
Белорусский государственный университет биологический факультет антитела
| Вид материала | Реферат |
СодержаниеПоказаны гипервариабельные регионы (CDRs). Классификация антител. |
- Белорусский Государственный Университет Биологический факультет Биореакторы Бабицкий, 198.24kb.
- «Белорусский государственный технологический университет», 383.31kb.
- «Белорусский государственный экономический университет», 405.95kb.
- Московский государственный университет имени М. В. Ломоносова биологический факультет, 1266.34kb.
- «Белорусский государственный университет информатики и радиоэлектроники», 644.3kb.
- Управление инновационно-инвестиционными процессами в регионах Беларуси, 43.09kb.
- Домашний телефон, 13kb.
- Осрб 1-36 04 02-2008, 702.53kb.
- Министерство образования республики беларусь белорусский государственный университет, 215kb.
- Белорусский Государственный Экономический Университет, Факультет менеджмента, специальность, 90.11kb.
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Биологический факультет
АНТИТЕЛА.
Реферат
студента 4 курса 6 группы
КОВАЛЬЧУКА К.В.
Минск 2004г.
ОГЛАВЛЕНИЕ
стр.
Открытие антител …………………………………………………………3
Строение антител.………………………………………………………….3
Классификация антител.…………………………………………………...5
Функции антител …………………………………………………………..6
Литература…………………………………………………………………..7
Открытие антител.
Термин «антитело» был введён в употребление в конце XIX века. В 1890 году Беринг (Behring) и Китасато (Kitasato) провели эксперименты, в которых они изучали на морских свинках действие дифтерийного и столбнячного токсинов. Они вводили животным сублетальную дозу токсина, через некоторое время брали у них сыворотку и вводили её вместе с летальной дозой токсина другим животным, в результате чего животные не погибали. Был сделан вывод, что после иммунизации токсином в крови животных появляется вещество, способное нейтрализовать его и тем самым предотвратить заболевание. Данное вещество получило название антитоксина, а затем был введён более общий термин – антитело; вещества вызывающие образование антител стали называть антигенами.
Только в 1939 году Тизелиус (Tiselius) и Кэбет (Kabat) показали, что антитела содержатся в определённой фракции белков сыворотки. Они иммунизировали животное овальбумином и из полученной сыворотки взяли две пробы, в одну из них был добавлен овальбумин и образовавшийся осадок (комплекс антитело-овальбумин) удалили. Электрофорез выявил, что в пробе куда добавлялся овальбумин, содержание γ-глобулинов значительно ниже чем в другой пробе. Это указывало на то, что антитела являются γ-глобулинами. Чтобы отличить их от других белков, содержащихся в этой фракции глобулинов, антитела были названы иммуноглобулинами. Сейчас известно, что антитела обнаруживаются в значительных количествах также во фракциях α- и β-глобулинов.
Структура антител была установлена в ходе разнообразных экспериментов. В основном они заключались в том, что антитела обрабатывались протеолитическими ферментами (папаин, пепсин), и подвергались алкилированию и восстановлению меркаптоэтанолом. Затем исследовались свойства полученных фрагментов: определялась их молекулярная масса (хроматографией), четвертичная структура (рентгеноструктурным анализом), способность связываться с антигеном и т.п. Также использовались антитела к данным фрагментам: выяснялось, могут ли антитела к одному типу фрагментов связываться с фрагментами другого типа. На основе полученных данных была построена описываемая ниже модель молекулы антител.
Строение антител.
Молекула антитела состоит из четырёх полипептидных цепей (рис.1): двух тяжёлых (H; мол.масса 50-70 кДа) и двух лёгких (L; мол.масса 23 кДа). Цепи соединены нековалентными связями (водородные, гидрофобные связи) и дисульфидными мостиками и состоят из двух (лёгкая цепь) или четырёх (тяжёлая цепь) доменов длиной около 110 аминокислотных остатков. Вариабельные домены VH и VL, представляющие собой N-концевые участки цепей, образуют антигенсвязывающий сайт. Помимо них лёгкие цепи содержат по одному (СL), а тяжёлые по три-четыре (СH1-4) константных домена.
При ферментативном расщеплении антител протеолитическим ферментом папаином образуются три фрагмента: два идентичных антигенсвязывающих фрагмента (Fab) и один кристаллизуемый фрагмент (Fc). Fab-фрагмент состоит из интактной L-цепи, связанной дисульфидной связью с доменами СH1 и VH, его N-концевая часть (Fv-фрагмент) обладает антигенсвязывающей активностью. Fc-фрагмент состоит из двух соединённых дисульфидной связью пар доменов CH2 и CH3. Данный фрагмент не участвует в связывании антигенов, а выполняет эффекторные функции – реагирование с клетками и факторами комплемента.
