
Удк 612. 45. 001. 6-092: 612. 7 Надпочечники при адаптации организма к двигательным нагрузкам и гипокинезии (экспериментально-морфологическое исследование) 14. 00. 02 анатомия человека
| Вид материала | Исследование |
- З. А. Зорина удк 612. 821. 6 Исследование, 666.77kb.
- Удк 616. 36+616. 61]: 612. 22-085. 275, 84.05kb.
- Венозная система сердца при хронической сердечной недостаточности (клинико-морфологическое, 551.53kb.
- Дк 616. 34-001. 4: 612. 014, 341.48kb.
- Вопросы к экзамену по дисциплине «спортивная физиология», 205.39kb.
- Рабочей учебной программы дисциплины б в. 05. «Физиология физической культуры», 10.21kb.
- Индивидуальной адаптации организма спортсменов к тренировочным нагрузкам в скоростно-силовых, 696.97kb.
- Оценки адаптации к физическим нагрузкам васильков А. А. Южно-Уральский государственный, 78.92kb.
- Психофизиологические основы адаптации и дезадаптации, 377.67kb.
- Конспект лекций Павлодар удк 591. 1+612](057. 8) Ббк 28. 673я7, 1073.37kb.
Надпочечники животных после небольших ИДДН (среднее время бега 27 мин) кровенаполнены, их весовой индекс незначительно увеличен (на 7,55 %). По сравнению с контролем отмечено увеличение ширины коркового (118,99 %, р<0,01) и мозгового (122,04 %, р<0,01) вещества (табл. 2). Имеет место тенденция к увеличению диаметра капилляров в корковом веществе, особенно в пучковой зоне. Композиция клубочковой зоны в надпочечнике у трети экспериментальных животных нарушена. Отчетливо выражена пучковость зоны, в основном за счет увеличения межклеточных пространств.
Размер ядер корковых эндокриноцитов в пучковой зоне достоверно увеличен (на 12,76 %, табл. 3). Диаметр ядер мозговых эндокриноцитов превысил значения в контрольной группе на 7,81 % (р<0,01).
Кроме увеличения размеров ядер выявлена тенденция к росту интенсивности гистоэнзиматических реакций на СДГ и 3β–ОН–СД. Оптическая плотность препаратов при постановке реакции на НАД*Н2 –ДГ увеличилась в клубочковой зоне на 47,69 % (р<0,01), в пучковой зоне – на 56,05 % (р<0,01).
Таблица 2
Ширина зон надпочечника (
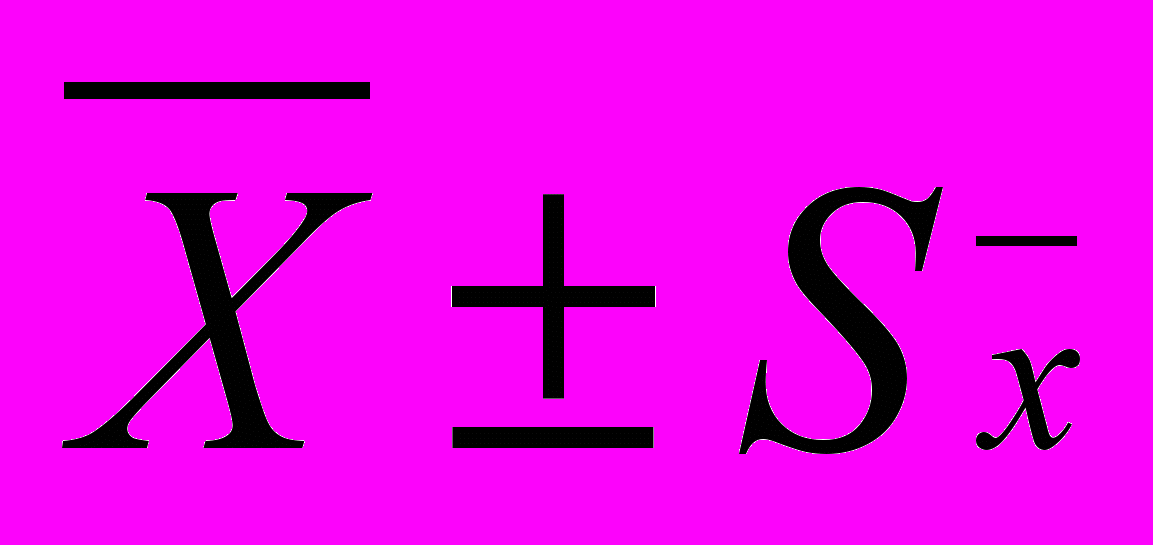 , мкм)
, мкм)| Группа | Клубочковая зона | Пучковая зона+ сетчатая зона | Мозговое вещество |
| Контрольная группа | 133,77±2,34 | 858,83±13,21 | 776,20±13,78 |
| Непродолжительный бег | 201,30±6,83** | 979,75±29,19** | 947,25±15,02** |
| Предельная двигательная нагрузка | 179,58±5,19** | 987,24±7,27** | 770,04±24,77 |
| Макроцикл тренировок | 182,40±3,93** | 886,51±17,98 | 832,44±22,37 |
| Три макроцикла тренировок | 148,60±4,70* | 1005,52±30,20* | 834,80±9,70* |
| Непродолжительная гипокинезия | 209,80±7,04** | 1029,13±28,68** | 903,45±16,93** |
| Длительная гипокинезия | 112,92±6,56* | 597,64±21,55** | 500,68±19,43** |
* – p<0,05, ** – р<0,01 (достоверные отличия от контроля)
Таблица 3
Диаметр ядер корковых и мозговых эндокриноцитов надпочечника (
, усл. ед.)| Группа | Клубочковая зона (длинный диаметр, короткий диаметр) | Пучковая зона+ сетчатая зона | Мозговое вещество |
| Контрольная группа | 79,63±1,49 37,54±0,56 | 51,88±0,61 | 57,07±0,48 |
| Непродолжительный бег | 81,77±0,92 39,06±0,43* | 58,50±1,06* | 61,53±0,62* |
| Предельная двигательная нагрузка | 93,41±1,52** 35,34±0,28* | 54,35±0,73* | 62,56±0,47** |
| Макроцикл тренировок | 84,05±1,53 38,77±0,52 | 55,07±1,02* | 60,71±0,70* |
| Три макроцикла тренировок | 92,98±1,26** 33,97±0,45** | 51,94±1,23 | 57,04±0,71 |
| Непродолжительная гипокинезия | 99,28±0,98** 38,86±0,56 | 54,67±0,74* | 61,88±0,51** |
| Длительная гипокинезия | 94,46±2,73** 35,51±0,56** | 52,85±0,67 | 57,62±0,73 |
* – p<0,05, ** – р<0,01 (достоверные отличия от контроля)
Полученные результаты свидетельствуют о росте энергетической и стероидопродуцирующей функции корковых эндокриноцитов (Gemell R.T., Laychock S.G., Rubin R.P., 1977; Hall P.F., 1984; Bomsel M. et al., 1986; Labrie F. et al., 1992; Feuilloley M., Vaurdy H., 1997).
В мозговых эндокриноцитах активность НАД*Н2 –ДГ высокая, на 41,05 % (р<0,01) превышала соответствующий показатель в контрольной группе, что является признаком увеличения окислительно-восстановительного потенциала клеток мозгового вещества.
Определено достоверное снижение интенсивности реакций на щелочную фосфатазу в корковой зоне на 25,90 %, в пучковой зоне – на 19,88 % и на 9,50 % – в мозговом веществе.
