
Мутации и мутационный процесс в природных популяциях
| Вид материала | Диссертация |
| Drosophila melanogaster D. melanogaster 1.3. Повышенная частота мутаций гена Drosophila melanogaster Drosophila melanogaster |
- «Элементарные эволюционные факторы», 91.21kb.
- Общие свойства мутаций, 125.22kb.
- Проблема мутации Введение, 225.06kb.
- Мутационная изменчивость, 47.38kb.
- Аннотация рабочей программы дисциплины охрана окружающей среды и рациональное использование, 40.26kb.
- За курс 9 класса билет, 42.57kb.
- В различных клеточных популяциях, 571.27kb.
- Лекция 3 Переработка природных углеводородных газов, 50.32kb.
- Содержание Введение, 120.11kb.
- Молекулярно-генетические и клинико-генотипические особенности муковисцидоза в российских, 924.89kb.
* Ежегодно исследовалось несколько популяций из различных регионов. За период 1981-1993 гг. исследовалось 36 популяций. Например, кроме популяции Умань, в 1986 г. были изучены популяции: Чернобыль, Ужгород, Брест, Одесса, Запорожье и Самарканд,
в 1990 г. - Запорожье, Магарач, Минеральные Воды, Хорог, Душанбе.
** N - общее количество изученных Х-хромосом.
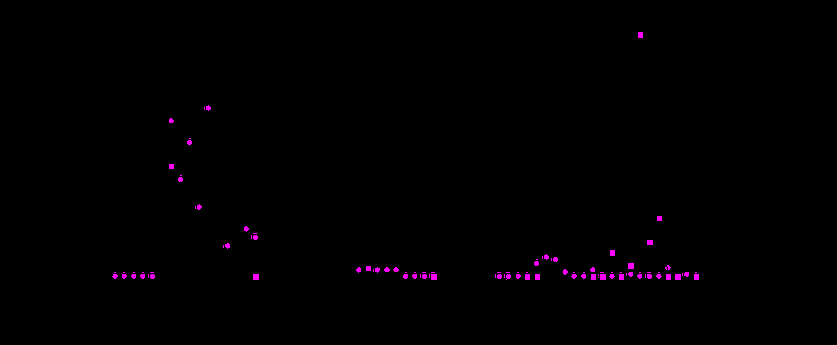
Риc. 1. Концентрация и частота возникновения yellow-мутаций в природных популяциях Drosophila melanogaster на территории СССР в период 1931-1993 гг. Данные по популяции Умань приведены отдельно.
Обозначения: ¡ - частота возникновения yellow-мутаций в популяциях;
- концентрация yellow-мутаций в популяциях;
¨ частота возникновения yellow-мутаций в популяции Умань;
n - концентрация yellow-мутаций в популяции Умань.
Данные по годам были получены: 1931-1935 - [Дубинин и др.,1934 а, б; Дубинин,1937]; 1937 - [Оленов, Хармац,1938; Гершензон,1941; Берг,1948]; 1938 - [Берг,1948; Дусеева,1948]; 1939 - [Гершензон,1941; Берг,1948; Дусеева,1948]; 1940 - 1943 - [Дусеева, 1948]; 1941 - [Берг,1948]; 1945 - [Берг,1948; Дусеева, 1948]; 1946-1958, 1957-1964 - [Берг,1948, 1961; Berg, 1965]; 1963-1984 - [Захаров, Голубовский,1985; Голубовский и др.,1987]; 1972-1993 - [Захаров и др.,1995]; 1985-1993 - [Захаров, Скибицкий, 1995; Захаров и др., 1995].
В период с 1947 по 1956 годы популяции не исследовались. С 1966 по 1971 годы популяции исследовались Р.Л. Берг и качественно известно, что содержание yellow-мутаций в популяциях не превышало фонового уровня.