Способность связывания антитела с тем или иным антигеном определяется аминокислотным составом вариабельных доменов, а точнее их гипервариабельных участков. Для этих участков характерна очень высокая изменчивость последовательности аминокислот. Каждый VH и VL домен содержит по три гипервариабельных участка, которые
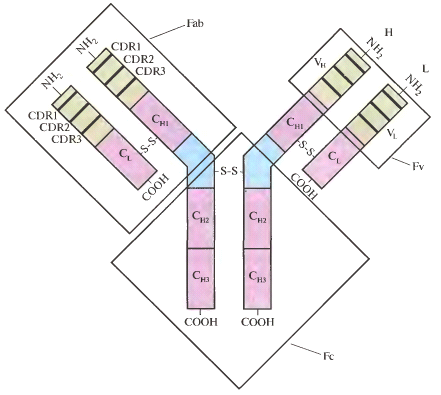
Рис. 1. Строение молекулы антитела. H и L, тяжёлая и лёгкая цепи; CDR, гипервариабельные участки.
собственно и образуют антигенсвязывающие сайты. Последовательности между ними названы каркасными; для них характерна более низкая структурная изменчивость.
Аминокислотная последовательность константной области слабо варьирует. Секвенирование лёгкой цепи выявило существование двух основных вариантов аминокислотных последовательностей СL-доменов, что привело к выделению двух типов лёгких цепей – каппа (κ) и лямбда (λ). Молекула антитела может одновременно содержать либо две κ–цепи, либо две λ–цепи (у антител человека чаще встречаются κ-цепи).
Также определение аминокислотных последовательностей позволило выделить пять типов СH-областей и соответственно - тяжёлых цепей (α, δ, ε, γ, μ). Цепи μ и ε содержат по четыре константных домена, остальные цепи – три константных домена, а также шарнирную область между доменами СH1 и CH2. В зависимости от того, какой тип тяжёлой цепи содержит антитело, различают пять классов иммуноглобулинов: IgA (тяжёлая цепь типа α), IgD (δ), IgE (ε), IgG (γ), IgM (μ). Из-за некоторых различий в аминокислотных последовательностях выделяют несколько типов λ-цепей, а также несколько типов α- и γ-цепей (и соответственно несколько подклассов IgG и IgA). С тяжёлыми цепями (в первую очередь с CH2-доменами) связаны несколько олигосахаридных цепей, которые вероятно увеличивают растворимость антител и участвуют в связывании с компонентами комплемента и клеточными рецепторами.
В доменах полипептидные цепи укладываются формируя β-складчатые слои, в которых антипараллельные цепи соединены петлями (рис.2). Эти петли могут иметь различную длину и аминокислотные последовательности, что очень важно, т.к. именно они формируют антигенсвязывающий сайт. В пределах каждого домена два β-слоя соединены дисульфидной связью и стабилизированы гидрофобными взаимодействиями. Четвертичная структура в форме Y (рис.3) формируется благодаря нековалентным взаимодействиям между доменами. Между доменами CH2 расположены молекулы углеводов, что приводит к выступанию этих доменов и делает их более доступными для взаимодействия с разнообразными молекулами, такими как компоненты системы комплемента.
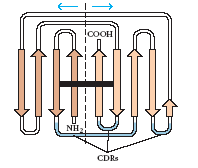
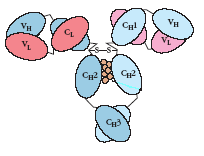
Рис.2. Двумерная схема укладки полипептидной цепи в пределах домена VL: два β-складчатых слоя, соединённых дисульфидной связью (чёрная полоска). Рис.3. Схема, показывающая взаимодействие между доменами лёгкой и тяжёлой цепи. Между доменами CH2 расположены молекулы углеводов.
Показаны гипервариабельные регионы
(CDRs).
Классификация антител.
Как уже было упомянуто выше, в зависимости от типа тяжёлой цепи различают пять классов иммуноглобулинов.
IgG составляют большинство антител сыворотки крови. Большинство антител вторичного иммунного ответа и антитоксинов представлено именно иммуноглобулинами класса G. Материнские IgG обеспечивают пассивный иммунитет ребёнка в первые несколько месяцев жизни, попадая в кровь плода через плаценту. IgG активируют систему комплемента и связываются с поверхностными антигенами клеток, делая тем самым эти клетки более доступными для фагоцитоза (опсонизация). Способны связываться с тканями вызывая анафилаксические реакции.
Молекулы IgM состоят из пяти одинаковых четырёхцепочечных субъединиц, соединённых дисульфидными связями. В их составе также присутствует дополнительная полипептидная цепь (J-цепь), образующая домен иммуноглобулинового типа и связанная дисульфидными связями с С-концевыми пептидами (18 аминокислотных остатков) тяжёлых цепей отдельных мономеров. Предположительно она участвует в полимеризации мономеров. Иммуноглобулины класса М содержатся преимущественно в крови. Доминируют в качестве «ранних» антител (первыми появляются при развитии иммунного ответа). Благодаря множеству участков связывания вызывают агглютинацию клеток. Более эффективно, чем IgG активируют комплемент.
