
Фауна, биология, экология трематод, развивающихся с участием пресноводных переднежаберных моллюсков приморского края 03. 00. 08 зоология 03. 00. 19 паразитология
| Вид материала | Автореферат диссертации |
| Глава 6. Зависимость заражения моллюсков от различных биотических и абиотических факторов 6.1.2. Характер грунта. 6.1.3. Уровень воды. 6.1.4. Размер стации. |
- Фауна, биология и экология рода anopheles mg. (Сем. Culicidae) в прибрежных экосистемах, 1214.31kb.
- Департамент по тарифам приморского края, 19.48kb.
- 1 Разработка урока. Урок-игра. Телепередача "В мире животных. Моллюски" 2 Учебный предмет:, 47.42kb.
- Департамент имущественных отношений приморского края распоряжение, 17.48kb.
- «О внесении изменений в Закон Приморского края от 02 августа 2005 года №271-кз «О бюджетном, 12.13kb.
- Инструкция по организации антивирусной защиты в автоматизированных системах, предназначенных, 35.31kb.
- Федеральное агентство по рыболовству, 102.49kb.
- Фауна иксодовых клещей и эколого-биологические основы мер борьбы с ними в условиях, 823.3kb.
- Конституции Российской Федерации, федеральном закон, 1181.14kb.
- 1. Биология как наука, ее достижения, связи с другими науками. Методы изучения живых, 864.83kb.
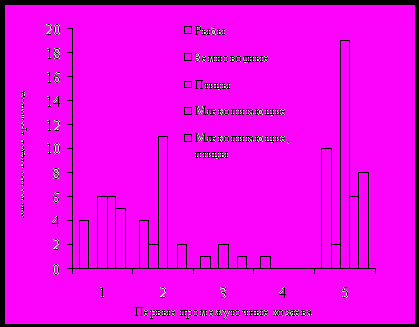
Рис. 2. Количество трематод развивающихся с участием определенных первых промежуточных и окончательных хозяев: 1 – Parajuga, 2 – Boreoelona-Parafossarulus, 3 – Cipangopaludina-Amuropaludina, 4 – Cincinna, 5 – Prosobranchia.
участвуют рыбы, насекомые и моллюски, а также имеется большое количество видов, инцистирующихся в воде. При этом среди насекомых-хозяев превалируют стрекозы, имеющие личинок с ректальной дыхательной системой. Такие личинки стрекоз обычны и многочисленны в стациях обитания, свойственных моллюскам родов Boreoelona, Parafossarulus и Cipangopaludina. В тоже время в русловой части реки, где обитают моллюски рода Parajuga, они малочисленны. В условиях реки доминируют поденки, веснянки и ручейники. Это относится и к земноводным, которые исполняют роль хозяев для трематод из моллюсков родов Boreoelona и Parafossarulus.
Такие различия в составе вторых, третьих промежуточных и окончательных хозяев трематод результат разновременности установления топических и трофических связей между животными, которые обеспечивают сосальщикам возможность циркуляции.
5.2. Некоторые аспекты взаимоотношений в системе паразит – хозяин
На примере некоторых видов трематод показано, что их потенциальные возможности паразитирования на стадиях метацеркарии и мариты у животных различных систематических групп шире, чем реализуются в природе. В экспериментах заражались те животные, у которых в естественных условиях трематоды определенного вида или не обнаруживались или случаи их нахождения были редки.
Глава 6. Зависимость заражения моллюсков от различных биотических и абиотических факторов
Имеющиеся в литературе сведения (Гинецинская, 1968; Галактионов, 1982; Галактионов, Русанов, 1983; Рыбаков, 1984; Арыстанов, 1986; Галактионов, Марасаев, 1986; Галактионов, Михайлов, 1987, 1990; Галактионов, 1993 и др.) о влиянии биотических и абиотических факторов на зараженность гастропод получены в основном при исследовании морских, солоноватоводных и пресноводных моллюсков из пойменных водоемов и устьевых частей рек. В то же время, экологические аспекты заражения трематодами моллюсков – обитателей русловой части среднего и верхнего течения рек практически не изучались. Такие наблюдения были проведены нами (Беспрозванных, 1990, 1999, 2000) в бассейне реки Раздольной, включающей в себя полугорные и равнинные участки и равномерно, по всей длине, подверженной воздействию антропогенного фактора. Река населена тремя видами моллюсков рода Parajuga, которые характеризуются сходной экологией, размерами и восприимчивостью к заражению трематодами (P. subtegulata - 20 видов трематод; P. subextensa - 23; P. subcalculus - 15). Наряду с этим, осуществлены исследования некоторых аспектов заражения различных видов переднежаберных моллюсков из пойменных водоемов.
