
3. биогеохимия биоты в природных ландшафтах и зоне геотехногенеза
| Вид материала | Документы |
СодержаниеМолибден в полыни гмелина на территории шерловогорского The molybdenium in the gmelini wormwood in the sherlovogorsk Tungsten in the gmelen’s |
- 1. минералого-геохимические процессы в техногенных и геотехногенных ландшафтах, 768.05kb.
- Функциональная структура биоты в природных экосистемах камчатки 03. 02. 08 Экология, 777.4kb.
- Кционировании техно-природных систем и другой антропогенной деятельности для последующего, 582.73kb.
- Основная образовательная программа высшего профессионального образования уровня бакалавриата, 1699.69kb.
- Патофизиология регионарного кровообращения и микроциркуляции, 124.19kb.
- 1. минералого-геохимические процессы в природных и геотехногенных ландшафтах особенности, 1666.92kb.
- Спросите любого сталкера, какое время суток в Зоне самое страшное? Икаждый ответит, 4342.79kb.
- Терминология, 90.36kb.
- Моделирование нейтронного потока в активной зоне ввэр с помощью нейросетевых технологий, 51.73kb.
- Ионова Наталья Владимировна, ст преподаватель Учебных часов: лекций 30 Практические, 98.17kb.
МОЛИБДЕН В ПОЛЫНИ ГМЕЛИНА НА ТЕРРИТОРИИ ШЕРЛОВОГОРСКОГО
ГОРНОРУДНОГО РАЙОНА
Г.А. Юргенсон1, Е.В. Малышева2
1Институт природных ресурсов, экологии и криологии, г. Чита, Россия, yurgga@ mail.ru
2Лаборатория минералогии и геохимии ландшафта г. Чита, Россия, yurgga@ mail.ru
THE MOLYBDENIUM IN THE GMELINI WORMWOOD IN THE SHERLOVOGORSK
MINING AREA
G.A .Yurgenson, E.V.Malysheva
Institute of Natural Resources, Ecology and Criology SB RAS, Chita, Russia, inrec.sbras@mail.ru
The article deals with the first data about molibdenium content in the Gmelin wormwood in Sherlovogorsk mining area on the sites with different molibdenium content in geological substrate components.
It is revealed that molibdenium allocation in roots, stalks, leaves, blossom is extremely irregular. The tendency to its maximal content in leaves, roots in the well-developed topsoil is discovered. The authors suppose that molibdenium content in blossom is optimal for species purity convention.
Известно, что представление об обязательной токсичности тяжелых металлов является заблуждением, так как в эту группу попадают Cu, Zn, Mo, Co, Mn, Fe, то есть те элементы, большое позитивное биологическое значение которых давно обнаружено и доказано [2, 3]. Основополагающим в этом отношении является знание о концентрации этих элементов в почве, растении и, в конечном случае, в биологических жидкостях и тканях животных. Основным источником поступления молибдена в живые организмы является геологический субстрат, на котором развиваются коры выветривания и почвы. Среднее содержание молибдена в земной коре составляет 1,1 г/т.. В главных типах горных пород составляет(г/т): в ультраосновных – 0,2, в основных – 1.4, средних – 0,9, кислых – 1,0, осадочных (глины и сланцы) – 2,0. Естественное содержание в почвах – 1,0 – 2,0 г/т [4]. Это средние значения. Общие вариации значительно шире и находятся в пределах 0,013 – 17,0 г/т [1].
Молибден испытывает сродство к железу и кислороду (сидерофильность) и к сере (халькофильность). В почве молибден встречается как в виде первичных эндогенных минералов, таких, как повеллит (CaMoO4), молибденит (MoS2), молибдошеелит (Са(МоО4, WO4)). Кроме того, известны молибдаты урана (молуранит (U24+ [MoO4]3 [OH]2·10H2O)) и иригинит (U26+[MoO4][OH]4·4H2O), развитые как в эндогенных, так и в экзогенных условиях.