IgA преобладают среди антител серозно-слизистых секретов (слюна, молозиво, молоко, секрет дыхательных путей), где они представлены в основном димерной формой. Как и IgM содержат С-концевой пептид, к которому может присоединятся J-цепь, связывая два мономера в димер. С данным комплексом дополнительно связывается белок, называемый секреторным компонентом, который способствует доставке антител в секреты и защищает их от протеолиза. В сыворотке человека представлены в основном мономерной формой, а в сыворотке других млекопитающих в основном димером. Препятствуют проникновению вирусов, микроорганизмов через слизистые оболочки.
IgD и IgE присутствуют в сыворотке в очень низких концентрациях. IgD часто встречаются на цитоплазматических мембранах В-клеток и предположительно участвуют в антиген-зависимой дифференцировке лимфоцитов. IgE встречаются на мембранах базофилов и тучных клеток. Участвуют в аллергических реакциях, вызывая секрецию клеткой-носителем IgE гистамина и других вазоактивных веществ, в ответ на связывание молекулы IgE с антигеном. Возможно, играют существенную роль в антигельминтозном иммунитете.
Функции антител.
Антитела синтезируются В-лимфоцитами и образующимися из них плазматическими клетками. Их молекулы встроены в цитоплазматическую мембрану В-лимфоцитов, где они функционируют в качестве антигенспецифичных рецепторов. Большинство В-лимфоцитов крови человека экспрессирует на своей поверхности иммуноглобулины двух классов - IgM и IgD. Но в определённых областях тела могут встречаться с высокой частотой В-клетки, несущие антитела других классов (например, IgA в слизистой оболочке кишечника). Плазматические клетки секретируют антитела в плазму крови и тканевую жидкость. Все антитела, синтезируемые одной В-клеткой (или плазматической клеткой), имеют идентичный антигенсвязывающий центр и могут связываться только с одним антигеном.
Первичной функцией антител является связывание с чужеродными (в норме) антигенами с последующей их инактивацией. Антитела способны инактивировать токсины связываясь с зонами молекулы токсина ответственными либо за адсорбцию на клеточных рецепторах либо непосредственно за токсическое действие. Аналогично связывание антител с белками, необходимыми для адсорбции вируса на рецепторы клеток, приводит к инактивации вирионов.
Кроме того, антитела способны вовлекать в иммунный ответ другие элементы иммунной системы: систему комплемента и клетки хозяина. С константными доменами тяжёлой цепи антител классов G и M (с доменами CH2 и CH3 соответственно) способен связываться компонент комплемента C1q. Это вызывает каскад реакций (процесс активации комплемента по классическому пути), в конечном счете приводящих к лизису клетки, с антигенами которой были связаны антитела. Некоторые клетки организма несут на своей поверхности Fc-рецепторы, с которыми посредством Fc-фрагмента могут связываться молекулы антител. Данные рецепторы имеются у макрофагов, что позволяет им распознавать комплексы антиген-антитело с последующим их фагоцитированием (антитела являются опсонинами, т.е. молекулами, которые при связывании с антигенами облегчают их фагоцитирование). Также Fc-фрагмент ответственен за фиксацию антител на клетках определённых тканей и развитие анафилоксических реакций.
К любым антигенам в организме животного изначально существуют антитела. Это предполагает, что каждый организм продуцирует миллионы различных иммуноглобулинов, различающихся своими центрами связывания антигенов. Такое разнообразие обеспечивается несколькими механизмами. Лёгкие и тяжёлые цепи молекул антител кодируются несколькими типами генных сегментов: лёгкая цепь - тремя типами сегментов (V, J, C), тяжёлая - четырьмя (V, D, J, C). В геноме обычно присутствует от нескольких до нескольких сотен сегментов каждого типа, несколько различающихся по нуклеотидной последовательности. Для синтеза цельного полипептида (лёгкой или тяжёлой цепи) необходимо объединение нуклеотидных последовательностей сегментов каждого типа. Такое объединение происходит сначала на уровне ДНК (соматическая рекомбинация), а затем на уровне матричных РНК (сплайсинг). В результате образуется огромное количество вариантов мРНК и соответственно полипептидных цепей. Во время соматической рекомбинации и сплайсинга могут происходить вставки и делеции нуклеотидов, что вместе с повышенной частотой мутаций в генах антител ещё больше повышает разнообразие этих уникальных по своим свойствам белков.
ЛИТЕРАТУРА
1. Иммунология / Ройт А., Бростофф Дж., Мейл Д.-М.:Мир, 2000.-592 с.
2. Иммунология: В 3-х т.; т.1 / Под ред. У. Пола.-М.: Мир, 1987-88.-476 с.