6.1. Проточность водоема и характер стации обитания.
6.1.1. Проточность. Известно, что с увеличением проточности водоема экстенсивность инвазии моллюсков уменьшается (Черногоренко-Бидулина, 1958; Куприянова-Шахматова, 1961; Бутенко, 1967; Гинецинская, 1968). Однако большинством авторов исследовалось влияние проточности на зараженность моллюсков - обитателей пойменных водоемов. Гастроподы рода Parajuga - типичные донные гидробионты рек.
В реке Раздольной при скорости течения 0.8 м/сек, экстенсивность инвазии моллюсков достигала 35%, а при 0.2 - 0.3 м/сек - 7-16.9%.
Основная масса гастропод концентрируется на глубине до 70 см, где и происходит их заражение. В этом случае именно те трематоды, которые на стадии яйца вместе с фекалиями попадают в прибрежную зону, имеют наибольшие шансы для осуществления жизненного цикла. Распределение скоростей течения в реке прямо пропорционально глубине, следовательно, яйца трематод, попавшие в жизненную зону гастропод рода Parajuga, оказываются в условиях малой скорости течения и перехода турбулентного движения воды в ламинарное. В такой ситуации скорость течения не оказывает какого-либо заметного влияния на зараженность гастропод, что подтверждается высокой зараженностью партенитами трематод моллюсков рода Parajuga.
6.1.2. Характер грунта. Данные о влиянии структуры грунта на зараженность моллюсков трематодами немногочисленны. Имеются сообщения о большей инвазированности моллюсков рода Littorina обитающих на каменистых россыпях, чем на илисто-песчанном грунте (Чубрик, 1966, Галактионова Михайлов, 1990 и др.). Сходные результаты получены и нами при исследовании зараженности моллюсков рода Parajuga, обитающих на участках реки Раздольной с галечниковым и илистым дном (Беспрозванных, 2000). В первом случае экстенсивность инвазии гастропод достигала 35%, во втором - 7%. По всей видимости, в условиях илистого дна уменьшается вероятность контакта моллюсков с яйцами или мирацидиями трематод, находящимися под постоянным прессом оседающих твердых частиц. Не исключено, что данный фактор значим и при заражении представителей родов Cipangopaludina (только изредка поднимаются в верхние слои воды по водной растительности) и Amuropaludina, которые, как и Parajuga - донные обитатели. Однако повсеместно низкий процент их зараженности не позволяет получить данные, подтверждающие или опровергающие зависимость их зараженности от характера грунта. К тому же, виды рода Cipangopaludina предпочитают в своей жизнедеятельности пойменные стоячие водоемы, дно которых обычно покрыто толстым слоем ила. Что касается гастропод родов Boreoelona и Parafossarulus - обитателей стоячих водоемов, живущих в прибрежных участках на обильной водной растительности, то источником их заражения, вероятнее всего, служат яйца трематод, оседающие вместе с фекалиями на растения. Таким образом, состав грунта в данном случае не оказывает заметного влияния на зараженность моллюсков, как это наблюдается у видов рода Parajuga.
6.1.3. Уровень воды. Определенное влияние на инвазированность моллюсков оказывает и изменение уровня воды в водоеме. Наблюдения, проведенные в течении 2-х беспаводковых лет (2001-2002 гг.) в бассейне реки Арсеньевки, позволили выявить стабильность в зараженности, возрастном составе и плотности поселения популяций моллюсков родов Cipangopaludina, Parafossarulus и Boreoelona (таблица 3). Первый же весенний паводок, охвативший практически всю пойму реки, привел к значительным изменениям как в зараженности, так и в популяционной структуре сообществ моллюсков (таблица 3). При этом в первую очередь он оказал катастрофическое воздействие на гастропод родов Parafossarulus и Boreoelona. В силу особенностей их морфологии (малые размеры), функциональности (медлительность) и экологии (большую часть жизни проводят в толще стоячей воды на водной растительности) эти гастроподы наиболее подвержены воздействию потока воды. По всей видимости, большинство особей из популяций были смыты сильным течением, возникшим при подъеме уровня воды и соединении старицы с рекой. Отдельные экземпляры этих моллюсков после завершения паводка встречались на прибрежной водной растительности в русле реки.
