектонных видов — веретенников, гем-пилид и др.), но не может быть малой, что явствует также из данных о значении этих объектов в питании тунцов и других хищников, кормящих в переходной между эпи- и мезопелагиалью зоне.
Распространение. Пространственное распределение макропланктонных видов океанических рыб более или менее соответствует распределению естественных участков пелагической среды — замкнутых или частично замкнутых круговоротов течений, в которых циркулируют водные массы. Крупномасштабные круговороты (тропические, субтропические и субполярные), простирающиеся от берега до берега каждого океана, и зоны их соприкосновения населены «океаническими» (в понимании К. В. Беклемишева) видами. В участках схождения и расхождения течений у материков живут дальпенеритические виды.
К этим большим круговоротам и к среднемасштабным круговоротам, существующим в их пределах в качестве особых структурных элементов, приурочены основы видовых ареалов макропланктонных рыб, т. е. такие участки, в которых вид может существовать неопределенно долго вне зависимости от его наличия в других частях океана. Ареал, кроме его основы, включает также нестерильную и стерильную области выселения в выносящих течениях, и они могут быть очень протяженными. Правда, определить границу между ними в большинстве случаев непросто: это требует специальных исследований физиологического состояния рыб, вроде осуществленных У. О'Деем и Б. Нафпактитисом на атлантических миктофидах рода Lobianchia.
Таким образом, основой ареалов планктонных видов рыб могут служить как крупномасштабные круговороты океанских течений, так и круговороты, расположенные в нейтральных областях деформационного поля. Основа ареала многих широко распространенных рыб (в частности, видов с циркумтропическим распространением) может охватывать несколько круговоротов. Существенно отметить, что во вторичных водных массах (например, между субтропическими и субполярными круговоротами) смешанное население состоит из зависимых популяций видов, имеющих основы ареалов в соседствующих водных массах или в дальненеритических круговоротах.
Изложенные взгляды вполне соответствуют широко распространенным представлениям (так называемая «гипотеза водных масс») о том, что стациями популяций планктонных видов являются водные массы, понимаемые как совокупности однородных по своим характеристикам (температура, соленость и др.) объемов воды. В то же время надо отметить, что выделение этих однородных объемов по одним только температурно-соленостным показателям недостаточно дробно для выявления всех пелагических биотопов. Некоторые круговороты (особенно среднемасштабные), практически не различающиеся по температурно-соленостным характеристикам, служат основами ареалов разных видов, что, по-видимому, определяется их обособленностью по другим факторам среды, важным для жизни этих видов.
Рассматривая географическое распространение конкретных видов, как правило, почти всегда удается с той или иной степенью достоверности, зависящей прежде всего от изученности объекта, определить, хотя бы в предположительной форме, положение основы (или нескольких основ) их ареалов. Почти все затруднительные случаи могут быть объяснены неполнотой данных или недостаточностью сведений о жизненных циклах вида, но имеются и такие факты, которые не поддаются объяснению в рамках изложенной концепции. В качестве примера можно упомянуть, например, ареалы двух централь-нотихоокеанских миктофид — Protomyctophum beckeri (к югу от Гавайских островов — между 8—20° с. ш. и 144— 166° з. д.) и Lampanyctus hubbsi (вдоль экватора — между 10° с. ш.— 5° ю. ш. и 120—160° з. д.), в пределах которых нет, судя по имеющимся данным, никаких замкнутых горизонтальных циркуляции (возможно, правда, что основы их ареалов приурочены к вертикальным круговоротам).
Основные типы ареалов (их называют также «географическими элементами фауны») макропланктоиных рыб группируются следующим образом (рис. 29, А—М):
I. «Океанические ареалы (ареалы океанических круговоротов).
A. Широкотропический тип. Широкотропические виды живут, грубо говоря, во всей зоне между 40° с. ш. и 40° ю. ш. Основы их ареалов находятся во всех тропических и субтропических круговоротах, зависимые популяции есть во вторичных водных массах на периферии тропиков, а некоторые виды распространены и в тропических дальнеиеритических участках. В то же время следует отметить, что многие широкотропические рыбы имеют более или менее обширные разрывы в географическом распространении, отсутствуя, например, либо в ядрах центральных круговоротов, либо в переходных, либо в дальненеритических районах.
