
Фауна и экология донных беспозвоночных лососевых рек урала и тимана 03. 00. 16- экология
| Вид материала | Автореферат |
СодержаниеГлава 7. влияние антропогенных факторов на зообентос лососевых рек Глава 8. бентос в пище основных видов рыб |
- Фауна, биология и экология рода anopheles mg. (Сем. Culicidae) в прибрежных экосистемах, 1214.31kb.
- Программа курса Методика организации курса \"Экология Тамбовской области\", 78.43kb.
- Экология Вопросы к зачёту, 25.45kb.
- Птицы селитебных ландшафтов северной части центральной азии (фауна, население и экология), 1158.65kb.
- Лекция 1 Что такое экология. Разделы экологии Термин «экология», 148.25kb.
- Тематика лекций (Наименование тем, содержание, объём в часах) № п/п, 74.97kb.
- Экология как наука Экология, 193.74kb.
- Промышленная экология) учебно-методический комплекс по дисциплине Прикладная экология, 384.12kb.
- Библиографический указатель книг, поступивших в библиотеку в декабре 2011, 2435.14kb.
- Комплексы беспозвоночных-сапрофагов в лесных экосистемах Кольского Севера 03. 00., 652.44kb.
Примечание. В числителе – средняя численность, экз./м3; в знаменателе – средняя биомасса, мг/м3; «–» – данные отсутствуют.
Каждый вид беспозвоночных отличается спецификой вовлечения в дрифт, обусловленной особенностями их биологии. В период открытой воды меняется роль отдельных беспозвоночных в дрифте. В мае отмечены массовые миграции поденок и хирономид, значительную долю по биомассе составляют ручейники. В июне-июле основу дрифта формируют личинки хирономид и частично – поденок; в августе-сентябре помимо них большое значение приобретают личинки веснянок и ручейников. Глубокой осенью (в октябре) перед ледоставом снос беспозвоночных невелик: мигрирует лишь пять групп бентоса, доминируют по численности личинки хирономид и поденок, а по биомассе кроме них – еще и личинки веснянок.
Сведения о дрифте донных беспозвоночных в период ледостава, продолжительность которого на лососевых реках Европейского севера России составляет почти семь месяцев, в литературе отсутствуют. Нами исследования выполнены на стационаре р. Щугор 13–14 апреля 1986 г. и 28–29 марта 1987 г. В подледный период в дрифте обнаружено восемь групп донных беспозвоночных (табл. 8); как и в период открытой воды, доминируют личинки поденок, веснянок и двукрылых, на долю которых приходится 59–95% общей численности и почти 100% общей биомассы мигрирующих организмов. Показатели количества и биомассы организмов в дрифте в марте и апреле оказались близкими к таковым осенних проб и между собой. В период закрытого русла рек в сравнении с летом происходит снижение количественных показателей дрифта и уменьшение в нем числа групп беспозвоночных. В пробах в это время присутствовали экзувии личинок поденок и веснянок, что свидетельствует о линьке этих гидробионтов, связанной с их ростом и развитием в период ледостава. Изменяющиеся потребности в процессе развития некоторых видов поденок и веснянок к условиям обитания, по-видимому, и являются одной из основных причин их миграций подо льдом.
Таблица 8
Состав беспозвоночных в дрифте р. Щугор в период ледостава
| | 13-14.04. 1986 г. | 28-29.03. 1987 г. | ||
| Беспозвоночные | доля, % по численности | доля, % по биомассе | доля,% по численности | доля,% по биомассе |
| | | | | |
Nematoda | 0.4 | <0.1 | 0.9 | <0.1 |
| Harpacticoida | – | – | 0.5 | <0.1 |
| Другие Copepoda | 4.7 | 0.2 | 39.4 | 1.4 |
| Hydracarina | – | – | 0.5 | 0.1 |
| Ephemeroptera, lv. | 16.3 | 20.4 | 25.8 | 44.8 |
| Plecoptera, lv. | 1.6 | 65.1 | 2.8 | 49.9 |
| Simuliidae, lv. | 6.6 | 2.5 | 0.5 | 0.3 |
| Chironomidae, lv. | 70.4 | 11.8 | 29.6 | 3.5 |
| Средняя численность, экз./м3 | 1.94 | | 1.61 | |
| Cредняя биомасса, мг/м3 | | 0.60 | | 0.45 |
Примечание. «–» – не установлены.
