
Фауна, биология, экология трематод, развивающихся с участием пресноводных переднежаберных моллюсков приморского края 03. 00. 08 зоология 03. 00. 19 паразитология
| Вид материала | Автореферат диссертации |
| Глава 8. Хронология филогенеза трематод и формирование фаун E. beleocephalus S. newmillerdamensis P. oschmarini Глава 9 Природноочаговые трематодозы человека в Приморском крае |
- Фауна, биология и экология рода anopheles mg. (Сем. Culicidae) в прибрежных экосистемах, 1214.31kb.
- Департамент по тарифам приморского края, 19.48kb.
- 1 Разработка урока. Урок-игра. Телепередача "В мире животных. Моллюски" 2 Учебный предмет:, 47.42kb.
- Департамент имущественных отношений приморского края распоряжение, 17.48kb.
- «О внесении изменений в Закон Приморского края от 02 августа 2005 года №271-кз «О бюджетном, 12.13kb.
- Инструкция по организации антивирусной защиты в автоматизированных системах, предназначенных, 35.31kb.
- Федеральное агентство по рыболовству, 102.49kb.
- Фауна иксодовых клещей и эколого-биологические основы мер борьбы с ними в условиях, 823.3kb.
- Конституции Российской Федерации, федеральном закон, 1181.14kb.
- 1. Биология как наука, ее достижения, связи с другими науками. Методы изучения живых, 864.83kb.
Глава 8. Хронология филогенеза трематод и формирование фаун
Один из интересных и немаловажных вопросов в паразитологии - это вопрос формирования фаун трематод на отдельных территориях континентов, который, естественно, неразрывно связан с историей становления и развития взаимоотношений в системе трематоды-хозяева. К сожалению, эта область трематодологии находится в сфере гипотез, так как, в отличие от многих других животных, трематоды не оставили каких-либо палеонтологических следов. Различие же гипотез во многом зависит от позиций, которых придерживаются авторы в вопросах последовательности включения хозяев в жизненный цикл трематод и хронологии этого процесса. В настоящей главе, на основании результатов исследований, а также анализа литературных сведений по биологии трематод и сопоставления их с известными данными палеогеологии, предложен собственный взгляд на хронологию филогенеза и формирование современных фаун трематод.
8.1. Последовательность включения хозяев в филогенезе трематод
В целом, придерживаясь взглядов Т.А. Гинецинской (1968) по вопросу последовательности включения хозяев в жизненный цикл трематод, мы не исключаем и иные пути развития взаимоотношений в системе паразит - второй промежуточный и окончательный хозяева (Беспрозванных, 2002, 2004). Гинецинская (1968) предполагает, что первоначально сформировался диксенный цикл первый промежуточный – окончательный (позвоночные – рыбы) хозяева и только затем трематодами был освоен второй промежуточный хозяин (рис. 7 А). Включение в цикл окончательного хозяина происходило при случайном заглатывании позвоночными процеркарий (Синицын, 1910; Гинецинская, 1968). Если это так, то почему до момента их появления и расцвета это не происходило с беспозвоночными, которые могли и не случайно заглатывать процеркарий?
К.В. Галактионов и А.А. Добровольский (1998) объясняют невозможность подобного тем, что пероральный путь проникновения в беспозвоночных был непригоден для личинок протрематод, так как они были адаптированы к существованию в менее агрессивной среде (гемолимфа и т.д.), чем кишечная. Однако данное суждение вступает в противоречие со взглядами авторов на формирование диксении (пероральный путь проникновения в позвоночных). Не исключено, что свободноживущее личиночное поколение протрематод служило объектом питания для различных беспозвоночных. О такой возможности свидетельствуют современные данные (Шигин, 1981, 1981 и др.). Несомненно, большинство из них в кишечнике беспозвоночных погибало, однако также вероятно, что некоторые в результате так называемой «реакции бегства» избегали губительного действия пищеварительных ферментов животных. Мигрировав из кишечника, личинки попадали в другие ткани и органы, где, не найдя благоприятных условий для дальнейшего развития, инцистировались. Процесс образования прометацеркарий мог происходить и при случайном попадании процеркарий под раковину моллюсков, а при появлении рыбообразных – и в
 Рис. 6. Обнаружение трематод, найденных нами, и близких к ним видов (в скобках трематоды обнаруженные в Приморье) за пределами Дальнего Востока России.