В корковом веществе на электронно-микроскопическом уровне выявлено просветление матрикса митохондрий, уменьшение количества крист, стертость мембран, появление митохондрий открытого типа, увеличение контактов митохондрий и липидных капель, уменьшение как количества, так и относительного объема последних, просветление липидных капель, иногда отсутствие в них содержимого, расширение цистерн эндоплазматического ретикулума, что, в совокупности, является признаком высокой функциональной активности (Basset J.R., Pollard I., 1980; Feuilloley M., Vaurdy H., 1997). В мозговых эндокриноцитах наблюдались сходные изменения. Матрикс части митохондрий просветлен, тип крист пластинчатый, их количество уменьшено. Гранулы катехоламинов объединены в различные по форме структуры. Одни из них образуют крупные осмиофильные глыбки, полностью заполняющие пространство. Другие имеют свободное пространство разной величины между содержимым и окружающей мембраной. Третьи не имеют формы сплошного конгломерата, а представлены ровными осмиофильными гранулами. Четвертые имеют конгломераты различной формы и величины (мелкие и крупные), заключенные в одной оболочке. Это, по мнению A.D. Smith (1973), E.A. Richter (1984), отражает разные стадии выведения секрета. О выходе содержимого эндокриноцитов в кровь свидетельствует наличие секреторных гранул в просвете капилляров.
Корреляционный анализ выявил функциональные связи между надпочечником и такими показателями как вес животного, систолический показатель (СП), кислородный пульс (КП), частота сердечных сокращений (ЧСС), частота дыхания (ЧД), потребление кислорода (О2) (рис. 1).
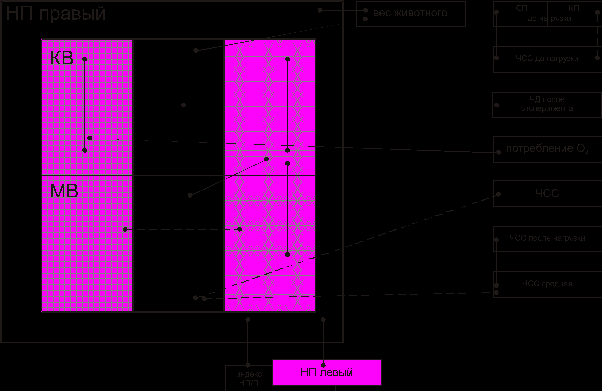
Рис. 1. Взаимодействие между элементами надпочечника при действии двигательных нагрузок в виде бега, в среднем в течение 27 минут:
——— положительные связи,
-------- отрицательные связи,
 показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессы
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыВ надпочечнике во взаимодействие были вовлечены элементы, характеризующие энергетическую, синтетическую активность клеток, обменные процессы как в корковом, так и в мозговом веществе. Соотношение количества связей в мозговом веществе к связям в коре надпочечника составило 0,70.
В поле отличий морфофункциональных показателей корковое и мозговое вещество располагаются по диагонали, имея одинаковый процент положительных и отрицательных значений.
Таким образом, после действия непродолжительной ИДДН (бег в среднем 27 мин) в надпочечниках имеют место адаптивные реакции как в корковом, так и в мозговом веществе, направленные на сохранение функциональной активности железы при некотором напряжении в системе, обеспечивающей обменные процессы.
Однократная предельная двигательная нагрузка (время бега в среднем 193 мин) привела к изменениям морфологического состояния надпочечников. Весовой индекс надпочечник/почка превысил на 14,81 % (р<0,05) показатель контрольной группы. Зоны коркового вещества расширены: ширина клубочковой зоны на 34,25 % (р<0,01), пучковой зоны на 14,95 % (р<0,01) больше, чем в группе контроля (табл. 2). Размеры ядер увеличились (табл. 3) в клубочковой зоне на 17,31 % (длинный диаметр), в пучковой зоне – на 4,76 % и в мозговом веществе – на 9,62 %.
Отмечена высокая интенсивность реакции ферментов митохондриального энергообеспечения (НАД*Н2 –ДГ), она составила в клубочковой зоне 128,30 %, в пучковой зоне 126,47 % от контрольных цифр. Оптическая плотность препаратов при постановке реакции на фермент специфического синтеза гормонов 3β–ОН–СД превысила значения в контрольной группе (в клубочковой зоне на 8,95 %, в пучковой зоне на 2,00 %). Уменьшилась оптическая плотность продукта реакции щелочной фосфатазы в клубочковой, пучковой зонах и в мозговом веществе (на 20,22 %, 17,91 %, 14,46 %).
Диаметр капилляров по сравнению с контролем (табл. 4) увеличен в пучковой зоне на 14,68% (р<0,05). Расстояние между капиллярами не отличалось от показателей контрольной группы. Стенка капилляров коры надпочечников отечная, ядра эндотелия уплотнены, выбухают в просвет.
Ретикулярные волокна извитые, интенсивно воспринимают серебро, относительная площадь, занимаемая ими, увеличена во всех зонах надпочечника. Обнаружены признаки, свидетельствующие о повышении функциональной активности эндокриноцитов: просветление матрикса митохондрий; увеличение объема, занимаемого митохондриями, на 21,04 %; расположение митохондрий в непосредственной близости от липидных капель, относительный объем которых составил 66,33 % от контрольных значений. В мозговом веществе интенсифицируются аэробные процессы. Оптическая плотность препаратов при выявлении НАД*Н2 –ДГ достоверно (р<0,01) выше, чем в контрольной группе (на 16,82 %).
Таблица 4
Диаметр капилляров надпочечника (
, мкм)| Группа | Клубочковая зона | Пучковая зона+ сетчатая зона | Мозговое вещество |
| Контрольная группа | 6,35±0,24 | 5,86±0,19 | 6,80±0,20 |
| Непродолжительный бег | 6,53±0,21 | 6,50±0,21 | 7,47±0,25 |
| Предельная двигательная нагрузка | 6,67±0,28 | 6,72±0,21* | 6,97±0,33 |
| Макроцикл тренировок | 5,98±0,10 | 7,34±0,25* | 7,88±0,29 |
| Три макроцикла тренировок | 8,64±0,36** | 8,58±0,42** | 8,59±0,41** |
| Непродолжительная гипокинезия | 7,02±0,32 | 7,39±0,35* | 9,07±0,37* |
| Длительная гипокинезия | 12,70±0,29** | 14,36±0,50** | 14,57±0,34** |
* – p<0,05, ** – р<0,01 (достоверные отличия от контроля)
Состояние капиллярного русла мозгового вещества такое же, как и в корковом веществе. В эндокриноцитах мозгового вещества количество митохондрий на единицу площади 18,45±1,45, что на 49,51 % больше, чем в контрольной группе (р<0,05), увеличен относительный объем, занимаемый секреторными гранулами (на 5,33 %) и их количество на стандартной площади среза (на 11,52 %), отмечено резкое уменьшение доли полупустых гранул, при этом доля пустых и полных возросла на 75,53 % и 85,94 % соответственно. Такой состав включений, по S. Shionoya (1963), определяется различными стадиями выведения секрета. К этому нужно добавить феномен появления участков секвестрирующей цитоплазмы с мелким гранулярным содержимым, что отражает один из возможных путей выведения секрета.
Корреляционный анализ выявил наличие 8 связей в пределах органа между элементами, обеспечивающими энергетический потенциал клетки, синтетические и обменные процессы как в корковом, так и в мозговом веществе. Соотношение количества связей между элементами мозгового вещества и элементами клубочковой и пучковой зон характеризуется коэффициентом 0,70. В контрольной группе этот коэффициент составил 0,60. Увеличение на 16,67 % связано с ростом количества связей в мозговом веществе (рис. 2). Обращает внимание появление достоверной корреляционной связи между временем бега животного и шириной пучковой зоны, расстоянием между капиллярами в мозговом веществе.
