
Рыбы открытого океана
| Вид материала | Документы |
- Казимир Малевич, я начало всего, 829.32kb.
- Конспект открытого урока биологии в 7 классе по теме: «класс хрящевые рыбы», 39.29kb.
- Урок-викторина по теме Животные. Класс рыбы, 75.59kb.
- Конспект открытого урока биологии в 7 классе по теме: «класс хрящевые рыбы», 48.22kb.
- Рельеф дна Мирового океана, 52.6kb.
- Урок биологии в 7 классе. Тема: «Внутреннее строение рыб», 112.94kb.
- Процессы и производственные линии замораживания продуктов растительного происхождения, 145.64kb.
- Тема урока Северный Ледовитый океан; особенности природы, хозяйственного использования,, 53.05kb.
- Магистерская специальность «Экономика ресурсов Мирового океана», 26.67kb.
- А. В. Клименко конкурсная документация по проведению открытого конкурс, 1840.42kb.
Зубатки (два рода семейства Anarhichadidao), распространенные в северных частях Атлантического и Тихого океанов, живут в основном в шельфовой зоне, но в зимнее время мигрируют на глубины (эта группа всесторонне изучена В. В. Барсуковым). Наиболее глубоководный вид - синяя зубатка (Anarhichas denticulatus), достигающая 1,4 м в длину, отмечена даже на глубине более 900 м. Все зубатки откладывают очень крупную донную икру, но их личинки поднимаются в толщу воды, где проходят весь мальковый период жизни (его продолжительность у разных видов сильно разнится).
Еще одна холодноводная группа рыб — нототениевидные, эндемичная для Антарктики и Субантарктики, где она доминирует. Эта группа отличается, как показано работами А. П. Андрияшева, X. Девитта, А. В. Балушкина, А. В. Неелова, Ж.-К. Юро и других ученых, большим систематическим и экологическим разнообразием. В сем. нототениевых (Nototheniidae), усатых дракончиков (Artedidraconidae), антарктических плосконосов (Bathydraconidae) и белокровных рыб (Channichthyidae) есть и глубоководные донно-придонные формы, обитающие на глубине 500—1500 м и более. Среди преимущественно прибрежных и шельфовых нототениид наиболее глубоководными являются антарктическая сквама (Lepidonotothen kempi) и черноротый трематом (Trematomus loennbergi), которые держатся на склоне континентальной ступени Антарктиды на глубине 500—1000 м, а также патагонский клыкач (Dissоstichus eleginoides) — очень крупная рыба (длина до 2,1 м, масса до 70 кг), добываемый у островов Южная Георгия и Кергелен на глубине до 800 м. Большинство плосконосов живет на глубине, не превышающей 500—800 м, но представители рода Bathydraco — самого глубоководного среди нототеноидных рыб — населяют большие глубины (вплоть до 2500 м). Усатые дракончики рода Pogonophryne также отмечены до глубины 2300 м. Глубинная белокровка (Chionobathyscus dewitti), у которой, как и у других ханнихтид, в крови нет ни эритроцитов, ни гемоглобина, также ловилась у берегов ледового континента на глубине до 2000 м. У всех нототениевидных икра развивается на дне а личинки ведут пелагический образ жизни.
В верхней мезобентали тепловодной зоны (на глубине до 500—600 м) вполне обычны также такие рыбы, как хампсодоны (сем. Champsodontidae), звездочеты (Uranoe scopidae: Kathetostoma, Uranoscopus и др.), перкофид (Percophidae, в том числе карликовые виды родов Osopesaron и Pteropsaron, не превышающие 4—5 см в длину) драконетки (Draconettidae).

Рис. 41. Руветта (1) а угольная рыба-сабля (2).
Для среднеглубинных горизонтов очень характерны саблеподобные рыбы (рис. 41) — два близкородственных семейства гемпиловых (Gempylidae) и волосохвостых (Trichiuridae), входящие в состав подотряда скумбриевидных. Все они имеют в той или иной степени удлиненное тело, большой рот и очень острые мощные клыкообразиые зубы, легко рассекающие добычу на части. В мезобентопелагических ихтиоценах на глубине 300—1000 м (нужно сказать, что эта группа представлена также на шельфе и в океанской пелагиали) особенно важное значение принадлежит прометихтам (род Promethichthys), рексиям (Rexea), бентодесмам (Benthodesmus) и угольным саблям (Aphanopus) — хищным рыбам, длина которых нередко достигает 1 м, а также еще более крупной руветте (Ruvettus pretiosus). Эта рыба может иметь длину 1,8 м при массе около 50 кг. Во многих районах мира она служит объектом специального лова на особые снасти, крючки которых опускаются на глубину 300—700 м. Такой промысел производится, в частности, у берегов Кубы, Мадейры, Коморского архипелага, многих островов Полинезии. Мясо руветты содержит очень много жира, который обладает слабительными свойствами (на некоторых языках ее называют «касторковой рыбой») и при приготовлении рыбы в пищу должен удаляться во время варки.
Саблеподобным рыбам я посвятил немало внимания и времени. Еще в 1955 г., в первом своем плавании в тропиках, мне удалось поймать сачком крупную змеиную макрель, в то время представлявшуюся большой редкостью. В очень популярной в те годы книге Тура Хейердала о путешествии на бальсовом плоту «Кон Тики» встрече с этим «чудищем» посвящено немало эмоциональных строк. С тех давних пор эта группа постоянно вызывала мой интерес, и в 1969 г. я предложил Владимиру Беккеру, давнему приятелю (мы вместе учились в институте) и коллеге по лаборатории, совместно разобраться в не очень большой коллекции, которой мы тогда располагали: я взялся за рыб-сабель, он — за гемпилид. В итоге этого внепланового исследования был опубликован предварительный обзор надсемейства, после чего Володя навсегда вернулся к своим любимым миктофидам, а я тоже занялся другими проблемами, хотя не терял группу из виду и между делом описал в ней еще три вида бентодесм. В середине 70-х я заинтересовал систематикой трихиурид промыслового ихтиолога из ВНИРО Сергея Михайлина, вместе с которым мы дали описания двух новых рыб-сабель, но трагическая гибель Сережи в конце 1981 г. не позволила ему до конца реализовать свои возможности (он погиб, как герой, спасая пассажиров в загоревшемся на ходу вагоне поезда Калининград - Москва). В память о друге (нас объединяло не только общее дело, но и одинаковая страсть к охоте) я назвал его именем один из новых видов угольных сабель, решив поставить на этом точку в изучении этой группы. Не получилось... При обработке коллекций с подводного хребта Наска выяснилось, что там обитают два вида рексий - оба неописанные,— а исследование сравнительных материалов из других районов Тихого и Индийского океанов (я получил их из музеев многих стран) показало, что в роде существуют по меньшей мере шесть видов (считалось, что их только два). Результаты этой работы еще не опубликованы.
Из океанических строматеевидных рыб к числу придонных принадлежат немногие. Это — гипероглифы (род Hyperoglyphe) и шедофилы (Schedophilus) из семейства центролофовых — крупные рыбы длиной до 1 м, обитающие главным образом в субтропических и умереннотепловодных районах (в том числе над подводными поднятиями) на глубине до 300 м и более. Придонно-пелагическими; являются также ариоммы (около полутора десятков видов рода Ariomma), обычные на глубине 100—600 м во всех тропических морях.