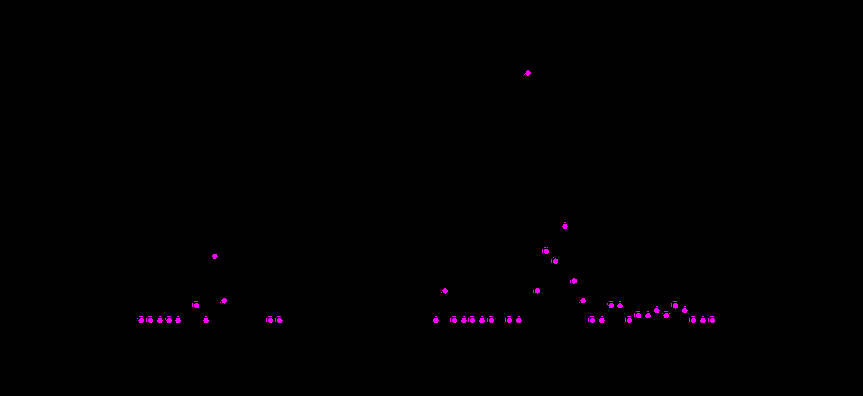
Рис. 2. Концентрация и частота возникновения сцепленной с полом мутации singed bristles в природных популяциях Drosophila melanogaster на территории СССР в период 1931-1993 года.
Обозначения: ¡ - частота возникновения singed;
- концентрация singed.
Данные по годам были получены следующими авторами: 1931-1935 - [Дубинин и др., 1934 а, б; 1937]; 1937 - [Оленов, Хармац, 1938; Гершензон, 1941; Berg, 1974]; 1938 - [Berg, 1974]; 1939 - [Гершензон, 1941; Berg, 1974]; 1940, 1945, 1946, 1957-1964 - [Berg, 1965]; 1960 - 1973 - [Berg, 1974 a, b]; 1974 - 1983 - [Захаров, 1984; Захаров, Юрченко, 1985]; 1984 - 1993 - [Захаров и др., 1995; Захаров и др., неопубликованные данные].
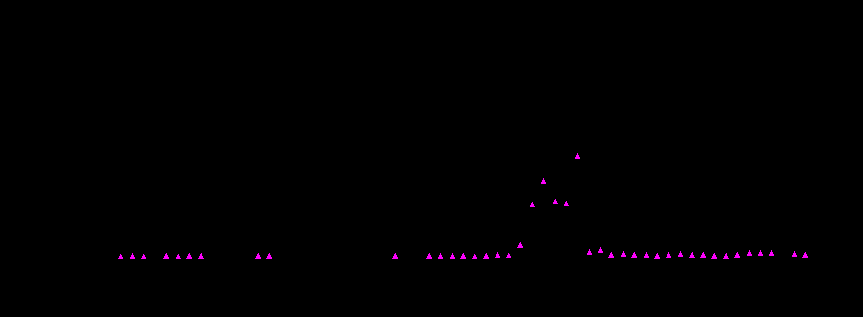
Рис. 3. Концентрация аномалии abnormal abdomen в природных популяциях Drosophila melanogaster на территории СССР в 1931-1993 гг.
Обозначения: t- концентрация abnormal abdomen среди самок;
D - концентрация abnormal abdomen среди самцов.
Данные по годам были получены следующими исследователями: 1933-1935 - [Дубинин и др., 1934 а, б; 1937]; 1937-1940, 1945, 1946, 1957, 1960-1971 - [Berg, 1972]; 1972, 1973 - [Berg, 1973, 1974]; 1973-1993 - [Голубовский и др., 1974; Захаров и др., неопубликованные данные].
Примечание. В отличие от объективно регистрируемых мутаций yellow и singed при сравнении результатов, полученных разными авторами, при количественной оценке признака abnormal abdomen в популяциях, в силу вариабельности его фенотипической экспрессии, следует учитывать субъективность оценок исследователя при проведении границы между нормой и аномальным признаком.
Таблица 2. Сравнительная характеристика мутационных периодов в природных популяциях Drosophila melanogaster Евразии в 1931-1994 гг.
| Годы | 1937-1946 | 1982-1991 |
| Длительность | Не менее 10 лет | 10 лет |
| Мутация, тип аллеля | Аллели как yellow-2-, так и yellow-1-типа. Другие гены: singed, forked, white. | Только аллели типа yellow-2 |
| Характер наследования | Сцепленная с полом рецессивная мутация | Сцепленная с полом рецессивная мутация |
| Характер распространения | Повсеместное распространение yellow-мутаций в популяциях Евразии. Мутация white была обнаружена только в 2 из 35 популяций. | Локальная вспышка, проявилась только в популяции Умань (Украина) |
| Концентрация | 0.05-0.25% | В отдельные годы - до 3.2% |
| Частота возникновения | 0.04-0.15% | В среднем 0.03% |
| Отношение концентрации и частоты возникновения | Концентрация в популяции и частота возникновения мутации de novo примерно одного уровня. | Концентрация в популяции превышает мутабильность и концентрация на порядок величин выше, чем в 30-40-х годах. |
| Генетическая нестабильность | Генетическая нестабильность аллелей не изучалась. Проверялось ревертирование только некоторых аллелей [Гершензон, 1941]. | Серии нестабильных аллелей yellow-2. Зарегистрированы супернестабильные аллели, мутирующие с частотой до 11% |
[Использованы данные авторов, перечисленных в подписях под рис. 1-3].
