
Эндокринопатии как результат нарушений механизмов образования, транспорта и усвоения гормонов
| Вид материала | Документы |
- Вопросы к итоговому экзамену по эндокринологии в клинической ординатуре, 322.5kb.
- «Молекулярные механизмы гормональной регуляции» Общая трудоемкость изучения дисциплины, 11.18kb.
- Тема: Гормоны, 249.68kb.
- «Средневековый героический эпос» (по одному из произведений, рекомендованных школьной, 79.92kb.
- Основные механизмы нарушений в эндокринной системе, 1046.92kb.
- Примерной программы дисциплины «Теория механизмов и машин» Направление подготовки 160400, 25.47kb.
- Деятельностный подход в обучении, 387.85kb.
- Социальные аспекты психиатрии (вопросы социальной реабилитации, профилактики психических, 236.27kb.
- «Визитная карточка» проекта «Идентификация как результат социализации», 96.28kb.
- «сош №1», 231.26kb.
Эндокринопатии как результат нарушений механизмов
образования, транспорта и усвоения гормонов
1. Общие принципы гормональной регуляции функций организма
Проблемами функционирования эндокринной системы в норме и при патологии гормональной регуляции функций организма занимается наука эндокринология. Можно выделить ряд стандартных (типовых) причин возникновения тех или иных эндокринопатий. Этой проблеме, в сущности, и посвящена настоящая лекция. Однако прежде чем мы рассмотрим происхождение различных эндокринопатий, необходимо разобраться с общими принципами деятельности эндокринной системы.
Эндокринная система, являясь составной частью нейроэндокринной регулирующей системы организма, выполняет функции регуляции и координации деятельности внутренних органов и систем, обеспечивая их адаптацию к постоянно меняющимся условиям внешней и внутренней среды организма, что и является основным условием сохранения гомеостаза. Следует добавить, что функционально нейроэндокринная система тесно связана с иммунной системой организма.
Свое регулирующее действие на ткани, органы и системы организма эндокринная система осуществляет за счет синтеза, секреции, транспорта и воздействия на клетки-мишени специфических биологически активных веществ, получивших название «гормонов» (от греческого «Hormone» - возбуждаю). Термин «гормон» был впервые употреблен Бейлисом и Старлингом в 1905 году при описании секретина – гормонального вещества, открытого ими в желудочно-кишечном тракте.
В общих чертах систему гормональной регуляции можно представить следующим образом (Рис. 1).
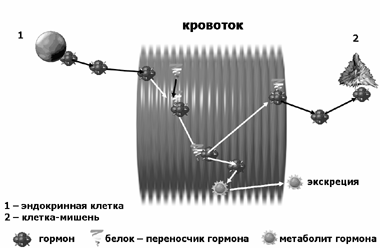
Рис. 1 Схема системы гормональной регуляции
«Классические» гормоны (об отличии «классических» гормонов от некоторых других гормонально подобных веществ будет сказано позднее) синтезируются и депонируются в клетках соответствующих эндокринных желез, как правило, в виде прогормонов. Далее, в соответствии с метаболическими потребностями организма, они поступают в кровоток и переносятся кровью обычно в виде комплексов с белками плазмы. Достигая клеток-мишеней, имеющих мембранные или внутриклеточные рецепторы, комплементарные молекулам соответствующего гормона, гормоны отсоединяются от своего белкового носителя, соединяются с рецептором и запускают каскад внутриклеточных биохимических процессов, в результате которого клетка изменяет свой метаболизм. Излишки гормонов разрушаются соответствующими ферментами (преимущественно в печени) и выводятся из организма через почки.
Механизмы секреции гормонов, их транспорта, использования гормонов клетками-мишенями, деградации гормонов и выведение продуктов их распада из организма образуют взаимоуравновешенную систему, которая обеспечивает поддержание так называемого «базального уровня гормонов в крови». Соответствующая схема приведена на Рис. 2.