Не опускаются глубже мезобентали и преимущественно шельфовые камбалообразные рыбы. Так, наиболее глубоководпый род ботусовых камбал (сем. Bothidae) — хасканопсетты (Chascanopsetta), недавно ревизованный К. АмаЯ ока, содержит 5 видов, которые живут на глубине 250-640 м. Один из этих видов — гавайская пеликанья камбала (Ch. crumenalis) отличается крайне своеобразным обликом: нижняя челюсть у нее очень длинная (она на треть: своей длины выступает перед рылом) и поддерживает кожистую горловую сумку, очень напоминающую ту, что имеется у настоящих пеликанов. Еще глубже обитают представители эндемичного для Южного океана семейства безруких камбал (Achiropsettidae), недавно выделенного С. А. Евсеенко и характеризующегося отсутствием грудных плавников: максимальная глубина вылова неоахиропсетты (Neoachiropsetta milfordl) составляет 840 м.
Среди настоящих, или правосторонних, камбал (сем. Pleuronectidae) есть немало видов, например северные малороты (род Microstomus) или тропические пецилопсетты (Poecilopsetta), приуроченных к пограничной между шельфом и склоном зоне (глубина порядка 100— 300 м). К наиболее глубоководным видам относятся палтусы, прежде всего черный, или синекорый (Reinhardtius hippoglossoides), и белокорый (Hippoglossus hippoglossus) — живущие в северных частях Атлантического и Тихого океанов хищники, активно преследующие свою добычу в толще воды. Полноразмерные особи черного палтуса достигают 1,3 м в длину, но белокорый значительно крупнее (рекордная длина 4,7 м при массе 337 кг). Скопления всех палтусов обычно приурочены к материковому склону (глубина 150—800 м), а наибольшая глубина их обитания превышает километр (черный палтус вылавливался даже па отметке 1700 м). Палтусы принадлежат к числу долгоживущих поздносозревающих рыб (белокорый живет до 30 лет, а созревает в массе на 10—15-м году). Нерест происходит в глубоких слоях воды, личинки сначала тоже держатся далеко от поверхности, но, подрастая, перемещаются на мелководья, где переходят к донному образу жизни. Среди камбал-плевронектид известны и еще более глубоководные — мезобатибентальные виды, например бородавчатая (Clidoderma asperrimurn) и глубинная (Еmbassichlhys bathibius), вполне обычные вплоть до 1800— 2000 м от поверхности. В состав батиального ихтиоцена полноправно входят и морские языки из рода Symphurus (сем. Cynoglossidae), живущие в тропической зоне на глубине 200—1000 м. К сказанному о камбалообразных нужно еще добавить, что среди глубоководных представителей группы практически нет чистых бентофагов (и уж во всяком случае их доля значительно ниже, чем среди мелководных).
И наконец, в отряде скалозубообразных к придонным глубоководным рыбам относится в полном составе семейство холлардиевых (Triacanthodidae), включающее 20 видов, обзор которых сделан недавно Д. Тайлером. Эти небольшие (длина не более 25 см) рыбы живут в тропических водах всех океанов на глубине 50—600 м и даже глубже. Биология их изучена слабо. Судя по некоторым особенностям, это наименее продвинутая группа скалозубообразных, но отдельные виды представляются очень сильно специализированными. Так, макрорамфосоды (род Macrorhamphosodes) имеют тонкое рыло, вытянутое в длинную трубку, на конце которой находится обращенный вбок дисковидный рот. Эта рыба, как и тидемания (Tydemania navigatoris), питается чешуей других рыб.
Сравнение состава бенто-пелагических комплексов разных вертикальных зон (табл. 4) показывает, что число видов в ихтиоценах быстро убывает с глубиной (цифры этой таблицы следует рассматривать как приблизительные). В самой верхней части склона, где представлены многие семейства, типичные для сублиторали, наблюдается наибольшее разнообразие систематического состава. Нужно подчеркнуть, что в большинстве случаев речь идет не о случайных проникновениях отдельных особей тех или иных шельфовых видов в чуждый биотоп, а о постоянном их обитании в экотонных условиях у границы смежных вертикальных зон. В справедливости такого утверждения убеждает тот факт, что такие виды живут и там, где эпибентальной зоны нет вообще. Речь идет о подводных поднятиях в открытом океане, имеющих минимальные глубины порядка 200—400 м, т. е. о верхней части талассобатиали в понимании А. П. Андрияшева. Все такие поднятия (их обычно называют «банками») населены тем не менее рыбами, которые более характерны для нижней части шельфа: как правило, их легко сразу опознать по яркой окраске и совсем не «глубоководному» облику. В качестве типичных эпимезобентальных экотонных рыб можно упомянуть аулопов (род Aulopus), длиннорылых рыб-солдатов (Ostichthys), антигоний (Antigonia), большеглазых каталуф (Cookeolus), каллантиасов (Callanthias). Верхнемезопелагические виды недавно обнаружены даже в семействах щетинозубовых, или рыб-бабочек (Chaetodontidae), и губановых (Labridae), всегда считавшихся приуроченными к прибрежным сообществам, — Chaetodon guyotensis и Bodianus thoracotaeniatus описаны с подводных гор хребта Кюсю-Палау (глубина 320—340 м).
На сообществах рыб, населяющих подводные поднятия, нужно остановиться подробнее в связи с тем интересом, который они вызывают по многим причинам. Важность их изучения первым оценил американский ихтиолог К. Хаббз, который еще в 1959 г., основываясь на очень скудных фактических данных того времени (фактически на одной случайной поимке триглы на гайоте хребта Наска), сформулировал ряд принципиальных вопросов, касающихся этой проблемы. Вот эти вопросы.
Постоянен ли видовой состав населения отдельных гор Каким образом они были заселены?
Не представляют ли они опорные пункты для преодоления открытоводных барьеров, разделяющих прибрежные фауны?
Не являются ли некоторые виды, населяющие постепенно погружающиеся горы, реликтами, адаптированными к увеличенной глубине обитания?
Ведет ли изоляция гор к видообразованию?
Не существует ли на горах и банках и над ними скоплений донных или пелагических рыб и других промысловых объектов?
Карл Левитт Хаббз (1894-1979) несомненно был одним из самых выдающихся ихтиологов нашего столетия. Он отличался необычайной широтой и глубиной научных интересов и оставил после себя весьма достойное наследие - более 700 книг и статей, касающихся разнообразнейших аспектов ихтиологии и смежных отраслей знания. Он был также простым в общении и доброжелательным человеком с хорошо развитым чувством юмора.