Иная ситуация с представителями рода Cipangopaludina - донными гидробионтами. При повышении уровня воды моллюски мигрируют в направлении привычной для жизнедеятельности глубины (от уреза воды до глубины 1.5 м), в результате чего способны за время разлива переместиться на значительное расстояние от исходной стации обитания. Впоследствии многие Cipangopaludina (в первую очередь, менее подвижные, с высотой раковины 17-30 мм), как и другие виды моллюсков, не успевают за происходящим быстрым падением уровня воды и погибают (обнаруживались в большом количестве после схода воды вдали от водоемов).
В послепаводковый период значительно уменьшается плотность поселения моллюсков, и в популяциях преобладают особи I-II-ой размерных групп (исключение - Cipangopaludina). Экстенсивность инвазии гастропод уменьшается, так как сокращается число обычно наиболее зараженных моллюсков III-ей размерной группы. Что касается представителей рода Cipangopaludina, то среди них выжили наиболее крупные особи, что и предопределило повышение зараженности в 2003 году (таблица 3).
В случае с моллюсками рода Parajuga воздействие рассматриваемого абиотического фактора не столь выражено как в случае с моллюсками родов Parafossarulus, Boreoelona и Cipangopaludina. В период паводка поведение гастропод рода Parajuga соответствует поведению представителей р. Cipangopaludina. После паводка (средний участок реки Раздольной) в лужах нами насчитывалось от 50 до 300 моллюсков рода Parajuga, причем часть из них извлекалась из толщи почвы. Возрастной состав таких изолированных групп моллюсков неоднороден. В среднем (обследовано 5 изолятов) до 78 % составляли особи с высотой раковины 17 - 20 мм, 15% - 23-28 мм, а остальные - менее 17 мм. Экстенсивность инвазии представителей всех размерных групп составляла 3.7%, что близко к показателю инвазированности моллюсков на данном участке реки (4.5%). По завершении паводка заметных изменений, как в соотношении особей различных размерных групп, так и в показателях зараженности моллюсков не наблюдалось. Это, по всей видимости, связано, с одной стороны, с большой численностью гастропод (более 200 особей на 1 м2), с другой - перераспределением их по руслу реки в результате обратной миграции в стацию по мере снижения уровня воды, а также неизбирательной гибелью как зараженных, так и стерильных моллюсков.
Исходя из изложенного выше, время паводка - это период, когда минимальна вероятность заражения трематодами, как первых, так и вторых промежуточных хозяев. С началом паводка гастроподы мигрируют из стаций обитания в направлении малых глубин, переходят на залитые участки суши, не содержащие яиц трематод. Последующее снижение объема стока приводит к возвращению мягкотелых в русло реки или пойменные водоемы, очищенные от инвазионного начала вследствие смывания и «погребения» яиц трематод. После паводка относительная зараженность гастропод может претерпеть значительные изменения по сравнению с допаводковым периодом или остаться неизменной. Фактически же количество инвазированных моллюсков (наряду со свободными от трематод) снижается за счет их гибели.
На зараженность моллюсков оказывает влияние и падение уровня воды в засушливые годы (2003-2004 гг., бассейн реки Арсеньевки). При малой плотности поселения гастропод родов Boreoelona и Parafossarulus наблюдалось увеличение их зараженности (таблица 3). Это, вероятнее всего, связано с тем, что в маловодных условиях немногочисленные выжившие моллюски распределяются на глубине 10-30 см (при стабильных - на глубине 60-70 см). Образовавшееся мелководье обеспечивает большую его доступность для различных околоводных животных и, соответственно, высокую концентрацию яиц трематод на 1 м2 в зоне жизнедеятельности моллюсков, а хорошая прогреваемость воды - интенсивное развитие мирацидиев. В тоже время снижение
Таблица 3
Зараженность моллюсков в условиях изменения уровня воды в водоеме
| Моллюски | Показатели за 2001-2002 гг. (до половодья) | Показатели за май 2003 г (после весеннего половодья) | Показатели за июнь 2004 г. (после маловодного летнего периода 2003) | |||||||||
| 1* | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | ||||
| а | б | а | б | а | б | |||||||
| Cipangopaludina sp. | 10 | 30 | 70 | 1.6 | 0-3 | 3 | 97 | 5.2 | 0-1 | 100 | 0 | 0 |
| Boreoelona sp. | 57 | 40 | 60 | 2.3 | 10 | 90 | 10 | 1.2 | 7 | 72 | 28 | 19.4 |
| Parafossarulus sp. | 25 | 25 | 75 | 2.7 | 0-3 | 85 | 15 | 1.0 | 0-3 | 18 | 82 | 25.0 |
Примечание: * - показатели в 1 и 2 колонках не абсолютные 1 - плотность поселения м2, 2 - размерный состав особей популяции % (а – Cipangopaludina sp. 17-30 мм, Boreoelona sp. до 4 мм, Parafossarulus sp. до 5 мм; б - Cipangopaludina sp. 35-50 мм, Boreoelona spp. <4-8 мм, Parafossarulus spp. <5-14 мм); 3 – зараженность %.