Рис. 6. Обнаружение трематод, найденных нами, и близких к ним видов (в скобках трематоды обнаруженные в Приморье) за пределами Дальнего Востока России.Примечание: * - первые промежуточные хозяева.
слизи на поверхности тела и дыхательного аппарата. Возможно, процеркарии могли инцистироваться и во внешней среде, оказавшись в слизистых скоплениях, например, водорослей. Некоторые стилетные церкарии, хозяева которых - водные насекомые, способны инцистироваться в воде под стеклом или в сгустках слизи (Гинецинская, 1968; отмечалось также нами), а у стилетной церкарии Lecithodendriidae gen. sp. 1, способность инцистироваться в воде (вторично) стала нормой в развитии. Вряд ли способность процеркарий к инцистированию предопределилась освоением протрематодами окончательного хозяина (Гинецинская, 1968; Галактионов, Добровольский, 1998). Более логично, на наш взгляд, предположить приобретение процеркариями способности инцистирования до появления окончательного хозяина в цикле развития червей (рис. 7 Б).
До определенного момента этот путь в развитии был для большинства червей тупиковым, или, например, после гибели хозяина и распада его тканей они завершали свое развитие во внешней среде. Подобное демонстрируют трематоды Prosotocus confuses (Павлюк, 1972) и Isoparorchis hypselobagri (Беспрозванных, Ермоленко, 1989). Существование же протрематод в этот период обеспечивало свободноживущее гермафродитное поколение (Гинецинская, 1968). При таком развитии событий вовлечение позвоночных в циркуляцию червей могло осуществляться как при заглатывании ими процеркарий (например, как у представителей сем. Azygiidae) или проадолескарий, так и при питании животными, в (или на) которых могли быть заключены личинки
протрематод (рис. 7 Б). Преимущество при становлении системы трематоды–окончательные хозяева в большинстве случаев получают протрематоды, освоившие беспозвоночных и позвоночных первоначально в качестве, скажем так, «ненужных хозяев» в развитии свободноживущих червей. В дальнейшем у трематод, «отобранных» окончательными хозяевами, на стадии церкарии формируются приспособления к нахождению второго промежуточного хозяина и к проникновению через его покровы или естественные отверстия или к избирательному инцистированию на субстрате.
8.2. Хронология филогенеза трематод
Согласно имеющимся представлениям (Гинецинская, 1968), совместная эволюция протрематод и моллюсков берет свое начало в древнем палеозое (кембрий) в морской среде. Становление примитивного жизненного цикла трематод с участием двух хозяев моллюска и позвоночного датируется верхним триасом – ранней юрой и связывается с расцветом костистых рыб, а треххозяинного - в мелу (рис. 7 А), т.е. совпадает с периодом становления современных фаун пресноводных легочных и жаберных моллюсков (Старобогатов, 1970). Следовательно, эволюционирование фаун симбионтов происходит изолированно в силу появления непреодолимых преград, например, морские воды. В этих условиях маловероятно появление идентичных или сходных видов сосальщиков (рис. 6) на удаленных друг от друга территориях, в различных климатических поясах.