В почвах преобладают соединения шестивалентного Mo, например, оксид молибдена молибдит (МоО3) и производные молибденовой кислоты H2MoO4 в виде молибдат-иона MoO42-. Соединения шестивалентного молибдена характерезуются заметной подвижностью в щелочной среде. В кислой же среде миграционная способность молибдена может снижаться за счет адсорбции аниона MoO42- положительно заряженными коллоидами гидроксидов железа и алюминия. Поэтому в регулировании концентраций молибдена в почвенном растворе большое значение имеет pH среды. В разбавленных растворах при pH 5 и выше преобладает форма MoO42-, при pH 2,5-1,0 господствует H2MoO4; при pH<2 появляются катионные формы, такие как линдгренит Сu3 (MoO4)2(OH)2 и ферримолибдит Fe2(MoO4)3 ·8H2O и другие. На территории Шерловогорского района распространены черноземы и каштановые почвы. Реакция почвенного рствора является слабощелочной pH = 7,2-7,5. Исходя из этого, можно сделать предположение, что в почвах Шерловогорского района молибден находится в виде MoO42- .
Молибден – жизненно необходимый для растений микроэлемент [2, 3]. Это важный компонент нитрогеназы и нитратредуктазы, присутствует он также и в других ферментах (оксидазах), которые ускоряют разнообразные реакции. Основная ферментативная роль молибдена связана с функцией переносчика электронов, что, по-видимому, находит отражение в переменной валентности элемента. Эти два молибденсодержащих фермента играют важную роль в метаболизме азота, участвуя как в фиксации атмосферного азота, так и в восстановлении NO3¯. Таким образом, потребность растений в молибдене, по-видимому, связана с их обеспечением азотом. Также очень важная потребность в молибдене объясняется прежде всего участием соединений металла в восстановлении нитратов, нитритов и гидроксиламинов до аммиака и, следовательно, важной ролью в биосинтезе аминокислот. Отсюда можно сделать вывод, что молибден является биофильным микроэлементом, поскольку он принимает активное долевое участие в построении живого вещества.
Несмотря на то, что легкорастворимые формы молибдена активно извлекаются из почв растениями, а некоторые виды растений известны как его концентраторы, в полевых условиях фитотоксичность этого элемента проявляется нечасто. Как правило, фитотоксичность молибдена проявляется только при очень высоких концентрациях. Например, по данным Дейвиса и др., признаки молибденового отравления у молодых проростков ячменя отмечались при содержании молибдена 135 мг/кг сухой массы [2].
В данной работе рассмотрено поведение молибдена в полыни Гмелина (Artemisia Gmelini) на территории Шерловогорского горнорудного района. Полевые работы проводились в первой декаде сентября 2007 года, когда вегетационный период был практически уже завершён. Растительные пробы были отобраны на участках: сопка Мелехинская, Пятисотка, падь Лукаво-Золотая, Поднебесных, Северный отвал, Жила Новая, хвостохранилище. Каждая проба растений формировалась из 10-20 экземпляров с площадки 4х4м. Общее число экземпляров растений с одного участка составляло около 40 штук, что позволило для обработки аналитических данных использовать методы матстатистики. Пробы растений предварительно разделяли на плоды, листья, стебли, корни и высушивали, затем измельчали. Поскольку в начале сентября формирование плодов у растений не было завершено и четко отделить их от засохших цветов не представлялось возможным, они рассматриваются вместе с цветами. Анализ растительного материала проводился в аналитическом центре Хабаровского института тектоники и геофизики им. Ю.А. Косыгина методом масс-спектроскопии с индуктивно связанной плазмой на приборе ISP-MS Elan DPC II PerkinElmer, аналитик Д.В. Авдеев.