6.3. Суточная ритмика. Численность и биомасса беспозвоночных в дрифте изменяются не только по сезонам, но и в течение суток. Суточные сборы дают возможность получить более полную информацию о качественном и количественном составах мигрирующих донных организмов и избежать ошибок, возможных при взятии разовой пробы. Изучение суточной динамики дрифта донных беспозвоночных представляет и практический интерес, так как эти сведения важны при рассмотрении суточной ритмики питания молоди рыб в лососевых реках. Миграции донных беспозвоночных «в массовом количестве при низкой освещенности способствуют меньшему выеданию их рыбами» (Смирнов и др., 1978).
Исследования суточной динамики дрифта беспозвоночных выполнены на стационаре р. Щугор в апреле, июне–сентябре, ноябре–декабре. В дни исследований уровень воды в реке в течение суток не изменялся и динамика мигрирующих беспозвоночных не зависела от его колебаний. Временные отличия в миграциях беспозвоночных связаны с особенностями их суточной активности. Считается (Леванидова, Леванидов, 1962; Waters, 1962, 1972; Müller, 1966; Thomas, 1970), что суточная ритмика донных животных обусловлена освещенностью, которая действует по принципу "включать-выключать", повышая активность многих беспозвоночных, когда освещенность уменьшается до некоторого предельного уровня интенсивности. Доминирующие по количеству в дрифте лососевых рек поденки и веснянки, как животные с отрицательным фототаксисом (Waters, 1972; Bishop, Hynes, 1969), наиболее активны при низкой освещенности и переходят в дрифт преимущественно в ночное время. Преобладание в дрифте лососевой р. Щугор беспозвоночных с ночной активностью обусловливает в период ледостава, как и в период открытой воды, выраженный пик численности и биомассы общего дрифта донных беспозвоночных в ночные часы в – 0 ч (рис. 5).


Рис. 5. Суточная динамика численности (А) и биомассы (В) донных беспозвоночных в дрифте р. Щугор. Верхний рисунок – в период ледостава (13-14.04.1986 г.), нижний – в период открытой воды (29-30.07.1986 г.)
В работе показано, что сходные картины суточной ритмики дрифта беспозвоночных установлены и для других лососевых рек Урала и Тимана.
ГЛАВА 7. ВЛИЯНИЕ АНТРОПОГЕННЫХ ФАКТОРОВ НА ЗООБЕНТОС ЛОСОСЕВЫХ РЕК
Изучение антропогенного воздействия на экосистемы рек имеет важное практическое значение для прогнозирования предупредительных мер по охране водотоков от загрязнения. Рассматривается влияние на донных беспозвоночных лососевых рек различных форм хозяйственной деятельности: 1. Строительство веток газопровода СРТО–Торжок на реках Щугор, Светлый Вуктыл (Северный Урал); 2. Разработка россыпных месторождений золота на р. Кожим (Приполярный Урал); 3. Сброс в реки промышленных, сельскохозяйственных и бытовых стоков, разработка бокситовых месторождений, интенсивное сведение лесов, многолетний лесосплав и др. (Тиман: реки Ухта, Ижма, Мезень, Вымь).
Наиболее показательные изменения выявлены на количественных характеристиках типичных для лососевых рек поденках, веснянках, жуках и ручейниках. С увеличением загрязнения резко снижаются их численность и биомасса (рис. 6), сокращается и видовое разнообразие этих отрядов. Например, если на чистом и слабо загрязненном участках р. Ухта (Тиман) установлено восемь видов веснянок, 20 видов поденок, четыре вида жуков, 19 – ручейников, то на грязном участке присутствуют по два вида веснянок, жуков и ручейников и четыре вида поденок. На территории промышленной зоны в зообентосе этой реки остались резистентные к загрязнению виды беспозвоночных. Подобная закономерность установлена и для загрязненных участков лососевых рек Северного и Приполярного Урала.