Изучение корреляционных связей по уровням организации НП показало их присутствие на всех изучаемых уровнях. Большинство связей объединяло элементы в пределах одного уровня (горизонтальные связи).
Информационный анализ определил изменения коэффициента относительной энтропии (h) по сравнению с контрольной группой. В большей степени состояние напряжения возникло между элементами мозгового вещества (h.=0,920 против h.=0,822 в контроле). Коэффициент энтропии в мозговом веществе превышал контрольные значения по таким показателям как расстояние между капиллярами, ширина мозгового вещества.
Таким образом, действие предельной физической нагрузки вызывало повышение активности структурных элементов всех зон надпочечника. В большей степени активизируется корковое вещество.
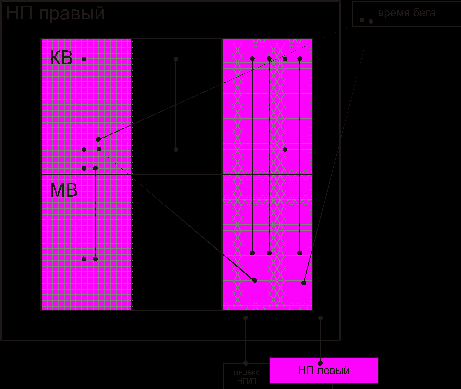
Рис. 2. Взаимодействие между элементами надпочечника при действии на организм предельных двигательных нагрузок:
——— положительные связи,
-------- отрицательные связи,
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыПроведенный анализ индивидуальных особенностей и морфофункционального единства по С.Б. Стефанову (1974) позволил выделить две подгруппы собак (рис. 3) с различными типами реакций структурных элементов надпочечника при действии предельной двигательной нагрузки. В надпочечниках собак первой подгруппы весовой индекс надпочечник/почка достоверно увеличен на 17,61 % (р<0,05), корковое вещество хорошо структурировано, имеет наибольшие размеры. Ширина клубочковой и пучковой зон на 13,85 %, и на 22,01 % (p<0,01) превышает соответствующие показатели контрольной группы. Просвет капилляров широкий, расстояние между ними невелико. Интенсивность реакции на СДГ в клубочковой зоне на 16,27 % и в пучковой зоне на 14,32 % выше, чем в контроле. Достоверно увеличена оптическая плотность препаратов при выявлении НАД*Н2 –ДГ в корковом веществе (на 28,41 % в КЗ, на 27,54 % в ПЗ). Активность щелочной фосфатазы в клубочковой зоне составила 80,66 %, в пучковой зоне 83,98 % от контрольных цифр. Диаметр ядер клубочковой зоны составил 115,41 % (р<0,01) от контроля. На субклеточном уровне обнаружены изменения митохондрий, характеризующиеся появлением большого числа митохондрий с размытыми мембранами крист и наружной оболочки. Расширенные цистерны эндоплазматической сети расположены вокруг липидных капель (ЛК) с неоднородной плотностью. Объем ЛК уменьшен. В совокупности с данными гистоэнзиматического исследования это свидетельствует об увеличении окислительно-восстановительного потенциала в клетках коры и усилении стероидогенеза (McCarty R., 1983).

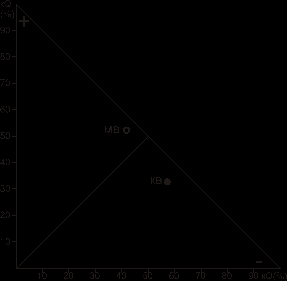
а б
Рис. 3. Поле отличий морфофункциональных показателей надпочечников собак
1 подгруппы (a) и 2 подгруппы (б) при действии предельных двигательных нагрузок (МВ – мозговое вещество, КВ – корковое вещество)
Для мозгового вещества надпочечников первой подгруппы собак характерно наличие неравномерно суженных синусоидных капилляров, расположенных на большом расстоянии друг от друга. Показатель энергетических процессов клеток НАД*Н2 –ДГ ниже среднего в группе. На электроннограммах мозгового вещества собак определено не только уменьшение количества, но и объема митохондрий на 24,95 % и 36,31 % соответственно. Количество гранул катехоламинов невелико.
В поле отличий морфофункциональных параметров корковое вещество собак данной подгруппы занимает зону положительных показателей (KQ+), а мозговое, наоборот, отрицательных (KQ-).
Животные первой подгруппы показали максимальное время бега (255,62±27,24 мин).
Для надпочечников собак первой подгруппы характерна значительная активность элементов коры и низкая активность мозгового вещества. По данным литературы (Виру А.А., 1977, 1980; Gauthier С. et al., 1983), это соответствует о высокой концентрации стероидов и низком уровне катехоламинов в крови, что определяет особенности энергообеспечения мышечной деятельности. Высокая активность эндокриноцитов клубочковой и пучковой зон надпочечника приводит к усилению синтеза глюко- и минералокортикоидов, нормализуя водно-электролитный баланс. Глюкокортикоиды мобилизуют аминокислоты для синтеза белков и переаминирования (Виру А.А., 1980), вызывают повышение синтеза гликогена в печени. Мы отмечали увеличение содержания гликогена печени в этой подгруппе до 30 % от контроля, наряду со снижением концентрации молочной кислоты и гипогликемией. Вероятно, у животных данной подгруппы наблюдалось усиление пластических и энергетических процессов, сохранение водно-электролитного баланса, что явилось необходимым условием для выполнения длительной, свыше 200 минут, мышечной нагрузки.
Животные второй подгруппы показали минимальное время бега (66,67±14,15 мин). В этой группе выявлена наибольшая степень активации структурных элементов в мозговом веществе. Ширина мозгового вещества достоверно увеличена (на 12,00 %), площадь ретикулярного каркаса незначительна, диаметр синусоидных капилляров максимальный. Размеры ядер мозговых эндокриоцитов составляют 116,87 % (р<0,01) от контроля. Интенсивность реакции на НАД*Н2 –ДГ достоверно выше (на 26,78 %), чем в контрольной группе. В митохондриях МВ имеются участки просветления матрикса. Относительный объем митохондрий и их количество значительны. Цистерны эндоплазматической сети расширены, много гранул катехоламинов: на 11,55 % больше, чем у собак первой подгруппы. В корковом веществе НП животных второй подгруппы резко увеличен относительный объем, занимаемый ретикулярными волокнами – на 57,47 % в клубочковой зоне (р<0,01) и на 56,41 % в пучковой зоне (р<0,01). Нет достоверных изменений размеров ядер, интенсивность реакции на СДГ, 3–ОН–СД не отличается от средних в группе. Все это, наряду с неравномерным просветом капилляров, увеличением межкапиллярного пространства, уменьшением относительного объема и количества митохондрий, просветлением цитоплазмы, может свидетельствовать о нарушении процессов продукции стероидных гормонов.
Анализ морфофункционального единства показал, что мозговое вещество надпочечников собак с минимальным временем бега занимает зону положительных показателей, а корковое вещество расположено в зоне отрицательных показателей (рис. 3).
Активация элементов мозгового вещества НП и невысокая функциональная активность коркового вещества, вероятно, определили преобладающее влияние катехоламинов при выполнении физической нагрузки (Виру А.А., 1980), которые способствовали энергообеспечению бега за счет усиления транспорта кислорода и увеличения гликогенолиза и липолиза. Это подтверждается относительным уменьшением содержания гликогена в печени – на 20 % по сравнению с первой подгруппой, повышением уровня молочной кислоты в крови. Такие признаки отражают преобладание тонуса симпатической нервной системы (Кендыш И.В., 1985).