Распределение молибдена в полыни Гмелина по участкам Шерловогорского горнорудного района дано на рисунке, а по органам растений в таблице. Сравнительный анализ полученных данных показал, что распределение молибдена по органам растений довольно неравномерно. Он однозначно свидетельствует о достаточно неоднородном распределении растений с контрастными содержаниями молибдена. Это связано как с особенностями его содержания в геологическом субстрате на различных участках, так и с особенностями захвата его растением.
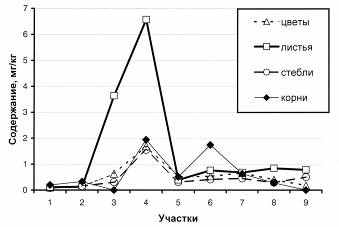
Рисунок. Среднее содержание молибдена в полыни Гмелина на территории
Шерловогорского рудного района.
По оси абсцисс указаны участки отбора проб: 1 – фоновый участок, 2 – сопка Мелехинская, 3 – жила Новая (дно карьера), 4 – жила Новая (бровка карьера), 5 – жила Поднебесных, 6 – жила Пятисотка, 7 – жила Лукаво-Золотая, 8 – Северный отвал, 9 – хвостохранилище.
Средние содержания молибдена в полыни Гмелина варьируют от менее 0.001 мг/кг в корнях до 6.57 в листьях, то есть, в пределах 4 порядков. Средние содержания в корнях составляют <0.001-1,94 мг/кг. Минимальное содержание молибдена в корнях полыни Гмелина обнаружено на участках, где отсутствует сформированный почвенный покров, например, в полотне карьера по жиле Новая и в почво-грунте хвостохранилища (<0.001 мг/кг). В этом же карьере на его кромке вскрыт почвенный слой мощностью до10 см, на котором развиты сообщества полыни Гмелина, в корнях которой обнаружены самые высокие (1,94 мг/кг) содержания молибдена. Подобное же развитие почв наблюдается и на участке Обвинская-Пятисотка, где среднее содержание молибдена в корнях полыни достигает 1.74 мг/кг. Существенный вклад в распределение молибдена в корнях полыни Гмелина вносит и содержание его в почвенном слое.
Содержание молибдена в стеблях полыни Гмелина находится в пределах 0.08-1,55 мг/кг и в целом существенно ниже, чем в корнях. Лишь для растений с участков Жила Новая (низ) и хвостохранилища, где в корнях его содержание было на пределе чувствительности метода (<0.001мг/кг), наблюдается его увеличение до 0.31 и 0.5 мг/кг, что, вероятно, связано с необходимостью захвата молибдена из почвы как жизненно необходимого микроэлемента. За редким исключением (Мелехинский отрог, Обвинская-Пятисотка), в листьях содержание молибдена возрастает, достигая абсолютного максимума (6,57 мг/кг) на участке Жила Новая (верхняя кромка карьера с хорошо развитым почвенным слоем).
Таблица
Средние содержания молибдена в органах полыни Гмелина
| Участки | Органы растений | |||
| Корни | Стебли | Листья | Цветы-плоды | |
| Фоновый | 0.2 | 0.08 | 0.11 | 0.1 |
| Мелехинский отрог | 0.33 | 0.15 | 0.13 | 0.11 |
| Поднебесных | 0.51 | 0.31 | 0.39 | 0.51 |
| Жила Новая, низ | <0.001 | 0.31 | 3.64 | 0.63 |
| Жила Новая, верх | 1.94 | 1.55 | 6.57 | 1.69 |
| Обвинская-Пятисотка | 1.74 | 0.41 | 0.76 | 0.53 |
| Лукаво-Золотая | 0.63 | 0.45 | 0.68 | 0.66 |
| Северный отвал | 0.28 | 0.29 | 0.84 | 0.4 |
| Хвостохранилще | <0.001 | 0.5 | 0.78 | 0.18 |
В цветах-плодах за единственным исключением (участок Поднебесных, где молибденит визуально наблюдается наиболее часто) содержание молибдена уменьшается кратно по сравнению с таковым в листьях для участков с минимальным содержанием в корнях. Данные таблицы и рисунка однозначно свидетельствуют о том, что наиболее оптимальное содержание молибдена в цветах-плодах полыни Гмелина находится в пределах 1мг/кг, а оптимально-максимальное > 0.4 < 0.7 мг/кг. Этот вывод можно обосновать тем, что при содержаниях в листьях менее 0.4 мг/кг (участок Поднебесных) в цветах-плодах оно возрастает до 0.51мг/кг, а при содержаниях в листьях более 0.7 мг/кг уменьшается до <0.7 мг/кг.