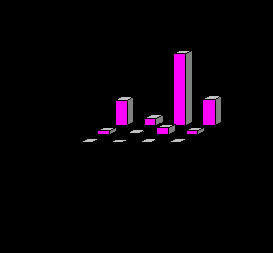
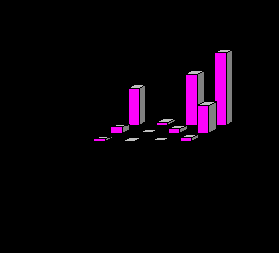
Рис. 6. Численность (А) и биомасса (В) типичных обитателей дна лососевых рек на участках р. Ухта: I – чистом; II – слабозагрязненном; III – грязном.
Загрязнение среды обитания гидробионтов в результате различных форм хозяйственной деятельности ведет к деградации и структурным перестройкам исходных литореофильных биоценозов уральских и тиманских рек, к обеднению их флоры и фауны за счет элиминации характерных для чистых вод видов, к снижению общего продукционного потенциала донных беспозвоночных, к возрастанию в летнюю межень катастрофического дрифта беспозвоночных. Нарушается стабильность речной экосистемы, ослабевает сопротивляемость неблагоприятным внешним воздействиям. Даже небольшое по масштабам загрязнение представляет для населения лососевых рек повышенную опасность из-за снижения на севере способности среды к самоочищению и восстановлению при суммарно низких температурах воды.
Восстановление бентоса лососевых рек на участках с нарушенными гидрологическим и гидрохимическим режимами после окончания хозяйственных работ идет медленными темпами. Зарегистрирована слабая тенденция к небольшому росту численности и биомассы бентоса; не установлено возрастание видового разнообразия основных представителей фауны. Процесс восстановления водных экосистем с завершением хозяйственных работ – одна из основных проблем, требующих разрешения.
ГЛАВА 8. БЕНТОС В ПИЩЕ ОСНОВНЫХ ВИДОВ РЫБ
Состав пищевого рациона рыб лососевых рек Урала и Тимана разнообразен: он включает донных беспозвоночных, рыбную и растительную пищу, воздушных насекомых. Рыбы высоких широт, которым принадлежит исследованная нами территория, при преимущественно низкой температуре среды эффективно усваивают широкой спектр пищевых объектов. Значительная эврифагия рыб достигается благодаря реализации нутритивных и температурных адаптаций пищеварительной системы и ее универсальным набором ферментных систем, адекватно реагирующих на изменения факторов экзо – и эндогенной природы (Пономарев, Шубина, Черезова, 1998).
В главе рассматривается роль бентоса в питании наиболее многочисленных и широко распространенных видов рыб в лососевых реках Урала и Тимана: сига-пыжьяна, европейского хариуса и молоди семги. Главные компоненты пищевого рациона сига – моллюски, поденки, ручейники и хирономиды, значимость которых меняется в зависимости от места обитания рыбы и сезона года.
Основу пищевого рациона хариуса и молоди семги составляют донные организмы: беспозвоночные, водоросли и мохообразные. Помимо основных гидробионтов лососевых рек (поденок, ручейников, двукрылых, моллюсков) в пище хариуса и молоди семги встречены и редкие, такие как волосатики, пиявки и др., которые в соответствии с современными концепциями питания и пищеварения могут оказаться источником ряда незаменимых аминокислот, витаминов и биологически активных веществ (Уголев, Кузьмина, 1993). В летний период на долю растительных остатков по массе приходится 12-15 % пищевого комка рыб (Пономарев, Шубина, Серегина, 2000). Структурные полисахариды растительной пищи традиционно считались как ненужный для рыб балласт, однако в последние годы доказано их жизненно важное значение как субстрата для эндофлоры и ключевого звена для формирования потоков вторичных нутриентов (Уголев, 1985). С возрастом в питании хариуса и молоди семги снижается роль донных беспозвоночных, меняется состав доминирующих пищевых компонентов, увеличивается в рационе значение рыбной пищи.