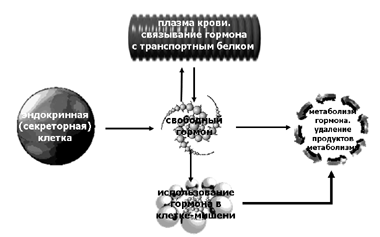
Рис. 2 Механизмы поддержания базального уровня гормонов в крови
Помимо уже указанного выше «классического» действия гормона на клетки-мишени, когда транспорт гормона осуществляется кровотоком на относительно большие расстояния, существуют и другие принципы действия гормонов на регулируемые ими клеточные популяции. При паракринном действии гормона он после секреции эндокринными клетками осуществляет свое влияние на рядом расположенные клеточные популяции. Так, например, действуют некоторые гормоны желудочно-кишечного тракта. Возможна и ситуация, когда гормон после его секреции оказывает воздействие на саму секретирующую клетку. Это так называемый аутокринный механизм действия гормонов. Примером может служить действие простагландинов (биологически активных веществ, обладающих гормоноподобным действием) на клетку, в которой эти вещества и были образованы. Можно выделить и еще один тип действия гормонов на клетку-мишень. Это – нейроэндокринное действие медиаторов и нейротрансмиттеров, секретируемых нервными окончаниями и передающими или изменяющими нервные регуляторные влияния на иннервируемую ткань. Схема, иллюстрирующая принципы действия гормонов, приведена на Рис.3.
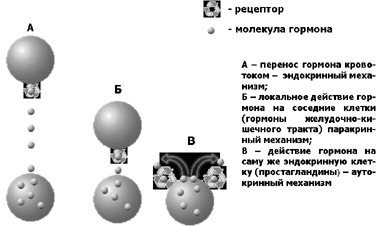
Рис. 3 Принципы действия гормонов
Регуляция функций организма по принципу обратной (положительной и отрицательной) связи широко известен физиологам и патологам. Не является исключением в этом правиле и механизмы регуляции деятельности желез внутренней секреции.
Наиболее простым примером регуляции функции по принципу отрицательной обратной связи является взаимозависимость уровня продукции гормона (например, инсулина) от концентрации вещества (например, глюкозы), степень усвоение которого клетками определяется данным гормоном. В нашем примере повышение уровня глюкозы в крови вызывает повышенную продукцию инсулина, а снижение его – уменьшение продукции гормона (Рис. 4).
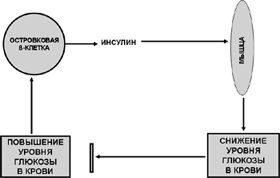
Рис. 4 Регулирование функции эндокринной железы по принципу
отрицательной обратной связи (Д.Ф.Лейкок, П.Г.Вайс)
Регуляция функций эндокринной железы по принципу обратной связи может иметь и более сложный, иерархически построенный механизм. В таких случаях от уровня гормона в крови зависит, с одной стороны, выработка релизинг-факторов гипоталамусом, а с другой, выработка соответствующего тропного гормона. При этом, отрицательные обратные связи, замыкаясь на разных уровнях, образуют короткие и длинные петли. Пример такой усложненной регуляции представлен на
рис. 5.
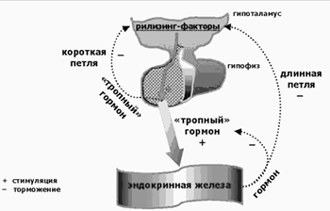
Рис. 5 Регуляция секреции гормона по принципу отрицательной обратной
связи (короткие и длинные регуляционные петли)
В редких случаях регуляция секреции гормона может осуществляться и по принципу положительной обратной связи. Иначе говоря, секреция определенного гормона может служить стимулирующим фактором для увеличения его выработки той же самой железой. Так, например, секреция эстрадиола развивающимся фолликулом яичника способна приводить к повышенной чувствительности гормонопродуцирующих клеток этого образования к действию гонадотропинов, вырабатываемых аденогипофизом. В результате синтез и секреция эстрадиола возрастают. Механизм этого процесса, скорее всего, объясняется тем обстоятельством, что под влиянием эстрадиола происходит увеличение числа рецепторов гонадотропина на мембране гормонопродуцирующих клеток фолликула.
Положительные обратные связи обычно существуют короткое время и быстро разрушаются, так как значительное повышение уровня соответствующего гормона
в крови быстро включает гораздо более действенные отрицательные обратные связи.
2. Классификации гормонов
Прежде чем перейти к обсуждению материала, посвященного проблемам синтеза, секреции и взаимодействия гормонов с клетками-мишенеми, необходимо разобраться с классификацией (а, точнее, с классификациями) гормонов.