В целом, можно полагать, что молибден, являясь водным мигрантом, легко усваивается корневой системой полыни Гмелина. Но, не задерживаясь в ней, вместе с водой передвигаясь через стебли, не накапливаясь в проводящих тканях, он движется к листьям, где происходит фотосинтез, другие жизненно важные процессы формирования важнейших структурных элементов тканей, промоторов, катализаторов и ферментов с участием молибдена. Но содержания его в органах, обеспечивающих наследственность и чистоту вида (цветы-плоды) контролируются организмом полыни Гмелина.
Полученные выводы в части количественных показателей могут отличаться от полученных авторами и приведенных в настоящей работе, но принципиальные положения, вероятнее всего, сохранятся.
Формы передвижения молибдена в самих растениях неизвестны и требуют постановки специальных исследований.
Литература
1. Барабанов В. Ф. Геохимия: Уч. для вузов. – Л.: Недра, 1985. – 423 с.
2. Башкин В. Н., Касимов Н. С. Биогеохимия. – М.: Научный мир, 2001. – 648 с.
3. Юргенсон Г.А. Геохимия ландшафта. Учебное пособие для вузов. – Чита.: Изд-во ЗабГГПУ, 2005. – 152 с.
4. Юргенсон Г.А. Минеральное сырье Забайкалья. Учебное пособие для вузов.Ч.I. Черные и цветные металлы. – Чита: Поиск. 2006. – 256 с.
Boльфрам в полыни Гмелина на территории Шерловогогорского
горнорудного района
А.А. Смирнов, Г.А. Юргенсон
Институт природных ресурсов, экологии и криологии СО РАН, Чита, Россия, root@chinr.chita.su, inrec.sbras@mail.ru
TUNGSTEN IN THE GMELEN’S wormwood on the territory of
Sherlovogorskiy mining area
a.a. Smirnov , G.A. Yurgenson
Institute of Natural Resources, Ecology and Criology SB RAS, Chita, Russia, root@chinr.chita.su, inrec.sbras@mail.ru
The distribution of tungsten in the organs of Gmelen’s wormwood (Artemisia Gmelini) on the territory of Sherlovogorsk mining area is studied. It is revealed that concentration of tungsten in the organs of this plant depends on place of its growing.
Биологическая функция вольфрама считается во многом аналогичной молибдену, близкого к нему по особенностям координационной химии [1, 4]. Эти элементы в составе ферментативного комплекса катализируют главные метаболические реакции азотного цикла, участвуют в биологической трансформации соединений серы и углерода [10]. Тем не менее, замещение молибдена вольфрамом в простетической группе нитратредуктазы приводит к инактивированию этого фермента, что объясняется большей стабильностью соединений вольфрама (+6) [3, 5]. Кроме того, согласно [10], участие вольфрама в биохимических процессах определяется окислительно-восстановительным потенциалом среды. Вольфрам, в отличие от молибдена, активен в бескислородной обстановке, вследствие большей растворимости его соединений. В определенных условиях и концентрациях вольфрам является антагонистом молибдена, блокируя его активность [3, 5].