Пищевые взаимоотношения рыб в реках Урала и Тимана рассмотрены на примере ручейников, которые в течение всего года относятся к числу постоянных и важнейших компонентов питания хариуса и молоди семги. В период открытой воды ручейники в пище рыб составляют в среднем 40–52 % от массы комка, с возрастом рыб встречаемость, численное и весовое содержание ручейников в пище увеличивается. Рыбы используют в питании личинок, куколок и имаго этих насекомых, но преимущественно личинок, на долю которых приходится 97 % от количества и 77 % от массы всех потребленных ручейников. Роль взрослых форм невелика (0.3 % от веса комка); доля куколок в период открытой воды варьирует в зависимости от гидрологических условий года. В годы с затяжной холодной весной в сравнении с теплой развитие ручейников задерживается: еще в июне–июле их куколки, уже покинувшие свой домик, непосредственно перед вылетом, находятся в числе основных компонентов пищи рыб. На долю куколок приходится до 40 % от массы потребленных ручейников, а в эти же месяцы теплого года – лишь 5 %. К осени в теплые годы возрастает значение (по массе) ручейников в питании рыб, в годы с низкой температурой и неустойчивым водным режимом наблюдается обратная картина.
Большое количество видов ручейников в бентосе рек приводит к их значительному разнообразию и в пище рыб, однако по биомассе, как правило, преобладают один–три вида ручейников, массовые в бентосе. Состав доминирующих видов в пище хариуса и молоди семги сходен и не зависит от возраста рыбы и ее пола, но определяется условиями года, сезоном и спецификой фауны ручейников участка реки. Наибольшая степень совпадения по массе ручейников (до 73 %) в пище хариуса и молоди семги установлена в конце летней межени и осенью.
ЗАКЛЮЧЕНИЕ
Современная речная сеть Северного, Приполярного Урала и Тимана формировалась на протяжении длительного периода и за это время претерпела значительные изменения. История развития и возраст современной речной сети Урала и Тимана определяются различными исследователями по-разному. По мнению одних (Зеккель, 1940), современная гидрографическая сеть была заложена еще до первого оледенения, быть может, в третичный период. Другие исследователи (Варсанофьева, 1929; Добролюбова и Сошкина, 1935; Чернов, 1944; и др.) считают, что современная гидрографическая сеть Урала начала формироваться в последний межледниковый период. По мнению В. А. Варсанофьевой (1961), в четвертичный период Урал подвергался трем плейстоценовым покровным оледенениям: рисскому (днепровскому), вюрмскому (московскому), не исключается возможность покрытия территории и древнечетвертичным ледником. Однако новыми геологическими данными ставится под сомнение наличие здесь вюрмского оледенения (Генералов и др., 1970; Дегтяренко и др., 1970; Палеогеография..., 1982; и др.).
Согласно гипотезе о геогидрократических колебаниях уровня Мирового океана (Линдберг, 1972) во время четвертичного периода существовали повсеместные неоднократные морские трансгрессии, две из которых, особенно мощные, простирались в глубь материков и достигали в высоту 180 м (более ранняя) и 80 м (более поздняя). Основываясь на этой гипотезе, можно предположить, что часть бассейнов уральских рек западного склона Урала с гипсометрическими отметками до 200 м покрывало море, а в Уральских горах не исключается возможность горно–долинных оледенений. По сравнению с Уральским хребтом Тиманский кряж – более древнее, сильно разрушенное, горное сооружение (Исаченко, 1964), связанное с Уралом.
Некоторые группы донных беспозвоночных (веснянки, поденки, ручейники, хирономиды) сформировались еще в пермский период, мшанки – в силурийский, моллюски, олигохеты, пиявки, остракоды известны с кембрийского периода. Богатый биофонд водотоков исследованных территорий, особенности состава и размещения беспозвоночных лососевых рек Урала и Тимана, наличие древних элементов фауны и флоры, доминирование древних отрядов насекомых свидетельствуют о многовековой эволюции, о продолжительном пребывании гидробионтов в этих реках. Полученные нами материалы поддерживают высказывания биогеографов (Кузнецов, 1938) о возможности сохранения древней дочетвертичной флоры и фауны в предгорной и горной полосах Урала.