Б. Экваториально-центральный тип. У этих видов основы ареалов расположены в экваториальных и центральных водах (часто с пробелами в тех или иных их участках), но области их распространения не столь обширны, как у широкотропических рыб, так как они отсутствуют во вторичных водных массах у границ тропической зоны, а обычно и в дальненеритических районах.
B. Экваториальный тип. Основы ареалов лежат в тропических круговоротах у экватора, но отдельные виды имеют дополнительные основы в некоторых дальненеритических участках, так что области их распространения расширяются у берегов (особенно в западных частях океанов). Очень характерный подтип этого географического элемента составляют ареалы видов, отсутствующих в восточноэкваториальных дальненеритических районах.
Г. Центральный тип. Все ареалы этого типа имеют основы в олиготрофных субтропических круговоротах. Некоторые из центральных видов живут во всех пяти этих круговоротах и имеют бицентральное (антиэкваториальное) распространение, другие встречаются только в одной-двух центральных водных массах.
Д, Е. Периферический тип. Эти виды живут на периферии центральных водных масс, главным образом на севере северных и (или) на юге южных субтропических круговоротов (Д), но в относительно немногих случаях также у восточных (реже у западных) берегов материков и на экваторе (Е).
Ж. Переходный тип. Эти ареалы приурочены либо к северной части Субантарктики («зона субтропической конвергенции»), либо Северотихоокеанскому и (или) Североатлантическому течениям, т. е. ко вторичным водным массам, образованным в результате смешения субполярных и центральных вод. Основы ареалов, по-видимому, находятся в круговоротах, прилегающих к материкам. Особенно многочисленны такие рыбы в южном полушарии.
3. Нотально-южнобореальный тип. Ареалы этого типа располагаются в южных частях Субантарктики (между субтропической конвергенцией и южным полярным фронтом) или Субарктической Пацифики, где примесь центральных вод незначительна. В Атлантическом океане этот географический элемент ихтиофауны отсутствует.
И. Антарктическо - севернобореальный тип. Эти ареалы имеют своей основой в северном полушарии западные круговороты субарктических вод Тихого и Атлантического океанов, а в южном — циркуляции, образованные течениями Восточных и Западных ветров.
К. Широкий антарктическо-бореальный тип. Виды, принадлежащие к этому типу, имеют основы ареалов в субполярных круговоротах либо северного, либо южного полушария.
II. Дальненеритические ареалы (ареалы нейтральных областей).
Л. Тропический тип. Основы ареалов тропических дальиенеритических видов расположены в нейтральных областях между тропическими и субтропическими круговоротами, но видов, общих для всех этих областей, нет. Особенно много эндемичных рыб обитает в восточной тропической Пацифике.
М. Переходный тип. Эти дальнеперитические ареалы находятся в нейтральных областях между субтропическими и субполярными круговоротами.
Н. Широкий дальненеритический тип. Виды с ареалами этого типа живут в тропических и в переходных нейтральных областях.