| 1968-1975 | 1973-1979 |
| 8 лет | 7 лет |
| Аномалии, сходные с мутациями типа abnormal abdomen | singed bristles, фенотипически и мутационно различающиеся аллели |
| Рецессивные и полудоминантные мутации, мутации с неполной пенетрантностью и изменчивой экспрессивностью, обусловленные разными генами, локализованными в различных хромосомах. | Сцепленная с полом рецессивная мутация |
| Повсеместное распространение в популяциях D. melanogaster Евразии | Обнаружены в различных популяциях Евразии. Скорее всего, следует отнести к мозаичному характеру распространения по ареалу вида. |
| Самки - от 10 до 60%. Самцы - от 10 до 30%. | 0.03-0.05% |
| Не оценивалась. | 0.01% |
| Чрезвычайно высокая концентрация, возможно, в результате повышения мутабильности фенотипически сходных мутаций разных локусов. Похоже на явление "гетерогения сходных фенов". Не исключено сильное влияние селективных факторов. | Концентрация и частоты возникновения близки. |
| Не исследована. | Множественные серии нестабильных аллелей singed. Супермутабильные аллели. Плеяды нестабильных аллелей. |
Таблица 3. Спектр и разнообразие аллелей гена singed bristles в зависимости от направлений и частот мутирования, выделенных из природных популяций Drosophila melanogaster в 1973-1990 гг. [Golubovsky et al., 1977; Захаров, 1984б; Захаров, Голубовский, 1984а; Захаров, Юрченко, 1985].
| Класс | Аллель* | Происхождение, год | Направление мутирования | Частота мутирования |
| A1 | sns-U36 | Умань, 1979 | snf | 3x10-3 |
| | | | sn+ | 1x10-3 |
| | | | snm | 7x10-4 |
| | sns-G78 | Геленджик, 1978 | snm | 1x10-3 |
| | | | snf | 1x10-4 |
| | | | sn+ | 7x10-5 |
| A2 | sns-K3-2 | Краснодар, 1978 | sn+ | 8x10-4 |
| | | | snf | 7x10-5 |
| A3 | sns-V49 | Владивосток, 1975 | sn+ | 3x10-3 |
| | | | snm | 3x10-4 |
| | sns-N88 | Нальчик, 1988 | snm | 3x10-4 |
| | | | sn+ | 6x10-4 |
| A4 | sns-T77-27 | Ташкент, 1973 | sn+ | 1x10-2 ** |
| | sns-11 | sns-T77-27 | sn+ | 2x10-1 ** |
| A5 | sns-90719 | Магарач, 1990 | стабильный*** | |
| B1 | snm-T63-15 | Ташкент, 1973 | snf | 1x10-2 |
| | | | sn+ | 3x10-3 |
| | | | sns | 2x10-3 |
| | | | Таблица 3, | продолжение |
| Класс | Аллель* | Происхождение, год | Направление мутирования | Частота мутирования |
| B2 | snm-K33-13 | Краснодар, 1974 | sn+ | 1x10-3.** |
| | | | snf | 1x10-3 |
| | snm-90667 | Магарач, 1990 | sn+ | 3x10-4 |
| | | | snf | 1x10-4 |
| B3 | snm-TB76 | Тбилиси, 1976 | sn+ | 2x10-3 |
| | | | sns | 9x10-4 |
| | snm-Z3195 | Запорожье, 1986 | sn+ | 5x10-4 |
| | | | sns | 1x10-4 |
| B4 | snm-K88-9 | Краснодар, 1974 | sns | 1x10-3 ** |
| B5 | snm-Z514 | Запорожье, 1987 | стабильный | |
| C1 | snf-D163 | Душанбе, 1981 | sn+ | 1x10-1 |
* Обозначение фенотипического проявления аллелей singed: sns - strong, snm - moderate, snf - faint, sn+ - дикий тип, щетинки и волоски практически неотличимы от нормального фенотипа.
** Данные по частоте мутирования этих аллелей взяты из работы [Golubovsky et al., 1977].
*** Здесь и в табл. 4 аллель обозначен как "стабильный" только в том случае, если его стабильность установлена (оценена его частота мутирования на множестве не менее 10 тыс.).