До настоящего времени многие лососевые реки Урала и Тимана сохраняют естественный режим, поэтому изучение их водной растительности, планктона, бентоса, дрифта беспозвоночных, питания рыб позволяет установить для водотоков этих регионов норму по данным показателям, которые служат основой для общего познания структуры и динамики естественных биоценозов горных рек. Биологические показатели состояния экосистем (видовое разнообразие, сообщества, популяционные и организменные показатели т.д.) интегрируют все изменения за продолжительный период времени (Ramm,1988).
В последние годы в результате интенсивного освоения богатых природных ресурсов Урала и Тимана возрастают масштабы различных форм водопользования, многократно увеличиваются нагрузки на водные экосистемы исследованных регионов. Экологические исследования в Уральском и Тиманском регионах по изучению закономерностей формирования речных экосистем и техногенных нарушений необходимо продолжить и расширить, при этом необходимо знать и потенциальные возможности изменений. Следует безотлагательно решить вопрос о технической и биологической рекультивации нарушенных территорий в бассейнах лососевых рек Северного, Приполярного Урала и Тимана, без чего дальнейшее планирование здесь работ с позиций охраны окружающей среды лишено всякой логики. При разработке проектов освоения природных ресурсов в бассейнах лососевых рек необходим жесткий режим охраны всех звеньев сложной речной системы с учетом региональных особенностей стока, гидрографии и биологии водоемов. Дальнейшее использование водных ресурсов горных стран Урала и Тимана необходимо соотносить с минимальным вмешательством в природную среду.
ВЫВОДЫ
1. Специфические условия обитания гидробионтов в лососевых реках Урала и Тимана дают возможность преимущественного развития донных беспозвоночных, экологический характер которых определяют холодолюбивые реофильные и близкие к ним виды, предъявляющие высокие требования к кислородному режиму и предпочитающие стабильные твердые грунты. Однако для большого числа речных беспозвоночных характерна устойчивая, выработавшаяся исторически в процессе эволюции, сравнительно широкая в отношении абиотических факторов экологическая пластичность, обусловленная адаптацией к динамичности условий среды лососевых водотоков (как горных). Доминирует в исследованных реках литореофильный биоценоз, который характеризует речное донное население в целом.
2. Древность Урала и Тимана, широкий спектр экологических факторов среды их водотоков обусловливают богатое видовое разнообразие литореофильного биоценоза. В результате проведенных исследований (1958-2006 гг.) в его составе зарегистрировано 58 видов мохообразных, более 500 видов водорослей и свыше 840 видов беспозвоночных. Наибольшим видовым разнообразием отличаются литореофильные биоценозы рек в горной и предгорной областях. Видовое богатство фауны лососевых водотоков определяют древние отряды амфибиотических насекомых: поденки, веснянки, ручейники и двукрылые (семейство хирономид), а также водные клещи и олигохеты. Подавляющее большинство видов водных растений и животных для исследованных регионов Палеарктики указывается впервые.
3. Особенность географического положения, многообразие природных условий, сложное геологическое прошлое Урала и Тимана определяют гетерогенность фауны водных беспозвоночных, которая в зоогеографическом плане представлена видами: голарктами (доля невысока – до 10%); палеарктами (наиболее многочисленны); характерными европейскими видами, за пределами Урала на востоке не известными; сибирскими видами, отсутствующими в водоемах Европы. Видовое разнообразие сибирских видов небольшое, но именно они вносят оригинальность и своеобразие в состав фауны лососевых рек. Значительной оказалась доля северных видов, свойственных главным образом тундровым и таежным водоемам Палеарктики. В реках Тимана в сравнении с водотоками Урала установлено больше европейских видов и западных палеарктов и на порядок меньше видов сибирских. Видовое сходство донной фауны рек Урала и Тимана высокое – более 60% по Серенсену и обусловлено генетической близостью территорий, общностью гидрологического, гидрохимического и термического режимов.