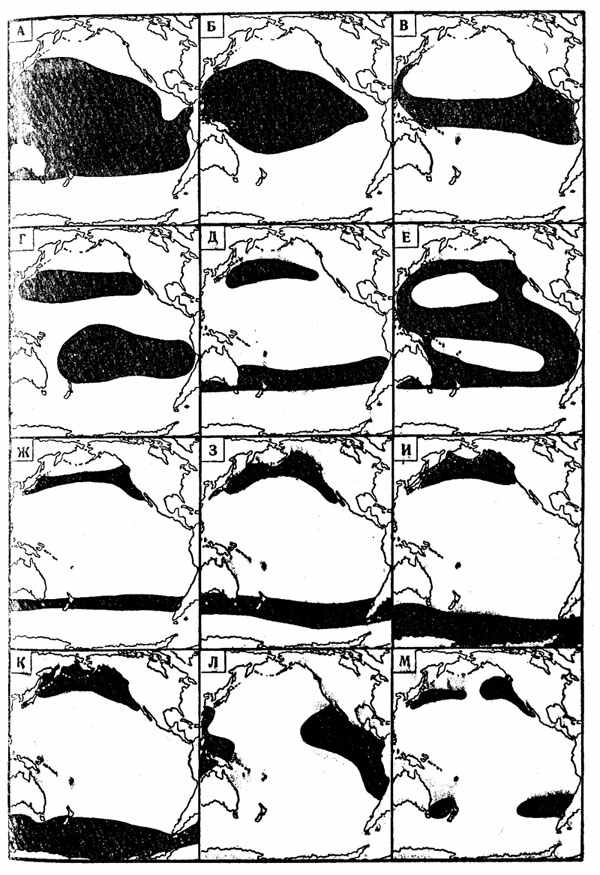
Рис. 29. Типы океанических (А - К) а дальненеритических (Л, М) ареалов макропланктонпых мезопелагических рыб (на примере тихоокеанских никтофид)
Ареалы батипелагических рыб изучены пока менее детально, чем ареалы мезопелагических видов. Некоторые из них имеют очень широкое распространение в Мировом океане. Крайний пример представляет в этом отношении одна из циклотон (Cyclothone pseudopallida), обладающая почти космополитическим ареалом (этого вида нет только в Северном Ледовитом океане, восточной тропической Пацифике и Аравийском море), но, к сожалению, положение основы ареала этого вида еще не установлено. Между тем, как показывает пример Gonostoma gracile (рис. 30), область размножения батипела-гического вида может занимать очень малую часть акватории, на которой этот вид распространен.
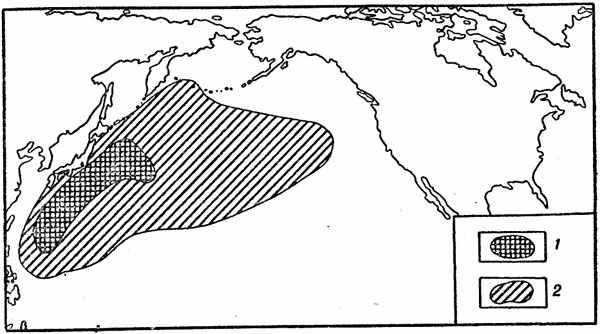
Рис. 30. Ареал северотихоокеанской гоностомы 1 — область размножения (основа ареала); 2 — область распространения взрослых особей.
Известно, что многие батипелагические рыбы имеют широкотропическое распространение. Из более ограниченных ареалов следует отметить такие географические элементы, как экваториальный, восточнотихоокеанский и вос-точноатлантический тропические, антарктическо-ноталь-ный и бореальный. Наблюдающееся сходство в типологии ареалов мезо- и батипелагических рыб объясняется, по всей вероятности, тем, как уже говорилось, что раннее развитие всех глубоководных видов, независимо от глубины, на которой они живут во взрослом состоянии, происходит в верхних горизонтах пелагиали.
Что касается нектонных глубоководно-пелагических рыб, которые, следует напомнить, характерны главным образом для переходного слоя между эпи- и мезопелагиалью и для верхних горизонтов последней, то их географическое распространение пока еще не изучено в деталях. Можно думать, что оно определяется, как и распределение эпипелагического нектона, температурой воды и кормовыми условиями. Гемпилиды и веретенник судис (Sudis hyalina) имеют циркумтропические ареалы, а алепизавры встречаются как в тропической зоне (там происходит их нерест), так и за ее пределами. Для антитропического киюкалозуба характерно разобщение репродуктивной части ареала, приуроченной к субтропическим водам, и области нагула, охватывающей субполярные районы.