Мне довелось встретиться с К. Хаббзом трижды (не могу запросто называть его Карлом, как при встречах: по-русски это было бы слишком фамильярным и не соответствующим тому уважению, которое я питаю к этому человеку). Знакомство состоялось на Гавайских островах в 1964 г. во время очередного Тихоокеанского научного конгресса, на который я попал с «Витязем», совершавшим рейс в центральную Пацифику. Первое общение было очень тяжелым как по причине моего несовершенного английского, так и из-за весьма далекого от классических оксфордских норм произношения К. Хаббза, по я мог с удивлением убедиться, что даже в очень узкой специальной области (речь шла о летучих рыбах Тихого океана) он чувствовал себя совершенно свободно. На следующий день после моего доклада он шутливо поздравил меня с победой в противоборстве «с этим варварским английским», а через пару месяцев прислал в Москву вырезку из какого-то комикса, в котором говорилось о «летучих коровах», выдуманных неким впоследствии разоблаченным лжецом, написав на ней: «А существуют ли все же летучие рыбы?» В 1968 г. «Академик Курчатов» зашел в Виллемстад (о-в Кюрасао), когда там происходила международная конференция по Карибскому морю, в которой, оказалось, принимал участие и К. Хаббз. Он был очень подвижек и каждый год предпринимал дальние вояжи вместе со своей верной женой и помощницей Лорой Хаббз. На конференции он заметил меня первым (я, надо сказать, совсем не ожидал этой встречи), сразу узнал и с такой непринужденностью «проехался» насчет моей изменившейся внешности (я стал носить бороду), что два молодых ихтиолога с Барбадоса, с которыми я беседовал, с полной уверенностью сочли его моим бывшим профессором. Последний раз я видел Хаббза в 1979 г. в Калифорнии. Он был болен и знал, что его дни сочтены, сильно ослабел и постарел, но продолжал трудиться, интересовался работой коллег, говорил о желании принять участие в следующем Тихоокеанском конгрессе в Хабаровске... Прах Карла Хаббза по его завещанию был развеян над Тихим океаном. Большой фотопортрет с надписью «...советскому коллеге от старого моряка», запечатлевший его стоящим в полный рост на корме штормующего в океане судна, напоминает мне о наших встречах.
Изучение рыб подводных поднятий только начинается, но уже сейчас можно отметить, что в разных районах Мирового океана в заселении этого биотопа приняли участие в основном представители одних и тех же, притом сравнительно немногих, семейств. Как показывает сравнение ихтиофаунистических списков подводных гор Метеор и Жозефина в северо-восточной Атлантике, банки Вальдивия Китового хребта в юго-восточной Атлантике, тихоокеанских подводных хребтов Кюсю-Палау, Гавайского, Северо-Западного, Наска и Сала-и-Гомес, сходство в составе населения этих далеко отстоящих друг от друга участков довольно велико, особенно на уровне семейств и родов.
Видовой состав ихтиоценов на подводных поднятиях, определяется в первую очередь глубиной над их вершинами, а разнообразие этого состава — действием «островного эффекта», прямо зависящего от площади поднятия Талассоэпибентальные ихтиоцены на вершинах самых мелководных банок (например, гор Келсо, Кейпл и Уанганелла в Тасмановом море, имеющих глубину 40—100 м) обычно представляют собой в той или иной степени обедненный вариант сообществ ближайших шельфовых районов. Не исключена возможность, что на крохотных вершинных площадках постоянные самовоспроизводящиеся популяции эпибентальных рыб вообще отсутствуют и их население пополняется только за счет оседания пелагических личинок, принесенных извне, т. е. с ближайшего шельфа.
Существование независимых горных популяций рыб эпимезо- и мезобентопелагического комплекса, напротив, не вызывает никаких сомнений. Об этом свидетельствуют не только многолетние наблюдения над локальными популяциями таких, к примеру, видов, как рыба-кабан Pseudopentaceros wheeleri на Гавайском подводном хребте, капродон Caprodon longimanus на уже упоминавшейся банке Уанганелла, безпузырный окунь Helicolenus lengerichi и зеркальный солнечник Zenopsis sp. (неописанный вид, близкий к Z. nebulosus) на подводном хребте Наска, но и существование эндемичных видов, ареалы которых ограничены одним поднятием или группой близкорасположенных поднятий. Эндемичные талассобатиальные виды известны сейчас из многих районов Мирового океана, в том числе с гор Азорского архипелага и с Китового хребта в Атлантическом океане, Мадагаскарского и Восточно-Индийского хребтов в Индийском океане, хребтов Кюсю-Палау, Северо-Западного, Наска и Сала-и-Гомес в Тихом океане. Последний район — хребты тепловодной юго-восточной Пацифики — представляется, кстати сказать, прекрасным местом для разнопланового модельного изучения талассных ихтиоценов, в том числе и для ответа на вопросы, поставленные К. Хаббзом. Уникальность этих хребтов состоит в том, что, несмотря на близость к материку Южной Америки, они заселены рыбами индовестпацифического происхождения, и поэтому ближайшие шельфы и склоны явно не поставляют никакого материала для пополнения их населения. Сами эти горы представляют собой погруженные коралловые острова (гайоты), различающиеся по глубине и возрасту (геологическому времени существования).
Весной 1980 г. в нашу лабораторию поступили материалы, несколькими месяцами раньше собранные моими бывшими аспирантами Г. А. Голованем (теперь он возглавляет научные работы в Севастопольском бюро по подводным исследованиям) и Н. П. Пахоруковым. Они принимали участие в рейсе «Ихтиандра» на подводные хребты Наска и Сала-и-Гомес, которые до того были совсем неизученными в ихтиологическом отношении (правда, еще раньше там работали сотрудники ТИНРО, но, к большому сожалению, они, как и во многих других рыбохозяйственных экспедициях, не собирали представительных коллекций рыб). Предварительная обработка полученных нами проб (ее проводили Ю. Сазонов, Ю. Щербачев и я) дала на редкость интересные и, я бы сказал, неожиданные результаты. Выяснилось, что среди донных и придонных рыб этого района восточной Пацифики преобладают виды, которые были известны только из западной и центральное частей океана — от Японии, Австралии, Гавайских островов, Новой Каледонии, а многие другие относятся к числу неописанных; Впоследствии состоялось еще несколько экспедиций в этот рая он - я сам побывал там дважды на «Профессоре Месяцеве» и «Профессоре Штокмаые» и в результате почти полностью переключился на изучение «горных» рыб. Сейчас список видов, обнаружейных на обоих хребтах, увеличился до 160, и почти треть из них — это исключительно высокий процент — оказалась эндемичной (описания 30 новых видов уже опубликованы, остальные готовятся к печати). В моих ближайших планах одно из первых мест занимает сейчас подготовка монографии «Рыбы подводных хребтов Наска и Сала-и-Гомес», над которой мы работаем вмести с сотрудником ВНИРО А. Н. Котляром (у него я тоже был когда-то руководителем по кандидатской диссертации), надеясь завершить ее через год — другой.