При многократных двигательных нагрузках (бег со скоростью 15 км/ч от 11 минут в первые дни тренировок до одного часа и более в сутки в конце опыта, всего 16-20 нагрузок) – первый макроцикл тренировки – происходят адаптационные перестройки в надпочечниках. Наиболее выраженные изменения выявлены в корковом веществе. Сосуды и капилляры надпочечника наполнены кровью. Ширина коркового вещества на 7,69 % (р<0,01) больше этого показателя в контроле (табл. 2). Размеры ядер в пучковой зоне на 6,15 % (р<0,01) превышают контрольные значения (табл. 3). Схожие изменения получены S.K. Droste и соавт. (2003) при тренировке мышей в течение одного месяца. Клубочковая зона состоит из многочисленных петель и клубочков, форма которых разнообразна (округлая, удлиненная, неправильная). Количество клубочков в поле зрения в среднем 2-3, колебания составляют от 1-2 до 3-5. Обращает внимание наличие клубочков, сформированных из небольших клеток округлой формы, содержащих мелкие ядра. Редко встречаются клубочки с клетками, имеющими темную цитоплазму, равномерно окрашенную, определяются «безъядерные зоны».
При постановке гистоэнзиматических реакций в корковом веществе надпочечников было отмечено повышение интенсивности реакций и увеличение площади, занимаемой ферментами, регулирующими энергетические и синтетические процессы в клетках (табл. 5). Оптическая плотность препаратов при определении сукцинатдегидрогеназы в клубочковой и пучковой зонах на 28,53 % (р<0,01) и 20,22 % (р<0,01) больше соответствующих показателей в контрольной группе животных. Интенсивность реакций на фермент, катализирующий процессы стероидогенеза (3β–ОН–СД), в клубочковой зоне – на 19,84 % (р<0,01), в пучковой на 14,11 % (р<0,01) выше, чем в контроле. Диаметр капилляров (табл. 4) в пучковой зоне составил 125,26 % от контроля (р<0,01). Выявлено увеличение расстояния между капиллярами в клубочковой и пучковой зонах (на 23,32 % и на 32,71 % соответственно, р<0,01) и снижение активности реакции на щелочную фосфатазу в корковом веществе надпочечника на 23,41 % (р<0,01), что может свидетельствовать о некотором напряжении в системе транспортных и обменных процессов.
Таблица 5
Оптическая плотность препаратов (усл. ед.) при постановке гистоэнзиматических реакций в надпочечниках
при действии многократных нагрузок (один макроцикл тренировок),
| Зоны | Показатели | Контроль | Опыт |
| КЗ | СДГ | 0,848±0,034 | 1,090±0,028** |
| НАД*Н2–ДГ | 0,887±0,031 | 1,226±0,045** | |
| 3–ОН–СД | 0,771±0,024 | 0,924±0,024** | |
| ЩФ | 1,251±0,036 | 0,959±0,032** | |
| ПЗ | СДГ | 0,831±0,029 | 0,999±0,021** |
| НАД*Н2–ДГ | 0,835±0,031 | 1,179±0,044** | |
| 3–ОН–СД | 0,801±0,024 | 0,914±0,021** | |
| ЩФ | 1,167±0,026 | 0,893±0,029** | |
| МВ | НАД*Н2–ДГ | 0,743±0,029 | 0,816±0,028 |
| ЩФ | 0,906±0,026 | 0,812±0,023** |
** – p<0,01 (достоверные отличия от контроля)
На электроннограммах выявлены признаки, характеризующие активацию структурных элементов коркового вещества надпочечника. Цистерны эндоплазматического ретикулума расширены вблизи липосом и ядер. Количество липидных капель уменьшено, плотность их неоднородная, имеются участки просветления, их можно обнаружить в просвете кровеносных капилляров. Липидные капли образуют многочисленные контакты с митохондриями.
Корреляционный анализ в экспериментальной группе показал наличие большего количества связей на клеточном уровне. Все связи горизонтальные и положительные. Их количество, по сравнению с контрольной группой, увеличено в 2 раза. Были выявлены достоверные связи между временем бега и приростом работоспособности, между весом надпочечника и весом животного (рис.4).
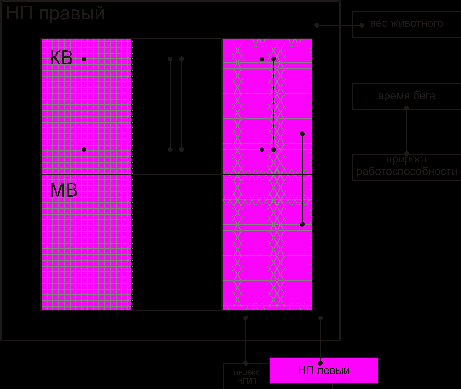
Рис. 4. Взаимодействие между элементами надпочечника при действии многократных нагрузок (один макроцикл тренировок):
——— положительные связи,
-------- отрицательные связи,
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыКорреляционные взаимосвязи между элементами надпочечника в этой группе отличались от контроля и от серии животных, получивших однократную индивидуальную дозированную нагрузку. Соотношение функциональных связей между элементами мозгового и коркового вещества составило 0,20. Это свидетельствует о превалировании связей в корковом веществе надпочечников: на одну связь в мозговом веществе приходится пять связей в корковом. В коре надпочечников во взаимодействие вовлечены элементы, характеризующие энергетические, синтетические и обменные процессы (СДГ, НАД*Н2 –ДГ, 3–ОН–СД, ЩФ и т.д.).
В поле отличий корковое вещество занимает зону преимущественно положительных показателей (KQ+), а мозговое вещество – зону отрицательных показателей (KQ-).
Коэффициент относительной энтропии в клубочковой зоне составил, в среднем, h=0,842, в пучковой зоне – h=0,867 и в мозговом веществе – h=0,868. Наибольшие показатели относительной энтропии были определены в мозговом веществе (ширина зоны – h=0,947, диаметр капилляров – h=0,891, расстояние между капиллярами – h=0,906), что свидетельствует о перенапряжении в системе, обеспечивающей обменные процессы (Кадыров Х.К., Антомонов Ю.Г., 1974).
При многократных двигательных нагрузках – чередовании трех макроциклов мышечных нагрузок – животные получили от 62 до 78 тренировок. Увеличение времени бега к концу эксперимента было, в среднем, на 654,50 %.
Исследования показали, что ширина коркового вещества (табл. 2) увеличена на 16,27 % (р<0,01). В клубочковой зоне ее увеличение произошло за счет роста количества рядов клубочков. Архитектоника клубочков была нарушена, границы между соседними клубочками часто стерты. Капсула надпочечника отдавала внутрь органа широкие соединительнотканные трабекулы, вдоль которых клубочки врастали в пучковую зону.
Между корковыми эндокриноцитами были заметны широкие прослойки соединительной ткани, содержавшие большое количество клеточных элементов, за счет которых происходила пролиферация клеток коры (Chamoux E., Otis M., Gallo-Payet N., 2005).