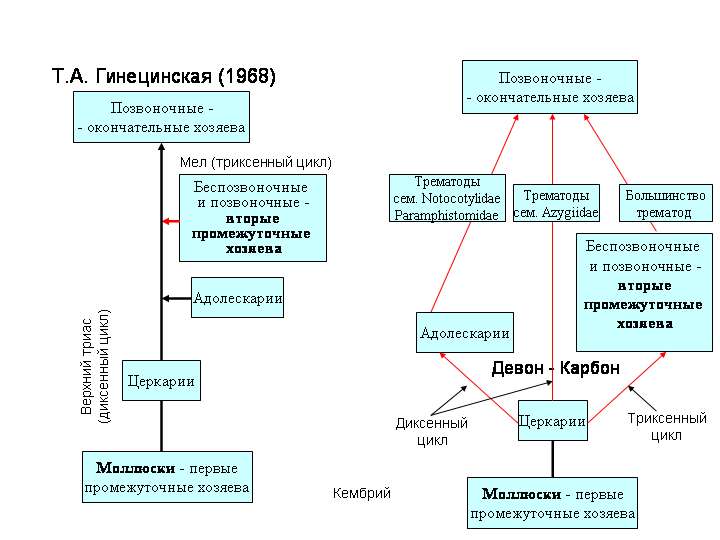
Рис. 7. Возможные пути становления циклов развития трематод: А – по Т.А. Гинецинской (1968); Б – наши представления.
Отправной точкой в восстановлении эволюционного пути организмов может служить какая-либо катастрофическая ситуация, ставящая на грань выживания изучаемый объект. В случае с трематодами - это переход в девоне-карбоне ряда моллюсков (их первых промежуточных хозяев) через амфибиотический к наземному образу жизни (Лихарев, 1965). Свободноживущие стадии трематод, изначально обитавшие в водной среде, оказались в ситуации, когда выживали только те из них, у которых сформировались приспособления к новым условиям жизни. Естественно, это не могло не сказаться отрицательно на биоразнообразии данной группы трематод. В настоящее время они объединены в надсемейство Brachylaimoidea Allison, 1943 и семейство Dicrocoeliidae Odhner, 1910.
Одна из важнейших морфологических структур большинства церкарий - локомоторный орган, обеспечивающий их перемещение в воде. Подавляющее число церкарий современных трематод из наземных легочных моллюсков имеет короткий хвост. Однако некоторые из них, например, Dicrocoelium lanceatum Stiles et Hassall, 1896, Brachylecithum americanum Denton, 1945 обладают хорошо развитым локомоторным органом, не несущим никакой функциональной нагрузки. Это может свидетельствовать о том, что трематоды попали из водной среды в несвойственную им наземную среду вместе с моллюсками, уже исполнявшими роль их первых промежуточных хозяев. Кроме того, сочетание у церкарий, например, Brachylecithum americanum, таких морфологических структур как длинный хвост и стилет – орган, изначально обеспечивавший проникновение церкарий через покровы второго промежуточного хозяина (церкарии дикроцелиид попадают во второго промежуточного хозяина алиментарным путем), позволяет предположить, что к моменту освоения моллюсками суши, развитие трематод осуществлялось с участием всех категорий хозяев. Для этого имелись все необходимые компоненты, способные обеспечить циркуляцию трематод, как в водной среде, так и на суше (Еськов, 2004). В современных условиях аналогично брахилоимоидеям и дикроцелиидам происходят сходные преобразования в онтогенезе микрофаллид, развивающихся с участием морских литоральных моллюсков (Галактионов, 1993).
Таким образом, можно предположить, что к моменту выхода моллюсков на сушу (девон - карбон) циркуляция трематод осуществлялась с участием всех категорий хозяев.
8.3. Формирование фаун трематод переднежаберных моллюсков
Как показано выше (рис. 6), в составе фаун сосальщиков длительно изолированных территорий имеются морфологически идентичные трематоды со сходной циркуляцией, развивающиеся с участием различных видов гастропод одного или разных семейств. В то же время, в популяциях близкородственных (в пределах семейства) гастропод, входящих в состав единого биогеоценоза, обнаруживаются морфологически сходные, но относящиеся к различным видам трематоды. Сходство проявляется или на стадии церкарии, или на всех этапах развития (таблица 6).
