
Влияние экологических факторов на содержание тяжелых металлов и cs-137 в микобиоте лесных экосистем 03. 00. 16 Экология
| Вид материала | Автореферат |
| 4.3. Содержание ТМ и Cs-137 в плодовых телах макромицетов, произрастающих в различных лесорастительных условиях Russula spp. |
- Влияние Экологических факторов на аллергическую заболеваемость детского населения российской, 441.49kb.
- План Введение 1 Действие тяжелых металлов на растительные организмы 3 Химическая природа, 600.02kb.
- Использование природного капитала лесных экосистем с учетом трансграничных факторов, 613.87kb.
- Удк количественное определение содержания тяжелых металлов в пробах почвы атомно-абсорбционным, 161.57kb.
- Рабочая программа дисциплины «Экология организмов (экология растений)» Код дисциплины, 139.35kb.
- Ержание подвижных форм тяжелых металлов Вцелом уровень загрязнения поверхностного почвенного, 31.17kb.
- Влияние факторов внешней среды на деятельность организации (на примере ОАО «Кировский, 44.93kb.
- Московский Государственный Институт Стали и Сплавов (Технологический Университет) Кафедра, 461.38kb.
- Мадиева Айгуль Шамильевна учитель биологии и химии высшей категории моу «Варваринская, 204.03kb.
- Лекция Влияние различных факторов на пластичность металла. Влияние омд на структуру, 126.99kb.
4.3. Содержание ТМ и Cs-137 в плодовых телах макромицетов, произрастающих в различных лесорастительных условиях
Результаты исследования содержания ТМ и Cs-137 в представителях дереворазрушающих и почвенных макромицетов, произрастающих в различных типах леса в Кузоватовском лесхозе, представлены в табл. 4.
Таблица 4
Содержание ТМ и Cs-137 в дереворазрушающих и почвенных макромицетах в различных типах леса Кузоватовского лесхоза
| Тип леса | Содержание ТМ ( 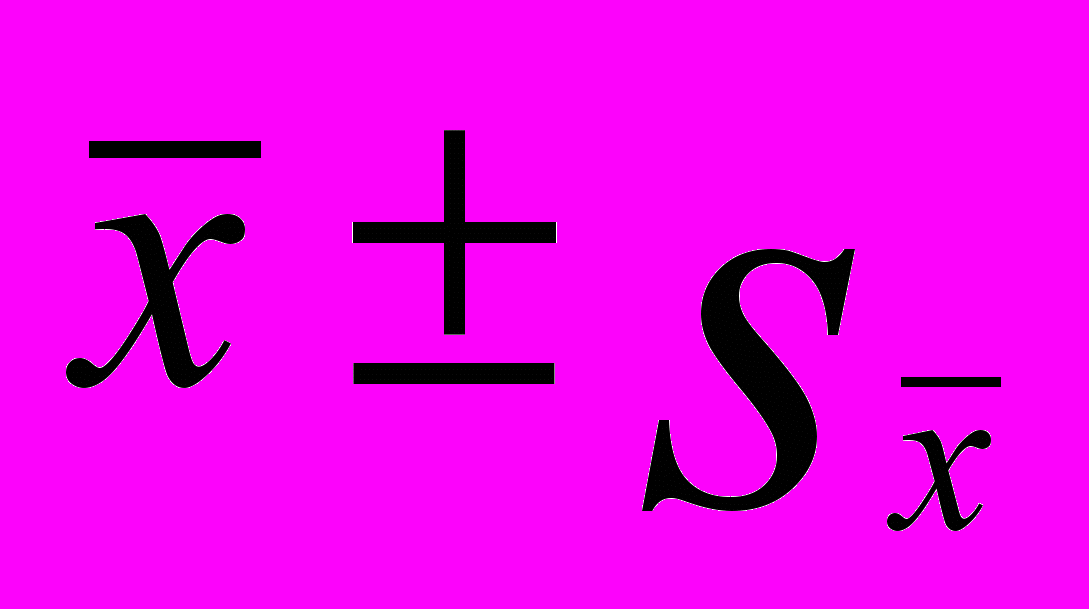 ), мг/кг ), мг/кг | Cs-137, Бк/кг | |||||||
| Cu | Zn | Ni | Cd | Pb | Co | Fe | Σ ТМ | ||
| Phellinus pini | |||||||||
| С лш | 1,54± 0,14 | 19,87+ 0,31 | 4,34+ 0,25 | 0,40+ 0,08 | 3,69+ 0,22 | 2,66+ 0,18 | 22,68+ 0,62 | 55,18 | 38,95± 1,83 |
| С брзм | 2,09+ 0,10 | 25,34+ 0,37 | 4,98+ 0,22 | 2,32+ 0,21 | 5,04+ 0,31 | 2,94+ 0,20 | 27,01+ 0,33 | 69,72 | 43,84± 2,67 |
| С ч | 4,54+ 0,26 | 26,74± 0,94 | 10,56+ 0,18 | 2,98+ 0,11 | 7,86+ 0,36 | 3,03+ 0,23 | 29,95+ 0,75 | 85,66 | 59,39± 2,15 |
| Phellinus robustus | |||||||||
| Д оссн | 2,54+ 0,11 | 8,40+ 0,68 | 3,06+ 0,26 | 1,04+ 0,11 | 2,06+ 0,16 | 1,95+ 0,25 | 115,32+ 11,31 | 134,37 | 1,76± 0,32 |
| Д сняс | 3,05+ 0,23 | 12,34+0,21 | 2,99+ 0,23 | 0,87+ 0,08 | 3,98+ 0,12 | 2,54+ 0,32 | 115,61+ 3,61 | 141,38 | 5,44± 1,28 |
| Д рзтр | 3,04+ 0,26 | 10,87+0,32 | 4,34+ 0,14 | 1,05+ 0,09 | 5,36+ 0,11 | 3,00+ 0,18 | 116,77+ 10,27 | 144,43 | 9,60± 1,09 |
| Russula spp. | |||||||||
| С лш | 2,85± 0,36 | 9,11± 0,23 | 12,93±0,78 | 1,02± 0,08 | 8,84± 0,65 | 2,21± 0,16 | 221,56 ± 14,53 | 258,52 | 13,55± 0,58 |
| С брзм | 5,98± 0,28 | 10,11± 0,32 | 12,74± 0,64 | 1,98± 0,11 | 8,76± 0,52 | 5,04± 0,47 | 220,65 ± 13,73 | 265,26 | 97,81±3,47 |
| С ч | 6,28± 0,11 | 13,95±0,14 | 13,61±0,40 | 2,08± 0,09 | 9,63± 0,13 | 5,07± 0,41 | 232,58±18,51 | 283,20 | 135,8± 7,14 |