Биогеохимия вольфрама определяется его анионогенными свойствами и определенной степенью биофильности. По заключению А. И. Перельмана, [6] несмотря на то, что молибден и вольфрам относятся к элементам биологического захвата, а не поглощения, первый из них относится к среднему ряду и содержится в количествах от n до 0,n %. Второй входит в ряд элементов слабого и очень слабого захвата и содержится в количествах от 0,n до 0,00n %. При биосферном кларке вольфрама 0,00013 мг/кг [4,9], кларк в наземных растениях составляет 0,07 мг/кг, в морских – 0,035 мг/кг. Отмечаются высокие концентрации вольфрама в корнях растений при повышенных фоновых его содержаниях, свидетельствующее о вероятно легкой доступности его для растений [5]. В зонах рудной минерализации при фоновых содержаниях вольфрама 2,7 мг/кг токсичность его для растений проявляется при превышении этой величины в 18 раз, что свидетельствует об умеренной токсичности, а соответственно и возможности его концентрирования в растениях [4, 5]. При этом для вольфрама характерны довольно высокие показатели техногенности и технофильности [4]. В связи с этим на территориях с повышенным фоновым содержанием вольфрама высока вероятность включения значительных его концентраций в трофические цепи местных экосистем, в том числе и антропогенных. Данное обстоятельство, наряду с высокой токсичностью вольфрама для животных и человека, ведет к необходимости более глубокого исследования особенностей его биогеохимии в горно-рудных ландшафтах. Следует учесть повышение миграционной активности вольфрама от слабокислых до слабощелочных условий среды, характерных для сухостепных ландшафтов.
Цель данной работы заключается в определении биогеохимических особенностей аккумулирования вольфрама растительностью в зонах рудной минерализации на примере Шерловогорского горно-рудного месторождения.
Особенностью Шерловогорского горно-рудного района является полиэлементный состав достаточно контрастных геохимических аномалий, сопровождающих олово-бериллиево-висмут-вольфрамовое месторождение. Вольфрамовое оруднение связано с первым магматическим этапом эволюции рудномагматической системы. Основным минералом вольфрама является вольфрамит ((Fe,Mn)WO4), представленный преимущественно ферберитом, в котором железо преобладает над марганцем. В значительно меньшем количестве распространен шеелит (CaWO4).
Площадь рудного поля составляет более 10 км2. Она относится к подзоне сухих степей и представляет собой холмистую степь с ярко выраженным пологими формами рельефа. Относительные превышения высот достигают 200 м. Пади шириной 1.0-1.5 км характеризуются пологими и мягкими склонами. Сухие степи формируются в условиях скудного увлажнения. Среднегодовое количество осадков составляет 300 мм. Под сухой редкотравной степью в условиях сухого климата образуются каштановые почвы, представляющие собой переходный тип от чернозёмов умеренно-сухих степей к серозёмам пустынь. Содержание гумуса в аккумулятивном горизонте находится на низком уровне, а карбонаты кальция и магния содержатся во всём почвенном профиле, что связано с отсутствием промывного режима и сильной щебнистостью. Реакция почвенного раствора является слабощелочной (рН = 7.2-7.5). Субстратом степного ландшафта в районе Шерловой горы являются порфировидные граниты, гранит-порфиры интенсивно грейзенизированные в пределах этого рудного поля, аплиты, кварцевые порфиры, а также роговики. Все они обогащены бериллом, вольфрамитом, касситеритом, висмутином, арсенопиритом и топазом. Нередко они представляют собой бедные Be-W-Sn-Bi руды. В пределах олово-полиметаллического месторождения Сопка Большая располагается нерекультивированный карьер, склады добытой руды, а также хвостохранилище, которое в настоящее время полностью обезвожено и находится под воздействием ветровой эрозии.
В ландшафте развиты растительные сообщества горной лесостепи. На ненарушенных горными выработками территориях на северном склоне находится берёзово-осиновый лес с подлеском из боярышника, рододендрона, ерника. На склонах южных экспозиций располагается сухая низкотравная степь с доминированием полукустарничков. В степи на верхних частях склона преобладающими видами являются полынь холодная и полынь Гмелина.