По особенностям растворимости и взаимодействия со внутренней средой организма гормоны можно разделить на две группы: липофильные и гидрофильные гормоны.
Липофильные гормоны, к которым относятся все стероидные гормоны и тироидные гормоны (тироксин и трийодтиронин) обладают следующими особенностями:
- нерастворимостью в плазме крови;
- относительно небольшим молекулярным весом;
- способностью свободно проходить через мембрану клеток-мишеней и взаимодействовать с внутриклеточными (плазматическими) рецепторами;
- сразу после синтеза покидать соответствующие гормонопродуцирующие клетки, не депонироваться в них;
- транспортироваться от гормонопродуцирующей клетки к клетке-мишени в соединении с транспортным плазматическим белком.
Гидрофильные гормоны, к которым относятся гормоны белковой, полипептидной природы и гормоны, являющиеся производными аминокислот, обладают следующими особенностями:
- растворимостью в плазме крови;
- большим молекулярным весом;
- способностью депонироваться в гормонопродуцирующих клетках и покидать их по мере необходимости в их использовании;
- синтезироваться в виде прогормонов и конверсироваться в менее высокомолекулярное вещество в клеточном аппарате Гольджи;
- взаимодействовать с клетками-мишенями, соединяясь с плазматическими клеточными рецепторами.
По своему химическому составу гормоны делятся на несколько групп.
Большую группу гормонов составляют белковые и полипептидные гормоны. В эту группу входят адренокортикотропный гормон, соматотропный гормон, меланостимулирующий гормон, пролактин, паратгормон, кальцитонин, инсулин, глюкагон, тиреотропный гормон, фолликулостимулирующий гормон, тироглобулин. Гипофизотропные гормоны (релизинг-факторы) и гормоны желудочно-кишечного тракта относятся к олигопептидам (малым пептидам).
Вторая группа гормонов – это липидные или стероидные гормоны. К ним относятся кортикостерон, кортизол, альдостерон, прогестерон, эстрадиол, эстриол, тестостерон.
Третья группа включает гормоны – производные аминокислот. Это адреналин, норадреналин, тироидные гормоны.
Следует указать, что многие исследователи и клиницисты относят к гормонам и такие сигнальные вещества как медиаторы, нейромедиаторы, ростовые факторы и цитокины. Однако в нашей лекции мы будем обсуждать проблемы эндокринопатий, связанных с нарушениями синтеза, секреции, транспорта и усвоения клетками-мишенями «классических» гормонов, о которых шла речь при описании их классификаций.
3. Механизмы биосинтеза и секреции гормонов
3.1. Механизм биосинтеза и секреции белковых и полипептидных гормонов
Биосинтез белковых и полипептидных гормонов осуществляется под контролем генетического аппарата гормонопродуцирующей клетки. Запуск процесса биосинтеза начинается после воздействия на рецепторный аппарат гормонопродуцирующей клетки соответствующей сигнальной молекулы, которая в большинстве случаев является молекулой тропного гормона. С рецептора сигнал передается на внутриклеточный эффекторный белок, чаще всего обладающий ферментативной активностью. Благодаря своей ферментативной активности внутриклеточный сигнальный белок приобретает возможность проникнуть в ядро клетки и там фосфорилировать и активировать белки, участвующие в транскрипции генов. После переноса информации о строении синтезируемого белка с ДНК на молекулу мРНК последняя связывается рибосомами, располагающимися в шероховатом эндоплазматическом ретикулуме (ШЭР), а тРНК в это время транспортирует на рибосомы молекулы соответствующих аминокислот, из которых и собирается синтезируемый белок. В результате образуется высокомолекулярное соединение, обозначаемое как препрогормон. Пептидный участок препрогормона, обозначаемый как «пре» (или сигнальный пептид) служит для прикрепления молекулы препрогормона к эндоплазматическому ретикулуму. После отщепления от молекулы препрогормона сигнального пептида происходит образование прогормона, который, путем ограниченного протеолиза, превращается в молекулу белкового гормона и в аппарате Гольджи «упаковывается» в секреторные везикулы, защищающие молекулы гормона от воздействия белков цитоплазмы. В этих везикулах гормоны могут длительное время депонироваться в гормонопродуцирующих клетках и по мере необходимости путем экзоцитоза покидать клетку и поступать в кровоток. Транспорт везикул к мембране клетки и последующий экзоцитоз осуществляется с участием ионов Са++ и требует затраты энергии. Повышение концентрации внутриклеточного Са++ осуществляется за счет его поступления в клетку из митохондрий, эндоплазматического ретикулума, а также из внеклеточной среды. Кроме того, в транспорте везикул и экзоцитозе принимают участие и такие внутриклеточные образования как микротрубочки и микрофиламенты. Схема, иллюстрирующая механизмы биосинтеза и секреции белковых и полипептидных гормонов, представлена на Рис. 6.