1.3. Повышенная частота мутаций гена singed в природных популяциях Drosophila melanogaster в 1973-1979 гг.
В 1973-1979 годы в природных популяциях Drosophila melanogaster в повышенной концентрации встречалась другая сцепленная с полом мутация - singed bristles [Berg, 1974; Ivanov, 1974; Иванов, Голубовский, 1977; Захаров, Голубовский, 1984; Захаров, 1984] (рис. 2). Повышенная концентрация мутаций гена singed наблюдалась на фоне повышенной мутабильности нормальных аллелей и вспышка продолжалась до 1979 года. В этот период были выделены нестабильные аллели гена singed, которые при изучении их мутационных свойств дали начало сериям фенотипически различающихся аллелей данного локуса (Табл. 3). В большинстве своем аллели singed из природы оказались генетически нестабильными [Ivanov, 1974; Иванов, Голубовский, 1977; Golubovsky et al., 1977; Голубовский, 1977; Захаров, 1984; Захаров, Голубовский, 1984а; Захаров, Юрченко, 1985]. В работе М.Д. Голубовского и Е.С. Беляевой (1985), методом гибридизации in situ на препаратах хромосом слюнных желез было показано, что в цитологическом районе локализации singed в sn-Х-хромосомах из природы выявляются различные типы мобильных элементов - часть аллелей связана с внедрением P-элементов, часть с внедрением мдг3.
Повышенная концентрация мутаций singed в природных популяциях, скорее всего, явилась следствием повышенного мутирования в локусе singed, а не результатом действия отбора. Об этом свидетельствует, во-первых, тот факт, что из 19 выделенных из природы аллелей 12 были зарегистрированы в момент их возникновения, во-вторых, аллели singed характеризовались высокой мутабильностью и обладали свойством многократного мутирования, сохраняющегося на протяжении десятилетий содержания линий в лаборатории. Возникающие при этом фенотипически нормальные аллели (неотличимые от дикого типа) оказывались в такой же степени нестабильными, как и аллели с мутантным выражением признака. В-третьих, аллели singed с фенотипически сходным проявлением могли существенно различаться по спектру и частотам мутирования как в половых, так и в соматических клетках, что говорит в пользу их независимого происхождения.
Обычно размножение мутантных Х-хромосом и генетический анализ сцепленных с полом мутаций удобно проводить в скрещиваниях анализируемых самцов с самками лабораторных тестерных линий (со сцепленными Х хромосомами, Basc и др.). Для некоторых аллелей гена singed это еще и вынужденная мера, так как гомозиготные самки оказываются стерильными. Это ограничение снимается для yellow-X-хромосом, генетический анализ мутирования которых можно проводить на основе анализа по потомству yellow-самок, не прибегая к скрещиваниям с лабораторными тестерными линиями. Отметим, что именно в наших работах было показано, что повышенная мутабильность в локусе singed не связана с межлинейными скрещиваниями мух из природы с лабораторными линиями, а наблюдается на природном генетическом фоне [Захаров, 1987]. Это было подтверждено и для случая повышения концентрации мутаций по гену yellow в популяции Умань - нестабильное состояние yellow2-аллелей наблюдается и в генотипе самок на природном цитотипе и генотипе [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров, Скибицкий, 1995].
Глава 2. ГЕНЕТИЧЕСКИЕ СВОЙСТВА АЛЛЕЛЕЙ ГЕНОВ Х ХРОМОСОМЫ, ВЫДЕЛЕННЫХ ИЗ ПРИРОДНЫХ ПОПУЛЯЦИЙ DROSOPHILA MELANOGASTER В ПЕРИОД ВСПЫШКИ МУТАЦИЙ
После вспышки мутаций в 30-40-х годах, концентрация мутаций yellow в природных популяциях на протяжении почти 4 последующих десятилетий не превышала фоновых и находилась на уровне порядка 10-5. В 1982 году в природной популяции Drosophila melanogaster Умани (Украина) нами было отмечено повышение концентрации мутации yellow [Захаров, Голубовский, 1985, 1989; Голубовский и др. 1987]. Распространение получили только аллели типа yellow-2. Концентрация yellow-2-мутаций с 1982 года по 1991 год в популяции Умань поддерживалась на уровне порядка 10-3-10-2 (Табл. 1). В отдельные годы концентрация yellow-2-Х-хромосом достигала значений 3%. Мутантные самцы и гетерозиготные самки по yellow-2-хромосоме в отдельные годы составляли 4%.