Материалом для изучения поведения вольфрама в растительности являлась полынь Гмелина (Artemisia Gmelini). Пробы растения и почвогрунта отбирались впервой декаде сентября 2007 г. точечным методом на определенных участках месторождения. Участки сопки Мелехинской, Пятисотки, Лукаво-Золотой и Поднебесных характеризуют природный биогеохимический фон зоны рудной минерализации. Участки Жилы Новая, Северного отвала и хвостохранилища определяют биогеохимию техногенных ландшафтов. Кроме этого, были отобраны пробы вне зоны месторождения, отражающие региональный природный фон. В лабораторных условиях пробы высушивались, разделялись на органы и измельчались истирателем до порошкообразного состояния. Анализ растительного материала проводился в аналитическом центре Хабаровского института тектоники и геофизики им. Ю. А. Косыгина методом масс-спектрометрии с индуктивно связанной плазмой на приборе ISP-MS Elan DRC II PerkinElmer, аналитик Д. В. Авдеев.
Согласно ранее проведенным исследованиям [7,8] содержание вольфрама в почвенном горизонте выделенных участков следующее (г/т): сопка Мелехинская – 220; Лукаво-Золотая – 185; Поднебесных – 585; Жила Новая -180; Хвостохранилище –160.
Все минералы вольфрама слаборастворимы, в связи с чем характеризуются низкой мобильностью в гипергенных условиях. В ландшафтах вольфрам мигрирует преимущественно в виде вольфрамат-аниона, а также хелатных соединениях. Как отмечалось ранее, миграция вольфрама осуществляется в окислительно-нейтральной и щелочной средах. В кислой среде подвижность вольфрама ничтожно мала, что свидетельствует в пользу того, что растения извлекают преимущественно анионную форму вольфрама [2]. Таким образом, миграция вольфрама в ландшафтах, а следовательно, и его доступность растениям определяется щелочно-кислотным и окислительно-восстановительным равновесиями.
Значения рН почвенных горизонтов исследуемых участков колебались от слабокислых до слабощелочных (таблица). Эти значения рН характеризуют обстановки, благоприятные для перехода вольфрама в подвижные и легко доступные растениям формы миграции.
Как следует из таблицы, средние значения и среднеквадратичные отклонения по растениям в целом отличаются для различных участков. Максимальные содержания вольфрама в полыни Гмелина наблюдаются для участков: Лукаво-Золотая, Пятисотка, Жила Новая и Хвостохранилище. Минимумы характерны для Северного отвала, участка Поднебесных и Сопки Мелехинской. Хотя прямой зависимости поглощения вольфрама растением от значений рН почвенных растворов не наблюдается, тем не менее для минимальных значений рН Северного отвала характерны минимальные содержания вольфрама в полыни Гмелина. Растения с повышенными концентрациями вольфрама произрастают, как правило, на почвах с более высокими значениями рН. Таким образом отмечается заметное, хотя и не всегда прямое, влияние рН на аккумулирование вольфрама полынью Гмелина.
Таблица
Вариации содержаний вольфрама в растениях по местам отбора проб, мг/кг
| Место отбора | Статистические параметры в целом по растению | рН | ||
| n | 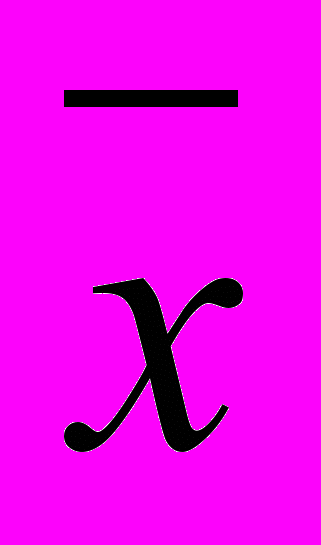 | σ | ||
| Фоновое содержание | 10 | 0,006 | 0,010 | 6 |
| Сопка Мелехинская | 10 | 0,083 | 0,112 | 6,47 |
| Пятисотка | 10 | 0,170 | 0,196 | 6,45 |
| Лукаво-Золотая | 10 | 0,208 | 0,114 | 6,84 |
| Поднебесных | 10 | 0,080 | 0,073 | 6,25 |
| Жила Новая | 10 | 0,150 | 0,155 | 7,6 |
| Северный отвал | 10 | 0,065 | 0,069 | 5,75 |
| Хвостохранилище | 10 | 0,113 | 0,064 | 7,94 |
Содержание вольфрама в растении варьирует как от места его произрастания, так и от органов самого растения (рисунок.).