Размеры ядер корковых эндокриноцитов клубочковой зоны (табл. 3) увеличены (р<0,01). Длинный диаметр на 16,77 % превышал таковой показатель контрольной группы (p<0,01), короткий диаметр уменьшен на 9,51 %. В пучковой зоне размер ядер не отличался от аналогичного показателя контроля. Корковые эндокриноциты пучковой зоны имели признаки повышения внутриклеточных метаболических процессов в виде изменений ядра (наличие выраженного ядрышка, расширение перинуклеарного пространства и ядерных пор) и митохондрий (очаговые просветления матрикса, отек и гомогенизация крист, появление тубулярных и пластинчатых крист), гипертрофии комплекса Гольджи, уменьшении количества липидных капель (на 40,41 %, р<0,01). Выявлена тенденция к уменьшению размеров митохондрии, относительный объем органеллы снижается на 17,01 %. При этом количество митохондрий на стандартной площади среза снижается на 42,47 %. В то же время отмечена тенденция к снижению числа контактов между митохондриями и липидными каплями. В единичных случаях наблюдались дистрофические изменения паренхимы.
Показатели состояния капиллярного звена кровеносного русла представлены в таблице 4. Диаметр капилляров клубочковой зоны составил 136,06 %, пучковой зоны – 146,42 % (р<0,01), мозгового вещества – 126,32 % от контрольной группы (р<0,001).
В эндотелиоцитах капилляров пучковой зоны обнаружены признаки усиленного транскапиллярного обмена, что подтверждается наличием органелл и фрагментов цитоплазмы корковых эндокриноцитов в просвете капилляров мозгового вещества. Расстояние между капиллярами пучковой зоны уменьшено, что свидетельствует об улучшении условий кровоснабжения паренхимы.
Изменения в обеих популяциях мозговых эндокриноцитов не столь выражены. Отдельные клетки имели признаки внутриклеточного отека, встречалось некоторое количество некротически измененных клеток. Размеры ядер мозговых эндокриноцитов не отличались от ядер контрольной группы. Отмечено достоверное снижение количества митохондрий на стандартной площади на 34,12 % при наличии тенденции к уменьшению их размеров на 11,51 %. Изменения митохондрий в виде отека, очаговых просветлений матрикса и деструкции крист свидетельствуют об активации энергообеспечивающей функции клеток. Относительный объем секреторных гранул не отличался от уровня контрольной группы. Выявлено снижение доли полупустых гранул, значительное повышение относительной доли и абсолютного количества полных секреторных гранул (табл. 6).
Отмечено значительное полнокровие капилляров внутренних участков надпочечников, особенно мозгового вещества. В мозговом веществе выявлено расширение кровеносных капилляров, что в целом свидетельствует об улучшении условий трофического обеспечения эндокриноцитов и секреции гормонов в кровеносное русло. В просвете крупных синусоидов регистрируются многочисленные секреторные гранулы. Изменение соотношения типов секреторных гранул говорит о накоплении катехаламинов в мозговых эндокриноцитах, что может явиться следствием пониженного уровня процессов секреции гормонов. Однако, учитывая отмеченные морфологические признаки активного секреторного процесса со стороны капиллярного звена кровеносного русла, можно говорить об экономизации функционирования секреторного эпителия и увеличении «резервного фонда» катехоламинов в мозговом веществе надпочечников.
Таблица 6
Показатели состояния мозговых эндокриноцитов надпочечников при действии многократных двигательных нагрузок (три макроцикла тренировок),
| Показатели | Контроль | Опыт |
| Относительный объем митохондрий, % | 6,95±0,95 | 6,15±0,61 |
| Количество митохондрий, шт. на 100 мкм2 | 12,34±1,38 | 8,13±0,84* |
| Относительное содержание пустых гранул, % | 5,15±0,80 | 7,21±1,46 |
| Количество пустых гранул, шт. на 100 мкм2 | 10,11±1,56 | 13,40±2,71 |
| Относительное содержание полупустых гранул, % | 79,99±3,14 | 65,37±3,77* |
| Количество полупустых гранул, шт. на 100 мкм2 | 157,11±6,18 | 121,55±7,02 |
| Относительное содержание полных гранул, % | 14,86±2,80 | 27,42±3,85* |
| Количество полных гранул, шт. на 100 мкм2 | 29,19±5,51 | 50,99±7,15* |
* – р<0,05 (достоверные отличия от контроля)
Корреляционный анализ выявил наличие связей на органном и организменном уровне. Показано взаимодействие отдельных элементов надпочечника и таких показателей как время тренировок на 1, 2 и 3 макроциклах, вес животного (рис. 5).
Соотношение корреляционных связей в мозговом веществе и корковом веществе составило, как и в предыдущей группе животных, получивших многократные двигательные нагрузки – 0,20 (на 5 связей коркового вещества приходится 1 связь в мозговом веществе). В коре НП функциональные связи в большей степени имели место в системе обменных процессов.
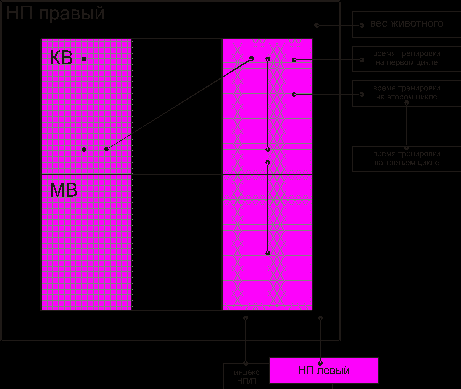
Рис. 5. Взаимодействие между элементами надпочечника при действии многократных двигательных нагрузок (три макроцикла тренировок):
——— положительные связи
-------- отрицательные связи
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыКорковое вещество в поле отличий по С.Б. Стефанову занимает зону положительных показателей (KQ+), а мозговое вещество – зону отрицательных показателей (KQ-).
Информационный анализ определил напряжение элементов коркового и мозгового вещества надпочечников (коэффициент относительной энтропии в клубочковой зоне и мозговом веществе на 3,00 %, в пучковой зоне на 9,81 % выше, чем в контроле).
Непродолжительное ограничение двигательной активности (около 30-и суток) вызывало изменения морфологического статуса надпочечника. Надпочечники кровенаполнены, по сравнению с контролем отмечено увеличение весового индекса надпочечник/почка справа на 22,22 %, слева на 34,62 %. Ширина коркового вещества увеличена на 24,81 %, мозгового вещества – на 16,39 % (табл. 2). Размеры ядер эндокриноцитов достоверно превышали (табл. 3) значения контрольной группы (99,28±0,98 усл. ед. в клубочковой зоне, 54,67±0,74 усл. ед. в пучковой зоне и 61,88±0,51 усл. ед. в мозговом веществе против 79,63±1,49 усл. ед., 51,88±0,61 усл. ед. и 57,07±0,48 усл. ед. соответственно).
Отмечен рост интенсивности реакций на НАД*Н2 –ДГ на 67,98 % в клубочковой зоне и на 44,67 % в пучковой зоне (табл. 7). Значительно увеличена оптическая плотность препаратов при постановке реакции на фермент, катализирующий стероидогенез (3β–ОН–СД), в клубочковой зоне – на 87,88 %, что свидетельствует о высокой активности клеток коркового вещества надпочечников.
Капилляры в НП широкие (табл. 4), особенно в пучковой зоне и в мозговом веществе, где диаметр достоверно превышал значения контроля на 26,11 % и на 33,38 % соответственно.
Изменения на ультраструктурном уровне также свидетельствовали о признаках активации коркового и мозгового вещества надпочечников: наличие хорошо выраженных ядрышек, гипертрофия комплекса Гольджи, расширение цистерн эндоплазматического ретикулума. Уменьшено количество липидных капель на единицу площади на 30,14%. Большинство митохондрий имели просветленный матрикс, форма и размеры митохондрий варьировали – от микро- до макромитохондрий. Наличие микромитохондрий служит признаком ускорения процессов новообразования данных органелл. Появление в цитоплазме клеток гигантских митохондрий связывают с состоянием гиперфункции коркового вещества надпочечников (Зуфаров К.А. и соавт., 1982). Выявлены признаки слияния липидных капель, характер их распределения в цитоплазме не изменился.