Согласно палеогеологии, в период поздний девон - карбон (когда, по нашему мнению, у трематод уже существовал треххозяинный цикл) суша представляла собой два гигантских материка - формирующиеся Лавразию и Гондвану, разделенных морским бассейном (Вегенер, 1984). Южное побережье Лавразии находилось в экваториальной зоне. Не исключено, что именно прибрежные воды, омывавшие южный берег Лавразии, - место формирования морской фауны брюхоногих, предшествовавшей современной пресноводной (в частности семейств Bithyniidae, Amuropaludinidae,
Bellamyidae и отряда Cerithiiformes), и, соответственно, фауны их паразитических симбионтов – трематод. В результате так называемых процессов – «экваториальной помпы» (Дарлингтон, 1966) и «зональной стратификации» (Жерихин, 1978; Еськов, 1984; Еськов, 2004) могло происходить расселение моллюсков и, соответственно, трематод, а в дальнейшем – вселение в континентальные водоемы и последующее формообразование. При этом для трематод не обязательно изменение морфологии. Хозяева и места их локализации, т.е. первичная среда, остаются неизменными, что не требует значительных перестроек уже морфологически адаптировавшихся к ней организмов. Необходимо только учитывать, что в условиях длительной изоляции морфологическое сходство трематод не всегда может гарантировать видовую идентичность. Это подтверждается и наличием видов-двойников среди трематод из моллюсков родов Boreoelona (сформировался как таковой на территории В. Сибири) и Parafossarulus (никогда не преодолевали северную границу их современного ареала – бассейн р. Амур), имеющих сино-индийские корни и обитающих ныне совместно (таблица 6).
Таблица 6
Трематоды моллюсков Boreoelona и Parafossarulus из водоемов Приморья, имеющие морфологическое сходство на стадиях онтогенеза
| Вид | Первый промежуточный хозяин | Вид | Первый промежуточный хозяин |
| E. beleocephalus | Boreoelona spp. | E. japonicus | Parafossarulus spp. |
| Opistorchidae gen. sp. 1 | - // - | Opistorchidae gen. sp. 2 | - // - |
| Maritrema sp. 1 | - // - | Maritrema sp. 2 | - // - |
| S. newmillerdamensis | - // - | S. monorchis | - // - |
| Sanguinicola sp. 1 | - // - | Sanguinicola sp. 2 | - // - |
| P. oschmarini, P. anserinum | - // - - // - | P. acutirostris | - // - |
Такое представление развития событий в той или иной степени может объяснить полигостальность трематод к первому промежуточному хозяину среди обнаруженных сосальщиков (см. гл. 7, рис. 6).
Глава 9 Природноочаговые трематодозы человека в Приморском крае
До недавнего времени считалось, что на территории Дальнего Востока России из рода Metagonimus распространены только трематоды вида M. yokogawai. Впервые нами в данном регионе был обнаружен второй вид этого рода - M. katsuradai (Беспрозванных и др., 1987). Также впервые на территории России зарегистрирована трематода Centrocestus armatus (Беспрозванных, 1988), возможность заражения человека которой установлена экспериментально (Tanabe, 1922). Получены данные по биологии и распространению трематод C. sinensis, M. yokogawai, M. katsurada, C. armatus, N. salmincola schikhobalowi и P. westermani ichunensis. В последнем случае экспериментально установлено, что обе формы парагонимоза - легочная и мышечная (ларвальная) вызываются одним и тем же подвидом – P. westermani ichunensis. Установлены причины, при которых может развиваться та или иная форма заболевания (Беспрозванных, 1994, 2002, 2004).
Впервые обобщены собственные и литературные данные по природноочаговым трематодозам человека в Приморье, возбудителями которых являются трематоды M. yokogowai, M. katsuradai, C. armatus, N. salmincola, P. westermani, C. sinensis и осуществлено их картирование (Беспрозванных, Ермоленко, 2005).