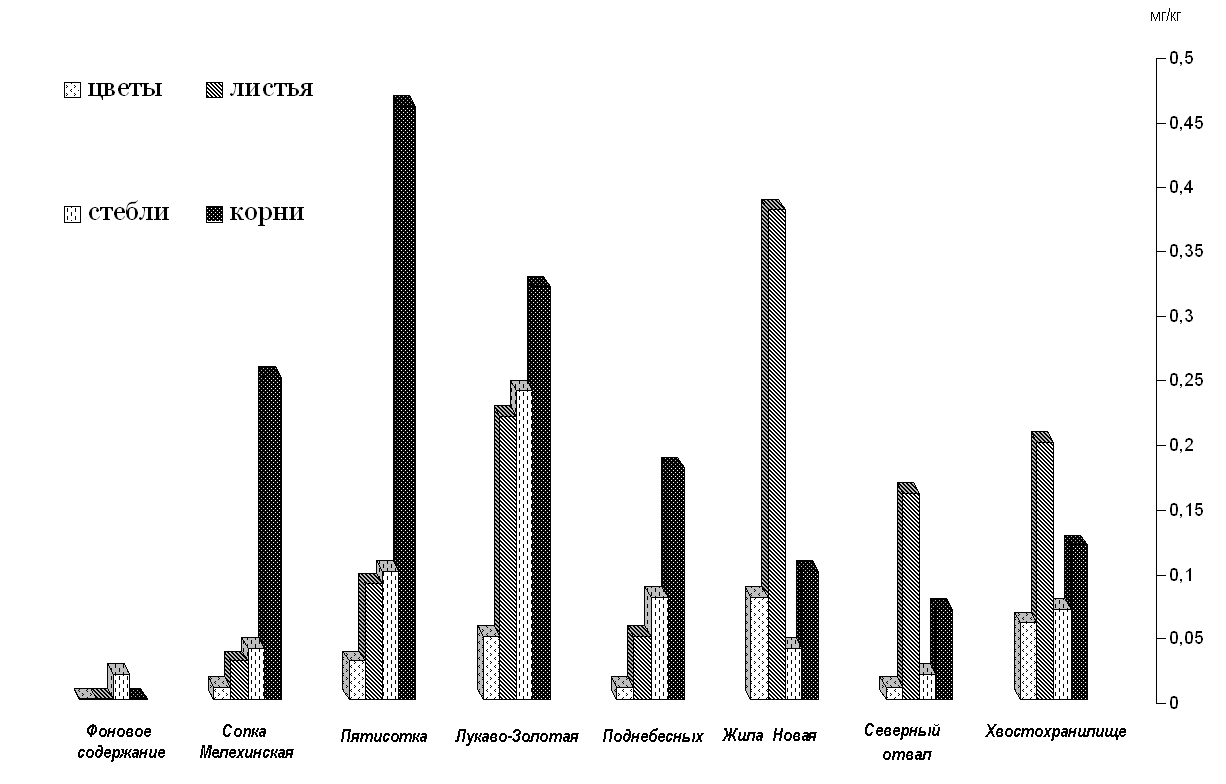
Рисунок. Распределение вольфрама по органам полыни Гмелина выделенных участков.