Основным свойством части выделенных yellow-2-аллелей является их генетическая нестабильность (Табл. 4). Частота мутирования yellow-2 в таких линиях могла колебаться от 10-4 до 10-2. Более того, получаемые в результате мутирования нормальные производные - у+, также могли оказаться нестабильными. Это свойство наблюдалось для аллелей, выделенных на протяжении всего десятилетнего периода вспышки мутаций, а с учетом времени развития Drosophila - на протяжении смены сотен поколений, и нестабильное состояние сохраняется столь продолжительное время как для выделяемых из природы аллелей, так и для содержащихся в лаборатории и вновь возникших в лабораторных условиях мутантных и ревертантных аллелей природных Х хромосом. Подчеркнем, что мутационная активность ограничилась только одной популяцией - Уманью, и одним локусом - yellow.
Выделенные из исследованных в этот период популяций Drosophila melanogaster мутантные аллели других генов могли быть как стабильными, например, некоторые аллели dusky, lozenge, rudimentary singed и vermilion, так и нестабильными - dusky, miniatute и singed (Табл. 5) [Захаров и др., 1995; Захаров, Скибицкий, 1994, 1995]. Само по себе присутствие в популяции нестабильных аллелей оказывается не достаточным для поддержания мутации в повышенной концентрации. Об этом свидетельствуют и данные сравнительного анализа соотношения между мутабильностью и концентрацией мутаций двух периодов мутабильности. Если в 30-40-х годах концентрация yellow-мутаций составляла 0,05-0,25% и частота возникновения de novo равнялась 0,04-0,15%, то вспышка 80-х годов имела совершенно другую количественную характеристику. Концентрация yellow-мутаций в популяции Умань в среднем составила 0,9% и в отдельные годы достигала значений 3,2%, что значительно превышало средний уровень частоты возникновения yellow в этот период в популяции - 0,03%. Следовательно, в популяции Drosophila melanogaster Умани должны были сложиться специфические условия, во-первых, для активации мутабильности в локусе yellow и, во-вторых, для сохранения мутантных аллелей и поддержания мутации в повышенной концентрации на протяжении целого десятилетия. Известно, что нестабильное состояние генов, в том числе и yellow, обусловлено процессами внедрения-вырезания мобильного элемента (а в некоторых случаях в гене могут быть локусы-мишени для различных типов мобильных элементов), поэтому полученные нами данные говорят о том, что в популяции должны были возникнуть и существовать длительное время генетические и (или) эпигенетические факторы, инициирующие подвижность мобильных элементов, которые и позволяют поддерживать в популяции инсерционные мутации в высокой концентрации.
Таким образом, выявлена и прослежена динамика локального повышения локус-специфической мутабильности и концентрации yellow-мутаций в природной популяции Drosophila melanogaster. Длительность и систематичность генетико-популяционных исследований популяции Drosophila melanogaster Умани позволяют использовать ее в качестве контрольной. Полученные результаты важны для понимания закономерностей спонтанного мутационного процесса и роли в нем инсерционных мутаций, которые являются значительным компонентом генома.
Таблица 4. Группы множественных yellow-2-аллелей, выделенных из природной популяции Drosophila melanogaster Умани в 1983 и в 1990 годах, различающихся по характеру нестабильности исходных аллелей и их мутантных производных [Захаров, Голубовский, 1985; Голубовский и др., 1987; Захаров, Скибицкий, 1995].
| Класс | Аллели yellow-2- | Мутирование от y2 к y+ | Частота мутирования | |
| | | частота | аллели | от y+ к y2 |
| I | y2-стабильные | 0 | | |
| II | y2U1-5 | 2x10-3 | y+(1-5) | стабильный |
| | y2U23-10 | 7x10-3 | y+(23-10) | 1x10-4 |
| | y2U1-4 | 1x10-3 | y+(1-4) | 2x10-3 |
| III | y2U1-6 | 6x10-4 | y+691 | стабильный |
| | y2U23-11 | 8x10-4 | y+(23-11) | 4x10-4 |
| | y2U23-4 | 4x10-4 | y+(23-4) | 3x10-3 |
| IV | y2U23-3 | 3x10-4 | y+(23-3)* | 3x10-2 |
| | y2U23-8 | 1x10-4 | y+(23-8)* | 1.1x10-1 |
* Возникшие ревертанты у+-аллели в группе IV, оказываются
более нестабильными, чем их исходные у2-аллели.