И еще: не могу не упомянуть о трудностях тралений н гоpax. Представление об этих трудностях легко можно получить вообразив, что нужно провести трал с дирижабля, летящего выше сплошного облачного покрова, по склонам, скажем, Кавказского хребта. Эта мысль постоянно приходит мне в голову, когда я смотрю на горы в иллюминатор самолета, и такое действие представляется просто невероятным, но тем не менее... При первых в моей практике работах на подводных поднятиях (это был горный массив Экватор в западной части Индийского океана) мы раз за разом опускали с «Рифта» оттертрал на склоны (естественно, не вслепую, а ориентируясь по эхолоту), порвали все, что рвется но так и не смогли получить полноценной пробы. Особенно запомнился один задев, когда траловая доска, попавшая в расщелину, превратилась у нас в мертвый якорь и вышедшее на верти кальный трос (моряки говорят «на панер») и содрогающееся oт напряжения судно не могло на любом курсе сдвинуться с местаЛ пока 25-миллиметровый стальной ваер не лопнул с таким громоподобным звуком, что создалось полное впечатление взрыва. Хорошо работать только на ровных вершинах гайотов, да и там трал могут подстерегать разнообразные и неожиданные сюрпризы Среди рыб, составляющих постоянное население вершин подводных гор, есть немало чисто донных видов (в том числе зеленоглазки, хаунаксы, скорпениды, морские петухи, камбалы), но наиболее массовый и характерпый компонент сообщества составляют бентопелагические объекты — колючие акулы, мавролик, специфичные для гор рыбы-топорики и светящиеся анчоусы, рыбы-бекасы мориды, долгохвосты, солнечники, бериксы, большеголовы, некоторые ставридовые, каменные окуни, эпигонусы, красноглазки, рыбы-сабли, масляные рыбы и др.
Биология большинства «горных» бентопелагических рыб пока изучена слабо. Известно, правда, что многие из них совершают суточные вертикальные миграции: днем они держатся вблизи дна, а в темное время поднимаются в верхнюю толщу вод, удаляясь на 100—200 м и более от твердой поверхности. Как уже указывалось, такие миграции предпринимают даже мелкие стайные виды (Maurolicus muelleri, Diaphus suborbitalis). В связи с этим возникает естественный вопрос, как удается этим рыбам, несмотря на наличие течений, иногда достаточно сильных, всю ночь оставаться над поднятием, с которым они теряют визуальный контакт, и точно опускаться на него на рассвете. Все объяснения этого феномена (ориентация по светилам, поляризованному свету, градиентам скорости и т. п.) чисто умозрительны и выглядят неубедительно.
Возможность массовых миграций талассобатиальных рыб от одного подводного поднятия к другому кажется мне весьма проблематичной, если не говорить о горах, расположенных на одном цоколе: в этом случае эврибатные виды (низкотелый берикс, например, обитает в диапазоне от 50 до 1000 м), естественно, могут перемещаться в пределах всего района, подходящего по глубине. Эта точка зрения не является, однако, общепринятой. Предполагают, в частности, что рыба-кабан Гавайского подводного хребта проводит от 4—5 до 7 лет в пелагиали северной Пацифики и только после этого приходит на горы, где размножается. Весьма логичная схема функциональной структуры попу-ляционных ареалов берикса в Атлантическом океане, в основу которой положены данные о размерно-возрастном составе и зрелости рыб в разных районах, была недавно разработана Ф. Е. Алексеевым с соавторами. Вот как описывается, например, цикл существования североатлантической популяции: нерестовая часть ареала приурочена к горам Углового поднятия в западной части круговорота; оттуда пелагические икра, личинки и мальки дрейфуют в Северо-Атлантическом течении на восток и юго-восток; молодь оседает на подводных поднятиях в восточной части круговорота, а в отдельные годы и на склоне северозападной Африки; по мере полового созревания берикс активно мигрирует на запад по южной периферии круго-ворота, и половозрелый контингент постоянно обитает в Репродуктивной области (т. е. на Угловом поднятии). Я думаю, что региональные различия в размерах берикса, взятого из промысловых уловов, могут объясняться разное глубиной лова (половозрелые особи обитают глубже, чем созревающие, а траловые участки на Угловом поднятии находятся на большей глубине, чем на Азорских банках) Большое сомнение вызывает и сама способность мигрантов к отысканию нерестовых участков — для этого нужно предположить почти сверхъестественные навигационные cпoсобности рыб. И наконец, этому представлению, как и всем основанным на косвенных доказательствах соображениям в пользу существования таких миграций, противоречит полное отсутствие этих рыб над большими глубинами в открытом океане. До сих пор не отмечено ни одной; поимки взрослых бериксов, красноглазок и подобных им объектов в далеком удалении от материковых склонов подводных возвышенностей.
Хуже всего изучены жизненные циклы бентопелагических рыб и особенно их размножение и ранние стадии онтогенеза, во многом загадочные в связи с исключительно редкой встречаемостью личинок и мальков в уловах, полученных любыми орудиями лова. Относительно воспроизводства рыб, обитающих на подводных поднятиях, высказывались два предположения. Согласно одной точке зрения, развитие их планктонных стадий происходит системах местных циркуляции вблизи гор. Альтернативная гипотеза о заселении поднятий личинками и мальками, приносимыми течениями «извне», также имеет своих сторонников.
Действительно, слабая изученность процессов воспроизводства талассобентальных рыб сильно препятствует однозначному решению вопроса. В то же время доказанпость существования независимых популяций на отдельных банках, например на горе Метеор в Атлантическом; океане или на некоторых группах близкорасположенных поднятий хребтов Наска и Сала-и-Гомес, дает полное ocнование отказаться от представления об основополагающей роли разноса икры, личинок и мальков течениями и наличии значительного обмена генофондами. Развития планктонных стадий может, по-видимому, происходите в непосредственной близости от подводных возвышенностей в локальных круговоротах любого пространственного масштаба, даже не имеющих стационарного характера, но обладающих такой временной устойчивостью, которая была бы соизмерима с продолжительностью пассивного пребывания икры и личинок в пелагиали (около 10—15 суток). Еще одна гипотетическая возможность, требующая экспериментальной проверки, состоит в откладывании икринок с отрицательной плавучестью в щели и углубления дна. Более того, как показало исследование разработанной В. Б. Цейтлиным и Ю. А. Рудяковым имитационной модели независимой популяции рыб, населяющих подводную гору, такая популяция вполне способна сохраняться и воспроизводиться даже и при разносе рыбной молоди на планктонных стадиях развития течениями, если благодаря турбулентной диффузии она будет иметь шанс случайно вернуться к месту рождения. Из этой модели следует, правда, что вниз по течению от горы должен вытягиваться довольно значительный «шлейф» икры и личинок, чего в природе никогда не наблюдали.