уровня и высокая температура (до 30о С) воды в водоеме отрицательно сказываются на жизнеспособности гастропод рода Cipangopaludina, в первую очередь, старшей возрастной группы (основная роль в поддержании очагов инвазии) и, как следствие - нулевые показатели зараженности моллюсков (таблица 3).
Таким образом, паводок и межень – это факторы среды, оказывающие значительное влияние не только на количество зараженных моллюсков, но и на численность особей в популяциях, как промежуточных хозяев трематод, так и трематод в целом, что неизбежно должно отразиться и на последующих поколениях животных.
6.1.4. Размер стации. Очаговость заражения моллюсков отмечалась в многочисленных работах (Фролова, 1958; Гинецинская, Штейн, 1961; Гинецинская, 1968; Черногоренко, 1983; Арыстанов, 1986 и др.) как фактор, характерный, в первую очередь, для больших водных акваторий. Кроме того, отмечается, что с увеличением площади водоема снижаются средние показатели зараженности первых промежуточных хозяев и видовое разнообразие трематод.
Полученные нами данные свидетельствуют о наличии в пределах как крупных, так и средних водоемов (озеро, старица, река) очагов инвазий, обусловленных как отдельно взятым видом трематод, так и их совокупностью.
Результаты исследований показывают, что взаимосвязь паразита и хозяина, проявляющаяся в изменении зараженности первого промежуточного хозяина в зависимости от размеров стации обитания, зависит от ряда факторов: плотности поселения хозяев, распределения по ее периметру промежуточных и окончательных хозяев и частотой посещения водоема последними. Исходя из этого, данная зависимость не может быть однонаправленной, и в каждом отдельно рассматриваемом случае будет определяться особенностями биогеоценоза.
6.2. Плотность поселения популяции позвоночных и беспозвоночных
Основная часть исследований проведена в бассейне реки Раздольной, как уже упоминалось выше, населенной тремя видами моллюсков рода Parajuga, и в отсутствии локальных скоплений окончательных хозяев. О более или менее равномерном их распределении свидетельствует качественно сходный состав фауны трематод моллюсков рода Parajuga на обследованных участках реки.
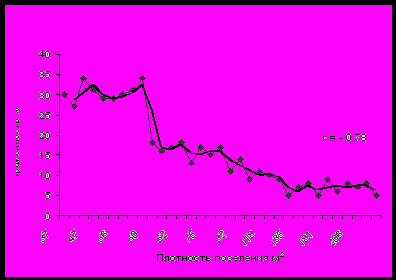
Рис. 3. Зависимость зараженности моллюсков рода Parajuga от плотности их поселения в реке Раздольной
Обычно считается, что увеличение плотности популяции хозяина безусловно приводит к увеличению экстенсивности инвазии его паразитами. Данное положение справедливо для паразитов с прямым жизненным циклом. Взаимоотношения в системе паразит-хозяин среди поликсенных видов сложнее. Прямое увеличение зараженности первых промежуточных хозяев будет наблюдаться только при неизменной плотности их поселения и, соответственно, повышении численности последующих хозяев в цикле развития трематод. В данном случае зараженность первых промежуточных хозяев будет ограничена 100% порогом. Инвазированность моллюсков при разной плотности их поселения также будет ограничена какой-либо пороговой величиной, если численность вторых промежуточных и дефинитивных хозяев стабильна. Однако она не будет находиться в прямой зависимости от плотности поселения гастропод. Вначале, с увеличением таковой, происходит повышение зараженности первых промежуточных хозяев (Зеликман, 1954; Фролова, 1964; Рыбаков, 1984; Арыстанов, 1986 и др.). После того, как численность моллюсков превысит величину, обеспечивающую максимально возможное использование наличного инвазионного материала, их относительная зараженность будет снижаться (Ewers, 1964, Беспрозванных, 1999, 2000). Снижение зараженности гастропод с увеличением плотности их поселения наблюдалось нами в реке Раздольной (рис. 3). Данная зависимость сохранится и при одновременном изменении численности всех категорий хозяев. Будет лишь происходить изменение пороговой величины и смещение графика по оси абсцисс.