| Д оссн | 20,19± 0,24 | 17,34± 0,23 | 11,15± 0,14 | 1,10 ± 0,08 | 9,12± 0,31 | 2,45± 0,12 | 304,24± 20,28 | 365,59 | 266,8± 4,55 |
| Д сняс | 19,04±0,13 | 22,07±0,24 | 15,04± 0,12 | 1,88± 0,21 | 9,02± 0,12 | 3,26± 0,11 | 301,20±18,32 | 371,51 | 963,66±6,71 |
| Д рзтр | 21,86±0,15 | 28,32±0,31 | 21,12±0,33 | 2,39± 0,16 | 9,89± 0,41 | 3,73± 0,32 | 302,06±15,32 | 389,37 | 1347± 24,11 |
Анализ данных табл. 4 показывает, что суммарное содержание ТМ по мере снижения оптимальности условий произрастания: от сосняка черничного к сосняку лишайниковому, от дубняка разнотравного к осоково-снытьевому, и удельная активность Cs-137 в плодовых телах дереворазрушающих и почвенных грибов, произрастающих в Кузоватовском лесхозе, достоверно уменьшается. Если рассматриваемые типы леса расположить в эдафическом ряду по степени убывания увлажнения почво–грунтов, то получим следующие ряды:
- дубняк разнотравный (Дрзтр)> дубняк снытьево-ясменниковый (Д сняс)> дубняк осоково-снытьевый (Д оссн);
- сосняк черничный (Сч)> сосняк бруснично-зеленомошный (С брзм)> сосняк лишайниковый (Слш).
Дело в том, что в более богатых условиях произрастания (Сч, Дрзтр) почвы отличаются наибольшим плодородием (Розов, 1939; Морозов, 1994), т.е. высоким содержанием гумуса (от 4,5 до 7-9%) и высоким уровнем рН (от 5,1 до 5,8), следовательно, в них аккумулируется больше микроэлементов, но в связанной труднодоступной форме для грибов и растений. Однако даже при отсутствии техногенного фактора грибы, как показывают результаты наших исследований, способны извлекать из субстрата значительное количество ТМ и Cs-137.
В естественных условиях почвы Ульяновской области достаточно сильно обогащены Fe, этим обуславливается наибольшее содержание этим элементом в плодовых телах исследуемых макромицетов. Pb содержится в грибах в количествах, во много раз превышающих ПДК (0,5 мг/кг) – от 4,54 мг/кг (Ph. pini) до 13,06 мг/кг (C. versicolor), установленных для грибов.
Наибольшие концентрации Co отмечены в представителях рода Russula spp., произрастающих в Сч (5,07±0,41 мг/кг), наименьшее – в плодовых телах Ph. pini, произрастающих в Слш. Данный элемент в значительных количествах входит в систему метаболизма грибов, активируя белковый обмен. Максимальная удельная активность Cs-137 обнаружена в Russula spp. (1347± 24,11 Бк/кг), собранных в Дрзтр, с наиболее благоприятными условиями для развития живых организмов, наименьшая – в Ph. pini (1,76± 0,32 Бк/кг). Повышенное содержание металлов и Cs-137 может быть обусловлено биогенным характером элементов и особенностями ферментативных процессов, присущих представителям грибного «царства» (Кирилюк, 2006). Исследования, проведенные в Инзенском лесхозе, показали аналогичные результаты.
Результаты наших исследований подтверждают данные, полученные ранее Н.В. Прохоровой (1996), А.В. Поддубным (1999), В.Н. Матвеевым (2004), И.И. Костюкевичем и его соавторами (1991), где говорится о том, что содержание ТМ и Cs-137 в грибах увеличивается по мере улучшения лесорастительных условий произрастания грибов.