Таблица 7
Оптическая плотность препаратов (усл. ед.) при постановке гистоэнзиматических реакций в надпочечнике после действия непродолжительной гипокинезии,
-
Зоны
Ферменты
Контроль
Опыт
КЗ
СДГ
0,848±0,034
0,717±0,055
НАД*Н2 –ДГ
0,887±0,031
1,490±0,090**
3β–ОН–СД
0,771±0,024
1,448±0,149**
ЩФ
1,251±0,036
1,372±0,143
ПЗ
СДГ
0,831±0,029
0,462±0,028**
НАД*Н2 –ДГ
0,835±0,031
1,208±0,117**
3β–ОН–СД
0,801±0,024
1,008±0,125
ЩФ
1,167±0,026
0,974±0,130
МВ
НАД*Н2 –ДГ
0,743±0,029
0,625±0,054
ЩФ
0,906±0,026
0,916±0,052
** – р<0,01 (достоверные отличия от контроля)
Обращает внимание, что при уменьшении количества контактов митохондрий и липидных капель (на 43,00 %) увеличилась их протяженность за счет изменения формы, структуры и уплощенности. Сохранение значительного количества протяженных контактов между митохондриями и липидными каплями можно расценивать как компенсаторную реакцию, направленную на поддержание адекватного уровня функционирования корковых эндокриноцитов пучковой зоны в условиях гипокинезии.
В мозговых эндокриноцитах (табл. 8) достоверно увеличена доля «пустых» секреторных гранул (на 184,23 % в эпинефроцитах и на 158,91 % в норэпинефроцитах), обнаружен массированный выход гранул катехоламинов в кровеносное русло и расширение цистерн эндоплазматической сети.
Нахождение липосом корковых эндокриноцитов в просвете кровеносных капилляров мозгового вещества предполагает наличие интракапиллярного транспорта фрагментов секреторных клеток (Пугачев М.К., 1983) в условии гиперфункции коры надпочечников (Савина Е.А. и соавт., 1980).
Таблица 8
Соотношение типов секреторных гранул в мозговых эндокриноцитах надпочечников после действия непродолжительной гипокинезии,
| Тип гранул | Тип клеток | Относительное содержание гранул, % | |
| Контроль | Опыт | ||
| Пустые гранулы | эпинефроциты | 4,63±1,03 | 13,16±1,57* |
| норэпинефроциты | 5,67±0,97 | 14,68±1,15* | |
| Полупустые гранулы | эпинефроциты | 78,14±4,39 | 77,38±1,94 |
| норэпинефроциты | 81,84±2,46 | 73,15±1,45* | |
| Полные гранулы | эпинефроциты | 17,23±4,21 | 9,45±0,79 |
| норэпинефроциты | 12,49±2,07 | 12,16±0,86 | |
* – р<0,05 (достоверные отличия от контроля)
В мозговом веществе число патологически измененных клеток невелико. Отмеченные в обеих популяциях мозговых эндокриноцитов изменения в виде гипертрофии комплекса Гольджи, расширения эндоплазматического ретикулума и увеличения доли везикул низкой электронной плотности свидетельствуют об усилении процессов синтеза гормонов (Смиттен Н.А., 1972). Наблюдаемые признаки повреждения внутримитохондриальных структур при сохранении целостности наружной мембраны митохондрий говорят об интенсификации процессов потребления энергии в клетке. Отмечена тенденция к увеличению относительного объема митохондрий, что отражает достаточно высокий уровень регенерации структур энергообеспечения клетки в условиях ее функционального напряжения.
Характер перераспределения типов секреторных гранул в цитоплазме мозговых эндокриноцитов, вероятно, связан с преобладанием скорости выведения секрета над его синтезом в эпинефроцитах и относительной сбалансированности этих процессов в норэпинефроцитах. Наблюдаемые признаки согласуются с данными о нарушении взаимодействия между гормональным и медиаторным звеном симпато-адреналовой системы при гипокинезии сходных сроков, сопровождающемся относительным преобладанием гормональных влияний на деятельность различных систем организма (Тигранян Р.А., 1985).
Состояние капиллярного русла мозгового вещества надпочечников свидетельствует о массированном выходе содержимого цитоплазмы мозговых эндокриноцитов в кровь. В просвете синусоидных капилляров мозгового вещества определяются многочисленные секреторные гранулы. Эндотелиоциты капилляров мозгового вещества обнаруживают признаки усиленного транскапиллярного обмена в виде разрыхления базальной мембраны. Отмечена апокриновая форма секреции мозговых эндокриноцитов. Регистрация липосом корковых эндокриноцитов в просвете кровеносных капилляров мозгового вещества свидетельствует об интракапиллярном транспорте фрагментов секреторных клеток в условиях гиперфункции коры надпочечников (Савина Е.А. и соавт., 1984; Dronjak S., Nikolic J., Varagic V., 1999).
Несмотря на разрыхление волокон соединительной ткани, относительный объем, занимаемый ретикулярными волокнами, в корковом и мозговом веществе превышал значения контрольной группы. В корковом и мозговом веществе надпочечника сохранялся высокий уровень функционирования эндокриноцитов, что, вероятно, позволяло адаптироваться к непродолжительной гипокинезии и способствовало использованию «местных» источников энергообеспечения и более экономичному расходованию энергетических депо. Доказательствами такого предположения может служить высокое содержание гликогена в печени и низкий уровень молочной кислоты (70,74 % от контроля), что характеризует преобладание гликогенсинтетических процессов (Pohoska E., 1979). Подобные изменения наблюдали B.B. Nankowa et all (1998), H.S. Nagaria et P.S. Jeganathan (1999), изучавшие влияние непродолжительной гипокинезии на надпочечники. Авторы показали, что при 30-дневной иммобилизации наблюдается не только увеличение веса надпочечников, но и активизация мозгового и коркового вещества.
Элементы нервной ткани в составе мозгового вещества надпочечников обнаруживают реактивные изменения, которые затрагивают миелиновые и безмиелиновые нервные волокна и глиальные клетки. Реакция нервных элементов органа, по-видимому, носит первичный характер по отношению к изменению структуры и функции мозговых эндокриноцитов (Ажипа Я.И., 1976), являясь отражением центральных влияний на деятельность эндокринного органа.
Такие изменения коркового и мозгового вещества согласуются с данными, полученными в условиях пониженной гравитации В. Цыганковой и соавт. (2002), E.E. Howe et all (2000).
Корреляционный анализ выявил изменения взаимодействующих элементов надпочечника (рис. 6). Меньше всего связей между показателями, характеризующими энергетическую активность клеток. Выявлены корреляции элементов, обеспечивающих обменные и транспортные процессы. Достоверные связи объединяют параметры сосудистого русла (диаметр капилляров) и интенсивность реакции на щелочную фосфатазу. Появились взаимодействия между элементами надпочечника и сроками гипокинезии. Соотношение связей элементов мозгового вещества и коркового составило 0,63, данный коэффициент такой же, как и в контрольной группе (0,60).
В поле отличий морфофункциональных показателей эта группа животных по совокупности всех структурных перестроек в надпочечниках занимает пограничное положение.