6.3. Особенности биологии трематод
Установлено, что для трематод, развивающихся с участием первых промежуточных хозяев - моллюсков рода Parajuga, характерна утрата свободноживущего мирацидия (Беспрозванных, 2000). У 14 из 28 видов трематод, обнаруженных у моллюсков рода Parajuga, мирацидии не выходят из яиц во внешнюю среду. Зараженность первых промежуточных хозяев такими трематодами достигает 22.6% (от общего числа исследованных гастропод), тогда как остальными - только 7.2%. Выделенные трематодами яйца оседают вместе с фекалиями хозяев на дно реки, где и заглатываются моллюсками рода Parajuga, питающимися детритом. Отсутствие непосредственного влияния внешних условий на развитие мирацидиев, а после попадания яиц в воду - на личинку, защищенную оболочкой яйца, и образ жизни моллюсков данного рода обеспечивают большую зараженность первых промежуточных хозяев трематодами, у которых нет свободноплавающего мирацидия. В условиях реки - постоянного тока и перемешивания воды, как и в условиях морской литорали (Цимбалюк, 1975, Галактионов 1987 и др.), преимущество получают трематоды, у которых нет свободноплавающего мирацидия.
6.4. Возраст моллюска
Полученные нами данные по зараженности моллюсков родов Parajuga (рис. 4) и Boreoelona, Parafossarulus, Cipangopaludina (таблица 4) в целом свидетельствуют о прямой зависимости зараженности моллюсков от их размеров. В то же время, в зараженности разноразмерных гастропод рода Parajuga, собранных на отдельных участках реки Раздольной, выявлены некоторые особенности. Если на среднем участке она выражается в постепенном увеличении инвазированности моллюсков с возрастом, то на двух других, по достижении гастроподами 20 мм на верхнем и 25 мм на нижнем участках, она резко возрастает. Распределение разноразмерных моллюсков по глубинам различно (таблица 5). Гастроподы с высотой раковины 7-17 мм, как на отдельных участках реки, так и в целом, тяготеют к малым глубинам и ведут малоподвижный образ жизни, чем объясняются низкий процент и сходство в их зараженности (верхний участок - экстенсивность инвазии 2.9%, средний - 1.3%, нижний - 1.0%). Что касается гастропод с высотой раковины <17-30 мм, то они расположены на отдельных участках неодинаково. На среднем участке такие моллюски преобладают на глубине 0-35 см и ведут образ жизни, сходный с таковым юных
особей (малоподвижны, часто зарываются в ил – дно илистое), что обуславливает как постепенное, равномерное увеличение зараженности гастропод с возрастом, так и низкую инвазированность крупных экземпляров.
На нижнем (дно песчаное) и верхнем (дно галечниковое) участках реки моллюски с высотой раковины <17-30 см рассеяны по глубинам более равномерно, что свидетельствует о большей их подвижности. Это в свою очередь увеличивает вероятность контакта гастропод с яйцами трематод (см. раздел 6.3) и, как следствие, большая их зараженность по сравнению с моллюсками со среднего участка.