Выводы
1. В Приморском крае циркулирует 72 вида трематод 35 родов из 22 семейств, развивающихся с участием первых промежуточных хозяев пресноводных переднежаберных моллюсков. Наибольшее количество видов трематод относится к семейству Pleurogenidae (9 видов), к остальным семействам: Echinostomatidae (8), Psilostomidae (6), Heterophyidae, Lecithodendriidae, Notocotylidae, Cyathocotylidae (по 5), Plagiorchidae (4), Sanguinicolidae, Microphalidae, Opisthorchidae, Monorchidae, Strigiidae (по 3), Prosthogonimidae (2), Nanophyetidae, Eumegacetidae, Phylophthalmidae, Opecoelidae, Paragonimidae, Azygiidae, Isoparorchidae, Halipegidae (по 1 виду). В числе обнаруженных трематод новые для науки - 8 видов, для фауны России – 16 видов и для Приморья – 7 видов.
2. Изучены жизненные циклы 39 видов трематод (20 - впервые, 12 - впервые на территории России и 7 – впервые для Приморья), для 13 видов выявлены вторые промежуточные хозяева и для обнаруженных трематод определено 17 путей циркуляции в условиях Приморского края. Установлено, что наибольшая роль в циркуляции трематод среди первых промежуточных хозяев принадлежит моллюскам рода Parajuga (28 видов трематод), а среди вторых–третьих промежуточных и окончательных хозяев – рыбам (30 видов трематод). Определено, что у трематод из моллюсков рода Parajuga (обитатели рек) среди вторых промежуточных хозяев доминируют рыбы и насекомые (поденки, веснянки и ручейники), а среди окончательных - эта роль равномерно распределена между рыбами, птицами и млекопитающими. У трематод из моллюсков родов Parafossarulus, Boreoelona и Cipangopaludina (обитатели пойменных водоемов) соответственно со сходной частотой, в циркуляции участвуют рыбы, насекомые (преимущественно стрекозы), моллюски, и много видов, инцистирующихся в воде, а среди окончательных хозяев преобладают птицы. Такие различия в составе хозяев трематод предопределены давностью становления жизненных циклов сосальщиков и разновременностью установления топических и трофических связей между их хозяевами.
3. Определено, что для большинства обнаруженных видов трематод характерна триксенния, для тринадцати видов - диксенния в жизненном цикле. У четырех видов сосальщиков выявлены приспособления к сокращению числа обязательных хозяев в цикле развития. Для трематод Echinochasmus sp., E. beleocephalus и Mosesia okabei отмечена необлигатная амфиксения на стадии церкарии, а трематода Isoparorchis hypselobagri при облигатной тетраксении способна развиваться по триксенному типу. В цикл развития Paragonimus westermani ichunensis вовлекаются нетипичные для паразита окончательные хозяева. Таким образом, наряду со сложившимися путями развития трематод выявляются тенденции в эволюции системы трематоды-хозяева, проявляющиеся в формировании у сосальщиков новых адаптаций для реализации жизненного цикла. Это, с одной стороны, обеспечивает сохранение вида, с другой – создает основу становления новых путей циркуляции и видообразования.
4. Изучены особенности биологии церкарий 53 видов трематод 18 семейств, что позволило вычленить группы трематод, характеризующиеся сходным комплексом адаптаций к заражению второго хозяина. Отмечено, что церкарии различной систематической принадлежности, использующие одних и тех же или близких по эколого-биологическим характеристикам животных, обладают сходными суточным ритмом выхода из моллюска и таксисами. В состав современных гемипопуляций церкарий входят особи, в большинстве обладающие характерными для вида адаптациями, позволяющими им заражать облигатного хозяина, а также особи с иной приспособленностью, что дает трематодам возможность расширить круг хозяев и, соответственно, выжить в условиях изменяющейся среды. В некоторых случаях успех реализации жизненного цикла трематод обеспечивается за счет вторичного изменения у церкарий приспособлений к проникновению во второго промежуточного хозяина.