Характерны следующие изменения содержаний вольфрама в органах растений изученных участков: цветы (
= 0,031, σ = 0,029); листья ( = 0,141, σ = 0,125); стебли ( = 0,076, σ = 0,072); корни ( = 0,188, σ = 0,150). Высокие значения σ указывают на большой разброс содержаний вольфрама по всем органам растения на разных участках. Это свидетельствует о существенно различных условиях, а возможно и процессах поглощения вольфрама растениями.Как видно из рисунка, наибольшие содержания вольфрама характерны для корней и листьев. Для растений участков Сопка Мелехинская, Пятисотка, Лукаво-Золотая и Поднебесных характерны наибольшие содержания вольфрама в корнях. Для растений участков Жила Новая, Северный отвал и Хвостохранилище наибольшие концентрации вольфрама выявлены в листьях.
Повышенные содержания вольфрама в корнях можно объяснить, вероятно, так называемым метаболически промежуточным процессом [2]. В ходе этого процесса происходит поглощение ионов корнями так, что их концентрация в клеточной жидкости зачастую во много раз превышает концентрацию в почвенном растворе. Повышенные концентрации вольфрама в листьях можно представить как результат интенсивного перемещения ионов, поглощенных корнями, по направлению к листьям [2]. Относительно невысокие содержания вольфрама отмечаются в стеблях как органе транспортирующем растворенные минеральные вещества от корней к листьям и генеративным органам. Самые низкие содержания вольфрама наблюдаются в соцветиях и семенах. Это обстоятельство подтверждается более ранними нашими работами [6]. Вероятно, этот элемент является относительно токсичным для данного типа органов, и растения практически не аккумулирует и не вовлекает его в различного рода физиолого-биохимические процессы. Таким образом растение сохраняет дееспособность семян и чистоту вида.
Определенную роль в аккумулировании вольфрама различными органами играют и вариации рН (таблица), определяя направление и интенсивность протекания биогеохимических процессов.
Литература
1. Барабанов В. Ф. Геохимия: Уч. для вузов. – Л.: Недра, 1985. – 423 с.
2. Башкин В. Н., Касимов Н. С. Биогеохимия. – М.: Научный мир, 2001. – 648 с.
3. Битюцкий Н. П. Необходимые микроэлементы растений: Учебник. – С-Пб.: Изд-во ДЕАН, 2005. – 256 с.
4. Иванов В. В. Экологическая геохимия элементов: справоч. в 6 кн. – М.: Экология, 1997. – 576 с.
5. Кабата-Пендиас А., Пендиас Х. Микроэлементы в почвах и растениях: пер. с англ. – М.: Мир, 1989. – 439 с.
6. Перельман А.И. Геохимия ландшафта. - М.: Высшая школа, 1966. – 392 с.
7. Смирнов А.А. Биогеохимические особенности концентрации вольфрама в растениях Шерловогорского горно-рудного района (Читинская область) // Наш край. Сборник статей/ под ред. О. В. Корсуна: ЗабГГПУ – Чита, 2007. – С. 87-89
8. Смирнов А.А , Юргенсон Г.А , Солодухина М.А , Гудкова О.В. Первые данные о биогеохимических особенностях концентрации вольфрама в растениях Шерловогорского горно-рудного района //Минералогия и геохимия ландшафта горнорудных территорий. Современное минералообразование: Труды I Всероссийского симпозиума с международным участием «Минералогия и геохимия ландшафта горнорудных территорий» и VII Чтений памяти акад. А.Е. Ферсмана «Современное минералообразование» 7-10 ноября 2006 г. Россия. Чита, 2006. – С.33-41
9. Справочник по геохимии / Г.В. Войткевич, А.В. Кокин, А.Е. Мирошников, В.Г. Прохоров. – М.: Недра, 1990. – 480 с.
10. Федонкин М. А. Сужение геохимического базаси жизни и эвкариотизация биосферы: причинная связь.