Итак, что же прояснилось в наших знаниях о рыбах подводных поднятий за три десятка лет, прошедших с того времени, когда Карл Хаббз задал свои вопросы? Стало ясно, что видовой состав ихтиоценов отдельных поднятий характеризуется значительным постоянством во времени, ибо каждая гора представляет собой особый биотоп с населяющим его биоценозом. Заселение гор происходило точно тем же путем, что и заселение островов и их цоколей — с шельфов и склонов материков, и поэтому подводные поднятия должны рассматриваться в том же ряду природных объектов, что и океанические острова. Колонизация островных биотопов в открытом океане (включая горы) шла в основном за счет более разнообразных прибрежных фаун, населяющих западные части океанов, и чем дальше расположено поднятие от главного источника своего формирования, тем выше эндемизм его населения (максимальная доля эндемиков — около 40% в диапазоне глубин 200—500 м выявлена на хребтах Наска и Сала-и-Гомес). Поднятия, конечно же, являются «опорными пунктами» в расселении тех рыб, которые способны к достаточно долгому выживанию в пелагиали во взрослом состоянии или на стадии молоди (хороший пример дают поимки в пелагиали восточной тропической Пацифики крупных мальков большеглазой каталуфы Cookeolus boops, явно происходящих от популяций подводных гор). Также очевидно, что среди среднеглубинных видов есть потомки мелководных форм (уже упоминавшийся щетинозуб Chaetodon guyotensis с хребта Кюсю-Палау — типичный дериват фауны коралловых рифов), возникшие в результате заглубления вершин поднятий. Не вызывает никаких сомнений и вполне очевидный сейчас факт видовой дивергенции рыб в условиях изоляции на горах, имеющих достаточно длительную историю существования. Вопрос о скоплениях рыб у подводных поднятий тоже не остался без ответа (этот аспект обсуждается в гл. VI). Таким образом, все «загадки» К. Хаббза вроде бы разгаданы, но полной ясности в талассобатиальной проблеме все еще нет: на передний план выдвинулись теперь экологические aспекты существования горных популяций. Будущие ихтиологические изыскания на океанских «банках» должны быть направлены прежде всего на изучение размножения, жизненных циклов, миграций и особенностей поведения рыб (здесь никак нельзя обойтись без использования обитаемых подводных аппаратов), а также на выявление популяционной структуры отдельных видов.
Биотопическое своеобразие подводных поднятий, о котором шла речь выше, не может затушевать, однако, очевидного сходства их населения с населением материковых склонов в тех же диапазонах глубин. Повсюду самая верхняя часть среднеглубинной зоны имеет переходный (иначе говоря, экотонный) характер, по батиметрические границы экотона и степень его своеобразия, которую можно выразить процентом видового эндемизма в пределах эпимезобентопелагического комплекса рыб, насчитывающего более 1100 видов (см. табл. 4), сильно варьируют в разных участках Мирового океана.
Ихтиоцены талассной мезобентали отличаются, насколько можно судить по имеющимся данным, главным образом заметным видовым обеднением по сравнению с прилегающими участками континентальных склонов. На Китовом хребте, например, на глубине 200—1500 м отмечено всего 60—70 видов рыб, что в 3—4 раза меньше, чем на соответствующих глубинах склона юго-западной Африки. По Ю. Н. Щербачеву, из 36 видов батиальных офидиид Индийского океана 20 найдены пока только на материковом склоне и лишь 3 только в талассобатиали (13 видов являются общими). Это обеднение связано не только с отсутствием видов, проникающих в мезобеиталь с шельфа, но, по-видимому, отражает и меньшую «емкость» талассобатиали — меньшее разнообразие экологических ниш.
Мезобенталь и прилегающая к ней водная толща заселены хорошо обособленным сообществом рыб, в состав которого входит почти 1200 специфических видов. Правда, число эндемичных для этого биотопа семейств невелико, по к нему приурочено наибольшее разнообразие многих групп. Особенно характерны для этой зоны колючие акулы, в частности из рода Etmcpterus, кошачьи акулы Apristurus, ромбовые скаты, слитиожаберыые угри, гладкоголовы, моровые, долгохвосты (наиболее богатая видами группа донно-придоиных глубоководных рыб), ошибневые, бериксообразпые, солнечиикообразные и др.
Большая часть рыб, населяющих верхнюю часть склона на глубине менее 1500 м, принадлежит к придонным и донным видам, однако среди них есть, как уже говорилось, и чисто пелагические формы, не имеющие непосредственной связи с дном и в какой-то степени аналогичные неритическим рыбам эшгаелагиали. Помимо настоящих пектонных рыб — колючих акул, долгохвлстов, лунников, гемпиловых и др., к этой мезобентопелагической группировке относятся и макропланктонные виды, близкородственные видам, характерным для океанской мезопелагиали. В их число входят, напомню, представители таких семейств, как топориковые (в том числе мавролик), фотихтовые, меланостомиевые, миктофовые, меламфаевые, меланоновые и др. Предполагалось, что основы ареалов этих макропланктонных видов находятся в системе горизонтальных и вертикальных вихрей, существующих над материковым склоном, но прямых доказательств этому нет. Отсутствие у большинства таких рыб значительного выноса за пределы биотопа (вод, покрывающих склоны и подводные поднятия) на любых стадиях жизненного цикла свидетельствует о существовании специального механизма удержания их в присклоновой зоне, сущность которого остается пока непонятной.
Общее число рыб, обитающих в мезобеитали и мезобентопелагиали, намного больше, чем в других океанических ихтиоцеиах. По довольно приблизительному (но отнюдь не завышенному) подсчету (см. табл. 4), в этой зоне обитают около 2600 видов (почти 1500 без экотонпых шелъфо-склоновых рыб). Обособленность этой фауны сильнее всего выражена, на видовом уровне, хотя число родов, обнаруженных только в водах верхней части материкового склона и подводных поднятий, также достаточно велико. В то же время к числу специфических мезобентальных таксонов более высокого уровня можно отнести только относительно немногие семейства, в то время как остальные группы представлены либо па шельфе, либо в нижней части склона, а часто и в обоих биотопах.
В сообществах нижней части склона (батибенталь) и над ней (батибентопелагиаль) представлены сравнительно немногие систематические группы. Из хрящевых рыб здесь живут только колючие акулы из родов Centropho rus, Centroscymnus, Somniosus, кошачьи акулы Apristurus, ромбовые скаты Bathyraja и Breviraja, а также носатые химеры-харриотты, по ни один вид не является эндемичным для биотопа. Среди костистых рыб наиболее характерны виды родов Aldrovandia, Synaphobranchus, Alepocephalus, Bathypterois, Bathysaurus, Antimora, Coryphaenoides, Cataetyx, Paraliparis, Lycenchelys и др. По числу видов и особей в тропиках явно доминируют три группы — гладкоголовы, долгохвосты и ошибневые, а в умеренных и холодных водах две — морские слизни и бельдюговые. Суммарное число видов достигает 450, но большая их часть (273 вида) оказывается общей с нижней мезобенталью (граница двух сообществ батиали представляет собой экотонную зону, расположенную на глубине 1300—1800 м), а многие другие (91 вид) — с абиссобенталью, так что на долю специфической фауны приходится менее 20% состава. В нижней части склона различия между континентальной и талассной батибентальпыми группировками еще слабее, чем в мезобентали. В Индийском океане, например, по данным Ю. И. Сазонова, около 75% видов гладкоголовов встречается как на склоне континентов, так и на подводных поднятиях.
Абиссобентальные и абиссобептопелагическяе ихтиоцены включают около 140 видов, однако эндемичные для ложа океана виды очень немногочисленны. Число специфических родов 10—12, среди них Ericara из гладкоголо-вых, Bathymicrops пз зеленоглазковых, Echinomacrurus пз долгохвостовых, Acanthonus и Typhlonns из ошибневых, Pachycara и Thermarces из бельдтоговых. Остальные виды принадлежат к родам, представленным и в батибептали (Synaphobranchus, Einoctes, Bathytyphlops, CoryI haenoides).