Таблица 4
Зараженность разноразмерных моллюсков (%)
| Род моллюсков | Размерные группы | |
| I-II | III | |
| Boreoelona | 5.6 | 7.1 |
| Parafossarulus | 1.0 | 9.2 |
| Cipangopaludina | 1.7 ( I ) | 9.0 (II) |
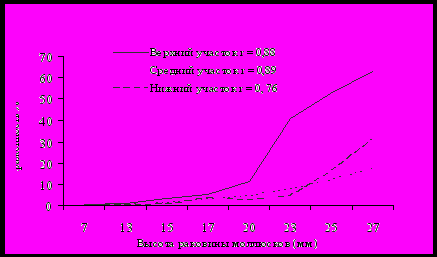
Рис. 4. Зараженность разноразмерных моллюсков рода Parajuga из реки Раздольной
Таблица 5
Распределение разноразмерных моллюсков рода Parajuga по глубинам на м2
| Участок реки | Плотность поселения моллюсков (м2) на глубине 0 - 35 см | Плотность поселения моллюсков (м2) на глубине 35 - 70 см | ||
| 7 - 17 мм | <17 - 30 мм | 7 - 17 мм | <17 - 30 мм | |
| Верхний | 37 | 33 | 14 | 28 |
| Средний | 248 | 69 | 122 | 29 |
| Нижний | 32 | 31 | 23 | 38 |
Таким образом, наряду с общезакономерным увеличением зараженности моллюсков по мере их роста, инвазированность разноразмерных гастропод зависит от их поведения в конкретных условиях.
6.5. Сезонная зараженность первых промежуточных хозяев трематод и ее зависимость от возрастной структуры популяции моллюсков
Обычно сезонная зараженность первых промежуточных хозяев выражается одним (чаще) или двумя пиками в зараженности моллюсков, приходящимися на середину лета - начало осени (Гинецинская, 1959; Куприянова-Шахматова, 1961; Чубрик, 1966; Черногоренко, 1983 и др.). Как указывает Т.А. Гинецинская (1968), данная закономерность обусловлена комплексом факторов, один из которых - возрастной состав популяции моллюсков, в свою очередь, зависящий от ряда биотических и абиотических факторов.
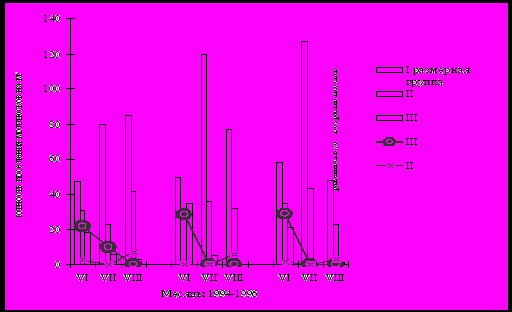
Рис. 5. Размерная структура популяции моллюсков рода Boreoelona и их сезонная зараженность
Изложенные выше результаты исследований свидетельствуют о повышении инвазированности моллюсков по мере увеличения их размеров. В этом случае естественные сезонные изменения возрастного состава популяций моллюсков не могут не сказываться на показателях их зараженности трематодами. Наличие данной зависимости подтверждаются наблюдениями за взаимоотношениями, складывающимися на уровне популяций двух видов трематод Nenimandijea kashmirensis и Pleurogenoides medians (паразиты лягушек) и их первого промежуточного хозяина - Boreoelona ussuriensis. На протяжении исследований популяция мягкотелых включала представителей всех возрастов (исключая сеголетков), но в различном количественном соотношении, как в целом, так и в отдельные периоды вегетации (рис. 5), что связано с появлением молоди и гибелью моллюсков, достигших предельного возраста. Наивысшие показатели экстенсивности инвазии моллюсков партенитами N. kashmirensis и P. medians отмечались в мае–июне и достигали 4.3%. При этом, наибольшую долю среди инвазированных моллюсков составляли представители III размерной группы - 21.7-29% (зараженность среди особей данной группы). В июле общая экстенсивность инвазии моллюсков снижается до минимума, что, в первую очередь, связано со снижением численности особей III размерной группы. В дальнейшем, в конце августа – начале сентября, происходит повышение зараженности первых промежуточных хозяев, которое обусловлено увеличением числа инвазированных моллюсков II размерной группы. Максимального значения этот процесс достигает в мае–июне следующего года. В июле–августе происходит массовый выплод молоди моллюсков, которые могут составлять до 76% особей популяции. Пополнение популяции за счет стерильных сеголетков, естественно, снижает относительные показатели зараженности моллюсков, но никак не сказывается на фактическом количестве инвазированных мягкотелых. Стабильные сезонные изменения возрастного состава популяции моллюсков B. ussuriensis обуславливали и сходные сезонные изменения в их зараженности трематодами на протяжении 1994-1996 гг.