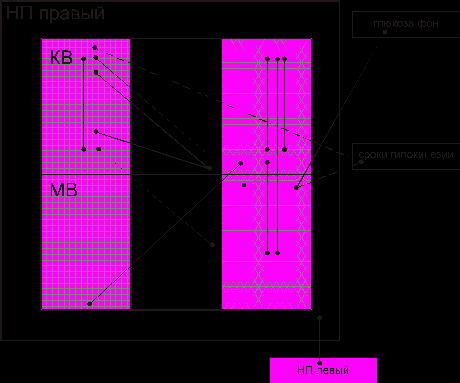
Рис. 6. Взаимодействие между элементами надпочечника при действии непродолжительной гипокинезии:
——— положительные связи,
-------- отрицательные связи,
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыИнформационный анализ показал увеличение коэффициента относительной энтропии, т.е. имело место усиление напряжений в корковом веществе надпочечника. В мозговом веществе он составил 0,79, что меньше, чем в контроле. Наибольшее напряжение было выявлено в системе элементов, обеспечивающих обменно-транспортные процессы. Диаметр капилляров, расстояние между ними в коре надпочечника характеризовались самым большим коэффициентом относительной энтропии (от 0,92 до 0,96).
Для длительного ограничения двигательной активности (свыше 100 суток) характерно ухудшение и угнетение психического состояния животных, снижение массы тела. С семнадцатой недели гипокинезии происходило урежение частоты сердечных сокращений, уменьшение выраженности электрокардиографических признаков перегрузки миокарда. Отмечался рост величины минутного объема дыхания, вентиляционного эквивалента, снижение потребление кислорода до 85% от исходного, рост вентиляционного эквивалента (Бирюкова О.В., 1998).
Надпочечники на разрезе бурого цвета. Ширина коркового и мозгового вещества по сравнению с контролем достоверно уменьшена: в клубочковой зоне – на 15,59 %, в пучковой зоне – на 30,41 %, в мозговом веществе – на 35,50 % (табл. 2). Нарушена композиция клубочковой зоны надпочечников. Форма клубочков разнообразная, больше всего клубочков округлой и неправильной формы, клубочки не всегда имеют четкие границы. Часть клубочков атрофирована. Между ними выражены тяжи соединительной ткани, которые иногда проникают в пучковую зону, их окружают клетки клубочковой зоны. Ретикулярные волокна утолщены, объем их составил 256,32 % по сравнению с контролем.
Нарушение пролиферативных процессов выражалось также в возникновении узелковой гиперплазии коры с формированием «добавочной коры». Комплекс изменений, наблюдаемых в корковом веществе надпочечников в виде активации пролиферативных процессов в подклубочковом слое, нечеткость межзональных границ, обозначается в литературе как «прогрессивная трансформация коры» (Савина Е.А. и соавт., 1980). Возможно, это связано с ускоряющейся пролиферацией и миграцией эндокриноцитов клубочковой зоны, которые являются источником обновления для клеток других зон (Wolkersdorfer G.W. et al., 1996; Okamoto M. et al., 1997; Mitani F. et al., 1999, 2003; Miyamoto H. et al., 2000; Pawlikowski M., 2005).
Увеличено количество некротически измененных клеток, особенно в клубочковой зоне. Размеры ядер эндокриноцитов пучковой зоны и мозгового вещества надпочечников не отличались от контрольных значений (табл. 3). В клубочковой зоне отмечена элонгация ядер. Длинный диаметр ядра увеличен на 18,62 %, а короткий уменьшен на 5,41 %.
При ультраструктурном исследовании корковых эндокриноцитов пучковой зоны надпочечников обнаруживаются признаки их функционального напряжения: ядра приобретают извилистые контуры с выраженным ядрышком, отмечается расширение перинуклеарного пространства, прилежание митохондрий к кариолемме, гипертрофия комплекса Гольджи и эндоплазматического ретикулума.
В то же время прогрессируют признаки ультраструктурной патологии клеток в виде появления грыжевидных выпячиваний наружной мембраны кариолеммы, очаговых просветлений матрикса и гомогенизации крист митохондрий. Сохраняется тенденция к увеличению размеров митохондрий в результате их отека. Выявлена тенденция к снижению относительного объема, занимаемого митохондриями. Изменение данного показателя происходит за счет уменьшения количества митохондрий на 52,01 % (табл. 9). Данные изменения говорят о недостаточном уровне энергетического обеспечения жизнедеятельности корковых эндокриноцитов пучковой зоны. Количество контактов митохондрий (МХ) и липидных капель (ЛК) снижено по сравнению с контрольными значениями. Липидные капли образуют плотные скопления. За счет прогрессирующего процесса слияния липидных капель сокращается их количество в цитоплазме корковых эндокриноцитов и возрастает относительный объем отдельной липидной капли.
Отмеченные изменения в целом свидетельствуют о снижении уровня синтетической активности корковых эндокриноцитов пучковой зоны на фоне сохраняющихся признаков функционального напряжения систем клетки.
Таблица 9
Показатели состояния корковых эндокриноцитов надпочечников после длительного ограничения двигательной активности,
-
Показатели
Контроль
Опыт
Относительный объем МХ, %
23,05±2,54
17,24±1,64
Относительный объем ЛК, %
41,53±3,90
48,33±2,96
Количество МХ, шт. на 100 мкм2
25,90±3,12
12,43±1,24*
Количество ЛК, шт. на 100 мкм2
7,40±0,71
4,20±0,40*
Количество контактов МХ и ЛК,
шт. на 100 мкм2
15,35±2,40
9,38±1,24*
* – р<0,05 (достоверные отличия от контроля)
Капиллярное русло во всех зонах полнокровно. Выявлены нарушения функционального состояния капилляров в виде диапедезных кровоизлияний в клубочковой и пучково-сетчатой зонах надпочечников.
В мозговом веществе надпочечников отмечены дистрофические изменения эндокриноцитов. Увеличено, по сравнению с предыдущей серией, число некротически измененных клеток. Значительное количество мозговых эндокриноцитов обнаруживает признаки внутриклеточного отека.
В то же время, для многих мозговых эндокриноцитов характерны признаки оводнения секреторных гранул и цитоплазмы клеток, что говорит о поддержании уровня секреторных процессов в мозговом веществе надпочечников. Отмечены прогрессирующее снижение относительного объема митохондрий за счет уменьшения их количества (на 37,93%) в цитоплазме клеток, явления набухания и тотальной деструкции митохондрий. Данные изменения отражают недостаточность энергетического обеспечения мозговых эндокриноцитов. Относительный объем секреторных гранул не обнаруживает достоверных отличий от уровня контроля. Выявлено уменьшение количества секреторных гранул на 27,05 % за счет субпопуляции полупустых гранул, в то время как количество гранул остальных типов остается на уровне контроля.
Диаметр капилляров (табл. 4) в мозговом веществе увеличен. В кровеносном русле определяются фрагменты цитоплазмы и ядер мозговых эндокриноцитов, что свидетельствует о нарастании альтерации в паренхиме железы.
Наблюдается выраженная реакция стромальных элементов мозгового вещества в виде расширения прослоек соединительной ткани между группами мозговых эндокриноцитов и вокруг капилляров.
Относительный объем, занимаемый ретикулярными волокнами, увеличен в 2-3 раза как в корковом, так и в мозговом веществе надпочечника.
Взаимодействие между элементами надпочечника при действии длительной гипокинезии отличалось от всех исследуемых групп. По сравнению с контролем, количество корреляционных связей сократилось на 72,73 %. Отсутствуют связи между структурами, обеспечивающими энергетические, синтетические, обменные процессы в эндокриноцитах (рис. 7). Имеют место связи на организменном уровне (с глюкозой крови и частотой сердечных сокращений в конце эксперимента).