При изучении сообществ термальных источников, обнаруженных недавно в Тихом и Атлантическом океанах па глубине более 2000 м, выяснилось, что в состав этих биоценозов входят и рыбы. Как показали подводные наблюдения, фотосъемки и ловы, вблизи геотермальных излияний Галапагосского рифта встречено более 20 видов рыб (в основном макрурид и офидиид), но почти все они держатся в некотором удалении от мест вытекания теплой, насыщенной сульфидами и лишенной кислорода воды. Единственное исключение составляли особи одного вида, предварительно отнесенные к роду Diplacanthopoma (пока не удалось поймать ни одного экземпляра), которые по-одиночке, парами и тройками постоянно появлялись из кратера источника и снова уходили внутрь. Предполагается, что эта рыба питается непосредственно бактериями, окисляющими сероводород. В термальной воде живет также зоарцида Thermarces cerberus, являющаяся, по всей вероятности, постоянным членом этого своеобразного сообщества; другой представитель того же рода обнаружен только в периферической зоне рифтового «оазиса». Слабовыраженный (только на родовом и видовом уровне) эндемизм гидротермальных рыб резко контрастирует с большой обособленностью бентосных беспозвоночных (подкласс погонофор, надсемейство моллюсков и т. д.).
В глубоководных желобах (хадобенталь) до сих пор обнаружено всего 7 видов рыб, ведущих дойный образ жизни (рис. 42). В их числе три вида ошибневых — Leucicorus atlanticus (наибольшая глубина вылова 6800 м в желобе Пуэрто-Рико), Holcomycteronus profundissimus (7160 м в желобе Сунда у южного побережья острова Ява) и Abyssobrotula galatheae (8370 м в желобе Пуэрто-Рико) и четыре вида морских слизней — Notoliparis macquariensis из желоба Маккуори (глубина 5410 м), N. kurchatovi пз Южно-Сандвичева желоба (5470 м) (в табл. 4 эти два вида условно помещены в графу АХ), N. kermadecensis из желоба Кермадек (6770 м) и Pseudoliparis amblysto-mopsis и неописанный вид этого же рода из Курило-Кам-чатского и Японского желобов (максимальная глубина лова почти 7580 м). Все хадобентальные офидииды встречаются и в абиссобентали (рекордист по глубине — абиссобротула — добывалась в восточной Пацифике всего лишь на глубине 2330 м), тогда как оба рода липарид (они, кстати сказать, описаны А. П. Андрияшевым) эндемичны для глубин более 5500—6000 м. Насколько сейчас известно, основную пищу всех этих видов составляют донные ракообразные (изоподы и амфиподы), но у кермадекских нотолипарисов, а их было поймано сразу 5 в одном трале, в желудках оказались также рыбья чешуя и кости. Хищничество представляется, однако, очень маловероятным для этих некрупных и к тому же вялых и пассивных, судя по всем данным, существ. Их поймали скорее всего там, где они собрались для поедания падали, опустившейся из верхних слоев океана. В яичниках двух самок этого вида было обнаружено 737 и 849 икринок, из которых самые крупные имели 7,5—9 мм в диаметре. И. Нильсен предполагал, что этот вид, подобно батиабиссобентальному карепрокту Careproctus ovigerum, может вынашивать развивающуюся икру в ротовой полости. Можно думать, что в хадобентали существуют в другие виды рыб, но их число вряд ли может быть значительным.
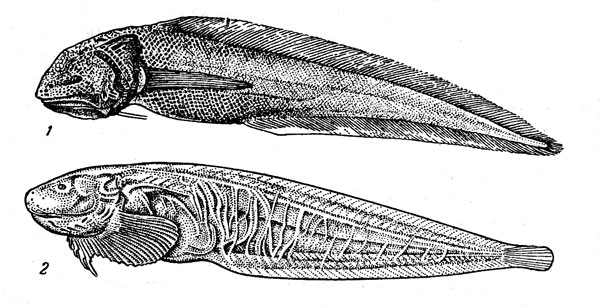
Рис. 42. Рыбы наибольших глубин абиссобротула (1) и псевдолипарис (2).
Все ловы донных рыб в глубоководных желобах выполнены с помощью очень примитивною и, казалось бы, анахроничного орудия лова, которое и в наш век технического прогресса остается таким же, как в прошлом столетии. Это — трал Сигсби, он же трал Агассица, названный так по фамилиям капитана американского корабля «Альбатрос» и выдающегося зоолога, неоднократного руководителя экспедиций на этом же судне. Трал этот предназначен для сбора бентоса.
Представьте себе две крутых дуги из железных пластин, скрепленных трубами и тросами, к которым прикреплен короткий мешок из грубой дели (внутри него помещается мелкоячейная вставка). Эта снасть опускается па дно на длинном тросе и, будучи совершенно симметричной (у нее нет ни верха, ни низа), всегда занимает на грунте рабочее положение. Само траление носит пассивный характер: судно, дрейфующее по ветру, медленно волочит трал, собирающий со дна все, что «плохо лежит» (точнее говоря, то, что лежит хорошо и не способно отплыть или отползти).
Каждое траление в ультраабиссали - большое событие в экспедиционной жизни. Посадка трала Сигсби па дно узкого желоба без применения специальной акустической аппаратуры (а она еще не нашла широкого применения в нашей практике) очень трудна. Она требует точного расчета дрейфа судна, а также оценки необходимой длины вытравливаемого троса (эта оценка, производимая «на глаз»» с учетом двух переменных - глубины по эхолоту и угла наклона троса, уходящего в воду,— самое слабое звено методики работ). Бывает, что трал не достигает дна или. наоборот, лишний трос завивается в кольца, «колышется» и рвется при
выборке, так что долгие часы работы пропадают впустую. Мне приходилось присутствовать при нескольких десятках удачных тралений в самых разных желобах (в том числе и в самом глубоком из них - Марианском), по только дважды - в Японском и Южно-Сандвичевом желобах они приносили рыб. Ясно, что на хадальных глубинах их совсем не так уж много.
Экология донно-придонных рыб открытого океана исследована, пожалуй, еще хуже, чем экология рыб глубоководной пелагиали. Данных о их размножении, росте, питании и других биологических особенностях пока немного, и общая картина, складывающаяся из разрозненных фрагментов, не может считаться вполне адекватной: многие вопросы еще ждут ответа. Тем пе менее уже сейчас выявлены некоторые общие закономерности, характерные для рыб бентопелагического комплекса.
Размножение. Хрящевые рыбы, обитающие в придонных слоях, демонстрируют все три возможных в этой группе типа репродукции — яйцерождение, яйцеживорождение и живорождение. К числу яйцеживородящих принадлежит п латимерия, но из костистых рыб этим путем размножаются только некоторые эпимезобентальные морские окуни (Sebastes) представители семейств Bythitidae и Aphyonidae. Преобладающее большинство видов имеет пелагические икринки, число которых сильно варьирует: от нескольких тысяч (у аргентин и батитифлопсов) до сотен тысяч (у моровых, долгохвостов, крупных камбаловых). У рыб, откладывающих донную икру (икру с отрицательной плавучестью), плодовитость, как правило, заметно меньше и измеряется десятками — сотнями икринок (минимально — 20 икринок у Rinoctes nasutns), по у некоторых видов гладкоголовов (крупные представители рода AlepocepLalus) может достигать 10 тыс.