6.6. Случаи множественных заражений моллюсков партенитами трематод
Множественные заражения гастропод различными видами трематод неоднократно регистрировались как у пресноводных (Cort, McMullen and Breackett, 1937; Черногоренко-Бидулина, 1958; Гинецинская, 1959; Здун, 1961; Никитина, 1986), так и у солоноватоводных и морских моллюсков (Зеликман, 1954; Рыбаков, 1984; Мехралиев, 1993). В наших исследованиях сопаразитирование различных видов партенит было отмечено у 3.7% моллюсков рода Parajuga от числа зараженных. Чаще других одним из сопаразитов были партениты вида Holosthephanus sp.- 86% (от числа моллюсков, зараженных несколькими трематодами). Наиболее распространено сопаразитирование пар Holostephanus sp. + Nanophyetus salmincola schikhobalowi (37%), Holostephanus sp. + Prosthodendrium dollfusi (27.3%) и Holostephanus sp. + Mosesia longicirrus (17.6%). Тройная инвазия моллюсков партенитами была отмечена один раз. Сопаразитирование партенит с метацеркариями представлено у моллюсков рода Parajuga только в комбинации партениты + метацеркарии Sphaeridiotrema spinoacetabulum и Euparyphium amurensis.
В отличие от моллюсков рода Parajuga, среди представителей родов Boreoelona, Parafossarulus, Cincinna и Amuropaludina сопаразитирование партенит не выявлено, у Cipangopaludina в одном случае обнаружено три вида трематод – Azygia hwangtsiytii + Paracoenogonimus ussuriensis + Pleurogenidae gen. sp. 1. Неиспользование перечисленных моллюсков в качестве хозяина одновременно несколькими трематодами обусловлено, по всей видимости, рядом причин. Одна из них - ограниченность среды (малые размеры пищеварительной железы моллюсков Boreoelona, Parafossarulus и Cincinna) для развития и функционирования партенит. Что касается Amuropaludina и Cipangopaludina (крупные моллюски), то обнаружение у одного из них нескольких видов совместно паразитирующих трематод исключает факт ограниченности среды для паразита. В данном случае первостепенное значение имеет видовое разнообразие трематод и насыщенность внешней среды яйцами сосальщиков в местах обитания этих моллюсков (возможно, для Boreoelona, Parafossarulus и Cincinna это также имеет значение). Экстенсивность инвазии моллюсков рода Parajuga на участках рек, где наиболее часто встречалось сопаразитирование трематод (от 3.3% до 19%), составляла 29-64% при разнообразии паразитов 9-14 видов. В то же время показатели зараженности гастропод Boreoelona, Parafossarulus, Cincinna, Amuropaludina и Cipangopaludina колебались в пределах 0.6-15.9%, а разнообразие трематод, за редким исключением (11 видов), составляло 1-5 видов.
Глава 7. Специфичность в системе трематоды - первые промежуточные хозяева
Проведенные исследования противоречат существующему представлению о моногостальности трематод на этапе развития партеногенетического поколения. Из 28 видов трематод, развивающихся с участием первых промежуточных хозяев моллюсков рода Parajuga, 26 циркулируют с участием различных видов гастропод данного рода. Наиболее часто встречающиеся виды трематод были обнаружены нами у 7-9 видов гастропод рода Parajuga. Кроме того, многие из этих трематод и близкие к ним виды ранее найдены или на стадии партенит у моллюсков других семейств отряда Cerithiiformes, или марит (где отсутствуют моллюски рода Parajuga) на территории Японии, США, Китая, Кореи, Филиппин, Грузии, Украины. То же самое проявляется и у трематод, обнаруженных у моллюсков родов Boreoelona и Parafossarulus (рис. 6).
Работы, в которых рассматриваются проблемы специфичности с позиций экологической сопряженности паразита и хозяина, не дают ответ о причинах моно- или полигостальности на уровне трематоды – первый промежуточный хозяин и, в частности, для видов, имеющих дезъюнктивные ареалы. Для решения этого вопроса требуются комплексные исследования. Они должны обеспечить получение знаний об истории развития взаимоотношений в системе трематоды-хозяева (насколько это возможно), включающей определение центров ее становления, путях расселения ее компонентов и формирования современных фаун, а также о моллекулярно-генетических особенностях животных, составляющих такую систему. Последние могут дать ответы на многие вопросы, касающиеся филогенеза системы трематоды–хозяева, видового статуса морфологически сходных трематод и, соответственно, о их поли- или моногостальности относительно первого промежуточного хозяина.
Таким образом, имеющийся уровень изученности сосальщиков не позволяет пока говорить о причинах поли- и моногостальности трематод в системе трематоды-первые промежуточные хозяева.