5. В зараженности партенитами трематод пресноводных переднежаберных моллюсков разных экологических группировок выявлен ряд закономерностей. Показано, что зараженность моллюсков находится в зависимости от соотношения числа яиц паразита в стации и количества его первых промежуточных хозяев. Степень инвазированности моллюсков зависит от возрастного состава и особенностей поведения особей, входящих в состав их популяций. Паводки и падения уровня воды в водоеме влияют на численность и возрастной состав популяции первых промежуточных хозяев и, соответственно, на показатели их зараженности. Определено, что скорость течения не оказывает влияния на зараженность моллюсков рода Parajuga, а участки реки с галечниковым дном, в сравнении с илистым, наиболее благоприятны для трематод при заражении моллюсков данного рода.
6. Из обнаруженных 72 видов трематод 26 способны развиваться с участием различных видов моллюсков рода Parajuga (сем. Pachychilidae, отр. Cerithiiformes), 8 – Boreoelona и 3 – Parafossarulus (сем. Bithyniidae). Трематоды 18-ти видов разными авторами найдены на стадии партенит у моллюсков других родов и семейств отряда Cerithiiformes (8 видов) и видов и родов семейства Bithyniidae (10 видов) за пределами Приморского края. Кроме того, трематоды 10 видов на стадии мариты были зарегистрированы на территориях, где отсутствуют гастроподы родов Parajuga, Boreoelona и Parafossarulus. Это противоречит общепринятым представлениям о моногостальности трематод в отношении первого промежуточного хозяина. Отсутствие достаточных знаний о фаунах и путях циркуляции трематод в различных регионах континентов, истории развития взаимоотношений в системе трематоды-хозяева, а также о молекулярно-генетических особенностях животных, составляющих данную систему, не позволяют в настоящее время говорить о причинах моно- и полигостальности отдельных видов сосальщиков на этапе развития в первом промежуточном хозяине.
7. Согласно господствующим представлениям диксения в циркуляции трематод (первый промежуточный – окончательный хозяева) датируется триасом, а триксения (вовлечение в циркуляцию трематод второго промежуточного хозяина) – меловым периодом. Анализ результатов исследований и литературных сведений по биологии трематод и сопоставление их с известными данными палеогеологии позволили сформулировать гипотезу, в соответствии с которой освоение окончательных хозяев протрематодами происходило одновременно на стадиях процеркарии, проадолескарии и прометацеркарии, и жизненный цикл трематод уже в период конец девона - карбон мог осуществляться с участием всех категорий хозяев. Исходя из времени становления жизненного цикла трематод - девон-карбон, расположения в этот период материков и современного распространения идентичных сосальщиков и их первых промежуточных хозяев определен центр формирования первичной фауны трематод переднежаберных моллюсков - южное побережье Лавразии. Из этого района, вместе с первыми промежуточными хозяевами, произошло расселение трематод вдоль материковых побережий, а затем независимое освоение континентальных водоемов и формирование современных фаун трематод (юра-мел). Это в конечном итоге привело к возникновению дезъюнктивных ареалов у ряда видов сосальщиков.
8. Из регионов России Приморский край – территория, где фауна трематод, развивающихся с участием первых промежуточных хозяев - пресноводных переднежаберных моллюсков, представлена наибольшим числом видов, способных паразитировать у человека. Выявлено 6 видов трематод (Metagonimus yokogawai, M. katsuradai, Centrocestus armatus, Paragonimus westermani ichunensis, Nanophyetus salmincola schikhobalowi и Clonorchis sinensis) - возбудителей заболеваний человека. Определены пути их циркуляции и распространение, а также установлено, что легочная и мышечная формы парагонимоза вызываются трематодой P. westermani ichunensis. Эти данные, а также проведенное картирование трематодозов - основа для разработки профилактических мероприятий, одним из компонентов которых должны быть мониторинговые исследования природноочаговых трематодозов в Приморском крае.