Судя по встречаемости личинок и мальков в уловах, донно-придонных рыб можно разделить на две группы. У одних постэмбриональное развитие проходит в толще воды, у других — в придонных слоях. К первой группе, для которой характерны значительные морфологические преобразования в конце личиночно-малькового периода (метаморфоз), должны быть отнесены все угри и родственные им галозавриды и нотакантиды, также имеющие лептоцефальную стадию, некоторые хлорофтальмиды (Bathytyphlops с мальком макристиеллой) и сиподонтиды (Balhysaurus с мальком макристиумом), те офидииды, которые обладают специфическими личиночными стадиями типа «эксериллиум» или «вексиллифер», а также, по-видимому, мориды и долгохвосты. Вторую группу образуют алепоцефалиды, хлорофтальмиды из родов Ipnops и Bathymicrops, некоторые офидииды, все липариды и зоарциды и др.
Эта простая классификация не может быть распространена, однако, на всех рыб бентопелагического комплекса, поскольку у многих из них ранние стадии развития все еще остаются неизвестными. Она не является бесспорной даже в отношении некоторых из перечисленных выше рыб. Представлениям о наличии пелагической стадии у всех видов семейств Moridae и Macrouridae противоречит, например, существование в этих группах локального эндемизма: в обоих семействах сейчас известно немало видов, которые ограничены в своем распространении препятствиями, непреодолимыми для них, казалось бы, только во взрослом состоянии — для батибентальных рыб это мелководные пороги, отделяющие глубинные бассейны, для мезобентальных — океанические глубины, окружающие подводные возвышенности. Личиночно-мальковые стадии всех массовых доино-придопных рыб, особенно населяющих батиаль, вообще очень редки в толще воды и почти никогда не встречаются вдали от материковых склонов и, как уже говорилось, горных поднятий в открытых водах.
В связи с этим высказывались предположения о том, что эмбриональное и раннее постэмбриональное развитие таких рыб может проходить в придонных горизонтах.
Размеры и рост. В среднем донно-придонные глубоководные виды рыб крупнее пелагических глубоководных, но мельче тех, которые живут в верхней толще воды (см. рис. 27). Среди них нет таких гигантов, как эпипелагические акулы, марлины и меч-рыбы, но некоторые из батиальных видов достигают все же значительной длины. К числу наиболее крупных относятся полярная акула (длина 6,4 м при массе около 1 т), шестижаберная акула Нехаnchus griseus (4,8 м), акула-домовой (3,3 м), ложнокунья акула (3 м), звездчатошипая акула Echinorhinus brucus (2 м), ромбовые скаты рода Bathyraja (2 м), химеры 1(1,5 м), а также латимерия (длина 2 м, масса почти 100 кг). Из костистых рыб наибольших размеров достигают белокорый палтус (длина до 4,7 м и масса более 300 кг), клыкач (2,1 м), некоторые долгохвосты, ателеоиовые и гемпилиды (по 2 м), слитиожаберные угри (1,8 м), зубатки (1,4 м), иекоторые офпдииды (до 1,3 м), а самыми маленькими являются бентопелагические стерпоптихиды и миктофиды из родов Maurolicus, Polyipmis, Diaphus, Laitipanyciodos, а также бептические липариды и перцихтиды из родов Paraliparis, Osopsaron, Pteropsaron, длина которых не превышает 5—6 см. В абиссали размерный спектр значительно уже: от 19—20 см у некоторых гладкоголовов (Rmoctes, Mirognathus) до 85 см (при массе 7 кг) у батизавра Bathysaurus mollis, который, как показано прямыми наблюдениями с подводного аппарата, способен к резким рывкам на расстояние 10—15 м и, без сомнения, должен быть отнесен к нектону (в толще воды нектонные рыбы отсутствуют в батипелагиали и ниже).
Глубоководные донные и придонные рыбы, как правило, отличаются медленным ростом и большой продолжительностью жизни. Исключение составляют, пожалуй, только мелкие беитопелагические виды (мавролик, светящиеся анчоусы) из семейств, характерных для открытой пелагиали, которые совершают суточные вертикальные миграции из придонных горизонтов в эпипелагиаль. Все они, подобно их чисто пелагическим родичам, созревают в возрасте, близком к годовалому, и живут ненамного больше. У таких рыб, как колючие акулы, аргентины, гладкоголовы, моровые, крупные макруриды (род Coryphaenoides), большеголовы, скорпеновые, камбалы, продолжительность жизни варьирует, насколько сейчас известно, от 10 до 30 и даже 40 лет, причем годовые приросты длины редко превышают 3—5 см. Половозрелыми опи становятся в возрасте 5—10 лет, а иногда я позже. Эти рыбы с полным правом могут быть отнесены к категории длинноцикловых.
Питание. Представляется естественным, что рыбы, живущие на дне или у дна, должны питаться в основном пищей, находящейся на грунте, т. е. бентосными организмами. Это вполне логичное заключение, которое, как правило, оправдывается в приложении к донпо-придонным рыбам шельфовой зоны, оказывается, однако, совсем не бесспорным, когда речь идет о глубоководных видах — почти все массовые формы потребляют в пищу пелагических животных (мезо- и макропланктон, микронектон). Такая «диета» характерна не только для рыб, поднимающихся в водную толщу, но и для многих постоянных обитателей дна.
Значительная часть рыб бентопелагического комплекса относится к числу хищников, поедающих крупную добычу — других рыб, головоногих, ракообразных. Наиболее глубоководными из донных хищников являются батизавры, в пище которых зарегистрированы слитножабериые угри, макруриды, лиценхелы, креветки, кальмары. Известен и способ их охоты: рыбы поджидают добычу, полустоя или лежа на дне в слегка наклонном положении (передняя часть тела приподнята), и ловят ее неожиданным стремительным броском, сопровождаемым хватательным движением челюстей. Некоторые хищники мезобентопелагиалн способны к совместному добыванию добычи. Так поступают, к примеру, некоторые колючие акулы, а также рексии. Эти рыбы режут крупную рыбу на куски своими мощными зубами, а затем заглатывают по частям, и части одной жертвы иногда обнаруживаются в желудках разных хищников, пойманных одним тралом.
Конечно, на тех участках дна, где имеется достаточное количество кормового бентоса, он также идет в пищу рыбам. Многие из преимущественно планктоядных видов, в частности гладкоголовы, долгохвосты, мориды, нередко разнообразят свое питание за счет донных моллюсков, полихет, ракообразных. Как показал Р. Хэдрич, отдельные группы видов (подроды) рода Coryphaenoides, различающиеся размерами и строением зубного аппарата, различаются и по характеру пищи. Мелкие виды со слаборазвитым зубным вооружением (например, С. carapinus) поедают главным образом бентос, в первую очередь офиур, тогда как более крупные виды с сильными зубами — пелагических ракообразных, рыб, головоногих. Преимущественно бентоядными являются многие офидииды, угри, камбалы, чисто бентоядными — галозавры, спиношипы, линариды, встречающиеся в связи с этим только на мягких грунтах.