Анализ морфофункционального единства по С.Б. Стефанову выявил, что по совокупности изменений в корковом и мозговом веществе надпочечника группа животных после длительной гипокинезии занимает зону отрицательных показателей (KQ ).
Информационный анализ показал увеличение коэффициента относительной энтропии во всех зонах надпочечника, т.е. имело место усиление напряжений в коре и мозговом веществе (h>0,90). Наибольшие напряжения были выявлены в системе элементов, определяющих обменные и транспортные процессы. По расстоянию между капиллярами в корковой зоне, пучковой зоне и мозговом веществе относительная энтропия составила более 0,93, по площади, занимаемой ретикулярными волокнами – h=0,94-0,97.
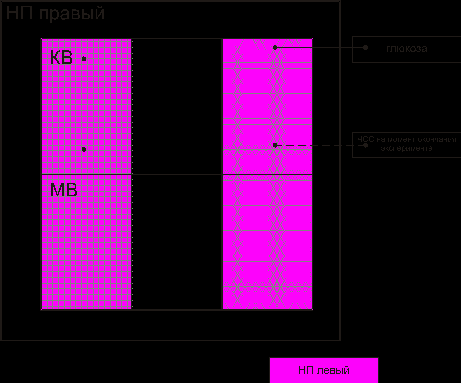
Рис. 7. Взаимодействие между элементами надпочечника при действии длительной гипокинезии:
——— положительные связи,
-------- отрицательные связи,
показатели, характеризующие обменные процессы, показатели, характеризующие энергетические процессы, показатели, характеризующие синтетические процессыВЫВОДЫ
1. Двигательная нагрузка или ее ограничение (гипокинезия) при однократном или длительном применении вызывали качественные и количественные преобразования в разных зонах надпочечника – преимущественно в коре или одновременно и в корковом, и в мозговом веществе – на всех иерархических уровнях (от субклеточного до органного), которые определялись величиной и продолжительностью воздействия.
2. При однократной двигательной нагрузке у собак (бег со скоростью 15 км/ч в течение 27 мин) в надпочечниках достоверно увеличивались: ширина коркового вещества – на 18,99 %, мозгового вещества – на 22,04 %, диаметр ядер эндокриноцитов пучковой зоны (12,76 %) и мозгового вещества (7,81 %). Интенсивность реакции на НАД*Н2–дегидрогеназу увеличилась в клубочковой, пучковой зонах и мозговом веществе (на 47,69 %, 56,05 %, 41,05 %, соответственно). Ферментативная активность реакций на щелочную фосфатазу уменьшилась во всех зонах (p<0,01).
3. При однократной предельной двигательной нагрузке (бег со скоростью 15 км/ч, в среднем, в течение 193 мин) выявлены изменения коркового и мозгового вещества надпочечников. Увеличивалась ширина клубочковой и пучковой зон (134,25 % и 114,95 %), размеры ядер в клетках клубочковой, пучковой зон и в мозговом веществе (на 17,31 %, 4,76 %, 9,62 %). Интенсивность реакции на НАД*Н2 –дегидрогеназу выросла в клубочковой, пучковой зонах и в мозговом веществе (на 28,30 %, 26,47 %, 16,82 %). Диаметр капилляров пучковой зоны составил 114,68 %. Уменьшилась оптическая плотность продукта реакции щелочной фосфатазы в клубочковой, пучковой зонах и в мозговом веществе (на 20,22 %, 17,91 %, 14,46 %). В корковых эндокриноцитах достоверно снижен относительный объем, занимаемый липидными каплями, на 33,66 %. В мозговых эндокрицитах количество митохондрий достоверно увеличено.
4. После многократных двигательных нагрузок у собак (бег со скоростью 15 км/ч от 11 мин в первые дни тренировок до одного часа и более в сутки в конце опыта) морфологические изменения выявлены в корковом веществе надпочечников. Увеличивалась ширина клубочковой и пучковой зон на 7,69 %, размеры ядер в пучковой зоне – на 6,15 %, диаметр капилляров в пучковой зоне – на 125,26 %, расстояние между капиллярами в клубочковой и пучковой зонах – на 23,32 % и на 32,71 %. Оптическая плотность продуктов реакции сукцинатдегидрогеназы и 3β–ОН–стероиддегидрогеназы в клубочковой зоне и пучковой зоне выше, чем в контроле. Выявлено снижение активности реакции на щелочную фосфатазу в корковом веществе надпочечника на 23,41 %.
5. После многократного чередования тренировок с разной продолжительностью (бег со скоростью 15 км/ч до двух часов в сутки в течение двух недель и бег в последующую неделю – по 10-15 мин) морфологические изменения происходят, в основном, в корковом веществе надпочечников. Увеличились: ширина коркового вещества надпочечников - на 16,27 %, диаметр ядер клеток клубочковой зоны – на 16,77 %. Значительно расширен диаметр кровеносных капилляров в коре надпочечников. В корковых эндокриноцитах уменьшилось количество митохондрий и липидных капель – на 42,47 % и на 40,41 % соответственно. В мозговом веществе достоверно увеличен только диаметр капилляров.
6. После непродолжительного ограничения подвижности (около 30-и суток) выявлены структурные изменения в корковом и мозговом веществе надпочечников. Увеличились ширина коркового и мозгового вещества (на 24,81 % и 16,39 % соответственно), размеры ядер в клетках всех зон, диаметр кровеносных капилляров в пучковой зоне на 26,11 %, в мозговом веществе – на 33,38 %. Интенсивность реакций на НАД*Н2 –дегидрогеназу в корковом веществе, на 3β–ОН–стероиддегидрогеназу в клубочковой зоне достоверно больше, чем в контрольной группе. В корковых эндокриноцитах уменьшено количество липидных капель, число контактов митохондрий и липидных капель.
7. Длительное ограничение двигательной активности (гипокинезия более 100 суток) привело к развитию дистрофических изменений как в корковом, так и в мозговом веществе надпочечников. Увеличено количество некротически измененных клеток, наблюдается атрофия части клубочков. Ширина клубочковой зоны, пучковой зоны и мозгового вещества составила 84,41 %, 69,59 % и 64,50 % относительно контрольной группы. Диаметр капилляров достоверно увеличен во всех зонах. Ретикулярные волокна утолщены, относительная площадь, занимаемая ими, увеличена в 2 и более раза. По сравнению с контролем на 52,01 % уменьшено количество митохондрий в корковых и на 37,93 % – в мозговых эндокриноцитах. Достоверно снижено количество секреторных гранул в клетках мозгового вещества (на 27,05 %).
8. В надпочечниках животных, показавших максимальное время бега при предельных нагрузках (255,62±27,24 мин) выявлены признаки повышения функциональной активности в корковом веществе надпочечников: ширина клубочковой и пучковой зон на 13,85 % и на 22,01 % (p<0,01) превышает соответствующие показатели контрольной группы. Диаметр кровеносных капилляров клубочковой и пучковой зон увеличился. Интенсивность реакции на сукцинатдегидрогеназу увеличилась в клубочковой зоне – на 16,27 %, в пучковой зоне – на 14,32 %. Достоверно возросла оптическая плотность продукта реакции на НАД*Н2 –дегидрогеназу в корковом веществе надпочечников.
В надпочечниках животных с непродолжительным временем бега (66,67±14,15 мин) выявлены изменения преимущественно в мозговом веществе: ширина мозгового вещества достоверно увеличена на 12,00 %. Размеры ядер мозговых эндокриноцитов составляют 116,87 %. Интенсивность реакции на НАДН2 –дегидрогеназу возросла на 26,78 %. В корковом веществе отмечено достоверное увеличение относительного объема, занимаемого ретикулярными волокнами.