4. Показатели количественного развития зообентоса исследованных водотоков колеблются в зависимости от горной страны и степени минерализации вод. По биологической продуктивности зообентоса, согласно классификации Альбрехта (Albrecht, 1959), лососевые реки Урала и Тимана относятся к рекам средней кормности. Основная масса донных беспозвоночных сосредоточена в моховых обрастаниях на галечно-валунных перекатах с поверхностной скоростью течения 0.6–1.2 м/с. Количественное развитие зообентоса лососевых рек определяют амфибиотические насекомые: поденки, веснянки, ручейники и хирономиды. Среди поденок наиболее многочисленны виды из родов Baetis, Heptagenia, Ephemerella, среди веснянок – Capnia, Diura, Arcynopteryx, среди ручейников – Arctopsyche, Hydropsyche, Brachycentrus, Apatania, Athripsodes, среди хирономид – Thienemannimyia, Cricotopus, Eukiefferiella. В тиманских реках с высокой минерализацией вод велико значение в бентосе моллюсков рода Lymnaea. При сравнительно большом видовом разнообразии зообентоса массовое развитие получают немногие виды, состав которых определяется климатическими условиями года, сезоном и спецификой участка реки.
5. В лососевых реках Урала и Тимана в конце ледостава наблюдается повышенное значение общей биомассы донных беспозвоночных вследствие развития и роста в зимний период доминирующих видов поденок, веснянок и ручейников. Минимальные показатели биомассы зообентоса зарегистрированы весной после вылета массовых видов насекомых, закончивших водный цикл развития. К лету идет нарастание биомассы за счет нарождения нового поколения беспозвоночных и их интенсивного роста. За период открытой воды кривая динамики биомассы зообентоса – одновершинная, как правило, с максимумом, в летнюю межень. Однако, в зависимости от климатических условий года этот максимум может быть в июле, августе или даже в сентябре.
6. Значительные межгодовые и сезонные колебания за период открытой воды установлены для общей численности донных беспозвоночных, в сооотношении их главных групп и объясняются преобладанием амфибиотических насекомых, момент появления имаго которых зависит от климатических условий весны и лета. Биомасса донного населения в целом за период открытой воды остается сравнительно стабильной благодаря разнообразию видов главных групп зообентоса с различными жизненными циклами: наблюдается замещение видов, причем в холодные годы доминируют более крупные виды, но с невысокой численностью.
7. Наибольшим перемещениям вниз по течению в речном потоке (дрифту) в исследованных регионах подвержены поденки, веснянки и хирономиды, в основном, в молодом возрасте и в период завершения водного цикла развития. Количество и биомасса донных беспозвоночных в дрифте лососевых рек варьируют по сезонам, не остаются постоянными они и в течение суток, что обусловлено поведенческими реакциями и биологическими особенностями организмов. Благодаря отрицательному фототаксису активно мигрирующих видов поденок, веснянок и хирономид дрифт беспозвоночных в лососевых реках возрастает в сумерки и ночью.
8. В период интенсивного нагула сига, хариуса и молоди семги кормовые ресурсы рек Урала и Тимана разнообразны, богаты, относительно стабильны, доступны рыбе по размещению и размерам беспозвоночных и обусловливают высокие биологические показатели и значительную степень накормленности рыб. Исследования выявили, что в настоящее время кормовая база большинства лососевых рек Урала и Тимана, сохранивших естественный режим, не является причиной снижения численности рыб.
9. В современных условиях при интенсивном промышленном освоении богатых минеральных и органических ресурсов Урала и Тимана усиливается нагрузка на реки этих регионов, что, в свою очередь, негативно сказывается на формировании их гидробиологического режима. Загрязнение водотоков Урала и Тимана в результате различных форм хозяйственной деятельности ведет к деградации и структурным перестройкам исходного литореофильного биоценоза, к обеднению водной фауны за счет элиминации наиболее характерных видов, к снижению общего продукционного потенциала гидробионтов, к возрастанию в летнюю межень катастрофического дрифта донных беспозвоночных. Любое нарушение среды обитания гидробионтов в лососевых реках, произведенное человеком, ведет или к значительным затратам на ее восстановление, или к невосполнимым потерям.
10. Полученные сведения об экологических особенностях водных экосистем Урала и Тимана, биологический материал по их фауне и флоре – база данных для организации мониторинговых работ за изменением донного населения под влиянием антропогенного воздействия и при решении проблемы охраны вод на европейском Севере.