Интересно сопоставить трофическую структуру глубоководного донного ихтиоцена и таксоцена беспозвоночных того же сообщества. Большая часть глубоководного макробентоса, по М. Н. Соколовой, входит в состав пищевой группировки детритоедов, питающихся органическим веществом донных осадков. Также довольно многочисленны среди донных беспозвоночных сестоноеды, пищу которых составляют взвешенные в воде органические частицы, включая живой планктон. Плотоядные беспозвоночные, потребляющие живую добычу и трупы, представляют меньшинство и в сессильвом (сидячем) и в вагильном бентосе, но решительно преобладают в нектобентосе, (это в основном придонные ракообразные). Среди рыб чистых детритоедов, по-видимому, нет, хотя остатки грунта, заглоченные, можно думать, вместе с другой пищей, не раз обнаруживались в желудках бентофагов (например, у спиношипов и галозавров). К числу сестонофагов-хватателей (почти что сессильных) можно отнести батиптеров, возвышающихся над дном на треножнике, составленном лучами хвостового и брюшных плавников, а возможно, и других глубоководных хлорофтальмид (кроме ипнопсов). Преобладают в ихтиоцене, конечно, плотоядные, среди которых следует выделять потребителей донных беспозвоночных, всеядных хищнп-ков-падалыциков и настоящих хищников, охотящихся только на крупную живую добычу.
Вершинные участки подводных поднятий, как правило, имеют скальное дно, полностью лишенное рыхлых осадков, что само по себе определяет доминирование рыб-планктофагов в сообществах таких биотопов. До самого недавнего времени бытовали два взгляда на источник пищевых ресурсов, определяющих существование и продукцию «горных» популяций рыб. Согласно одному из них, продуктивность поднятий определяется локальным орографическим эффектом — подъемом глубинных вод, обусловленным завихрением набегающего на гору потока постоянного течения и приводящим в действие механизм биологической продуктивности в его полном цикле: от повышения концентрации биогенных солей в эвфотическом слое к увеличению первичной продукции водорослей и за ее счет к увеличению биомассы зоопланктона. Для более точного («более наукообразного») объяснения этого явления была выдвинута гипотеза о формировании над вершинами квазистационарных вихрей, которые п обеспечивают постоянный подъем вод, будучи настолько изолированными от окружающих вод, что внутри них образуются самостоятельные экологические системы. Действительно, подводный рельеф, усложняя в известной степени структуру набегающего потока, может способствовать образованию локальных вихрей, приводящих в конечном счете к обогащению поверхностных слоев биогенными элементами. Однако пространственно-временная устойчивость таких впхрей представляется довольно слабой, а их продолжительная изоляция — в высшей степени проблематичной. Ведь время, необходимое для развития фитопланктона, исчисляется несколькими сутками, а кормовая база рыб — мезопланктон и макропланктон — развивается намного дольше, соответственно около месяца и полугода.. Столь длительное существование над подводной возвышенностью неподвижных и существенно замкнутых вихревых образований почти невероятно: если влияние локального обогащения поверхностных вод биогенами и может отразиться на продуктивности зоопланктона и тем более па рыбопродуктивности, то произойдет это только в значительном удалении от места подъема вод. Таким образом, эффект увеличения биомассы кормовых организмов за счет локального «апвеллинга» может быть использован не бептопелагическими рыбами той подводной возвышенности, которая послужила причиной возникновения такого «шлейфового» пятна повышенной биомассы планктона, а только истинно пелагическими хищниками (например, тунцами и кальмарами). Поэтому гораздо более правомочным представляется другой подход, связывающий кормность поднятий с горизонтальным приносом (адвекцией) пищевых объектов набегающим потоком. Прямые исследования питания рыб, живущих над подводными возвышенностями, показывают, что их откорм происходит в основном за счет макропланктона, входящего в состав мигрирующих звукорассеивающих слоев (миктофиды, эвфаузииды и т. п.) и опускающегося днем на самую вершину горы (если, конечно, она лежит выше нормальной глубины дневного опускания этих слоев). Распространение. Географическое распространение океанических рыб, входящих в состав бентопелагического комплекса, еще не изучено в деталях, но представляется, что размеры их ареалов сильно варьируют. Некоторые из них весьма обширны — такие виды, как, например, эпимезобентальные антигония Antigonia capros и рыба-бекас Macrorhamphosus scolopax, мезобентальпые мягкоголовый долгохвост Malacocephalus laevis и низкотелый берикс Вегух splendens, мезобатибеитальные альдровандия Aldrovandia phalacra и антимора Antimora roslrata, батибентальные гладкоголов нарцет Narcetes sloimias и ксиелациба Xyelacyba myersi, батиабиссобентальные гладкоголов беллоция Bellocia koefoedi и батизавр Bathysaurus mollis, абиссобентальные слепой тифлонус Typhlonus nasus и эхиномакрурус Echinomacrurus mollis, обнаружены на соответствующих глубинах в тропических водах всех трех океанов. В то же время во всех вертикальных зонах есть и виды, которые известны из очень ограниченных участков и, по-видимому, на самом деле имеют очень узкое распространение (хотя представления об эндемизме могут основываться на недостаточных данных). Крайний случай почти «точечного» ареала демонстрирует распространение латимерии, обитающей только на склонах двух маленьках островов Коморского архипелага. Большинство других узколокальных эндемиков также обитает на вершинах подводных поднятий.
Известно, что дробность биогеографического деления бентали, достигающая наибольшей выраженности на шельфе и в верхней части склона (т. е. на глубинах залегания поверхностных и подповерхностных вод), уменьшается в нижней батиали и сводится к минимуму на ложе океана, где можно обнаружить только самые главные границы (те, которые разделяют регионы высшего ранга — области). Многими работами показано, что главные зоогеографические границы в мезобентали как бы продолжают вглубь границы, проходящие на шельфе. Действительно, анализ географического распространения рыб, населяющих склоны континентов и подводные поднятия, дает возможность выявить те же крупномасштабные регионы, что и в сублиторальной зоне. Так, Г. А. Го-ловань, рассматривая ареалы мезобентальных рыб, обитающих у западных побережий Африки и Eвропы, обнаружил в составе ихтиофауны 12 географических элементов и выделил четыре района — бореальный, северный субтропический, тропический и южный субтропический — в районировании шельфа.
Немногочисленность поимок многих видов батибентальных рыб сильно затрудняет анализ их географического распространения. Тем не менее представления о преобладании в этой зоне узколокальных ареалов, существовавшие до недавнего времени, не подкрепляются, как правило, новыми данными. Почти для всех видов, известных по многим нахождениям, характерно широкое распространение. Достаточно четкое разделение существует, однако, между фаунами тропических, умеренных и холодных вод. В тропической зоне выделяются экваториальный и широкотропический типы ареалов с циркумглобальным, индовестпацифическим, атлантическим и восточно-тихоокеанским подтипами. Умеренноводные ареалы подразделяются на потальные и бореальные, а холодноводные — на антарктические и арктические.
Что касается абиссобентальных и хадальных рыб, все они известны по немногим, часто единичным находкам, что сильно затрудняет рассмотрение их зоогеографии. Нужно отметить, впрочем, что на ложе океана некоторые виды уже сейчас найдены в далеко разобщенных районах (в двух или трех океанах встречены Bathymicrops regis, Ipnops meadi, Holcomycteronus profundissimus, Abyssobrotula galatheae и др.), и это позволяет предполагать их широкое распространение.