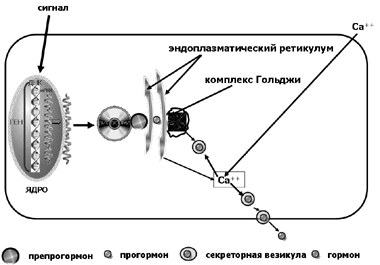
Рис. 6 Механизм биосинтеза и секреции белковых и полипептидных
гормонов
- Механизм биосинтеза и секреции стероидных гормонов
Общим предшественником всех стероидных гормонов является холестерин.
Первый этап синтеза стероидных гормонов – превращение холестерина в прегненалон является общим для всех клеток, участвующих в стероидогенезе. Более того, именно эта реакция и регулирует скорость общего стероидогенеза в организме. Далее, путем сложных ферментативных реакций из прегненалона образуются конечные продукты синтеза стероидов: минералкортикоиды, глюкокортикоиды, андрогены и эстрогены. Обобщающая схема этого процесса представлена на Рис.7.
Избыток холестерина накапливается в виде эфиров жирных кислот, которые образуют так называемые «липидные капли» в протоплазме клеток. Скорость и объемы синтеза стероидных гормонов в конечном итоге зависят только от количества холестерина поступающего в клетки или образующегося в них.
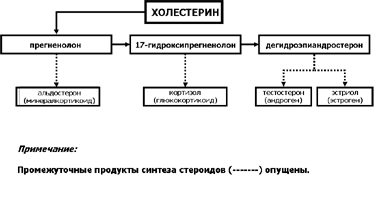
Рис. 7. Основные этапы синтеза стероидных гормонов
Существуют два пути поступления (образования) холестерина в гормонопродуцирующие клетки эндокринных желез. Во-первых, холестерин может синтезироваться в этих клетках из АцКоА. Во-вторых, поступать в клетки в составе липопротеинов низкой плотности (ЛПНП). В этом случае ЛПНП соединяются с соответствующими рецепторами гормонопродуцирующей клетки, которые транспортируют их в цитоплазму. Там под влиянием ферментов лизосом от комплекса рецептор – ЛПНП отщепляется рецепторы, которые могут снова встраиваться в мембрану клеток. Кроме того, под влиянием лизосомных ферментов из ЛПНП выделяется холестерин, на основе которого в митохондриях синтезируются стероидные гормоны.
Так как стероидные гормоны относятся к группе липофильных гормонов, они свободно проходят через мембрану гормонопродуцирующих клеток и поступают в кровоток, где соединяются с транспортными белками плазмы. По мере потребности стероидные гормоны отделяются от транспортных белков и поступают в клетки-мишени. Определенная часть гормонов, оставаясь в соединении с транспортными белками, депонируется в плазме крови. Инактивация стероидных гормонов происходит в печени, а выведение продуктов их распада осуществляется через почки и, частично, с желчью.
Схема, иллюстрирующая синтез стероидных гормонов на основе холестерина, полученного из ЛПНП, приведена на Рис. 8.
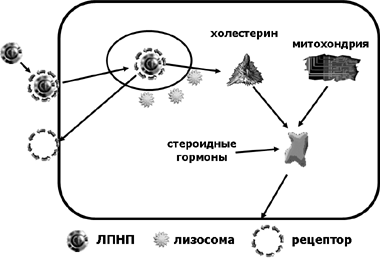
Рис. 8. Участие липопротеинов низкой плотности (ЛПНП) в синтезе
стероидных гормонов
- Механизмы действия гормонов на клетку-мишень
4.1. Механизм действия липофильных гормонов на клетку-мишень
Как известно, липофильные гормоны транспортируются от гормонопродуцирующих клеток к клеткам-мишеням в соединении с белками плазмы крови. Перед началом воздействия липофильного гормона на клетку-мишень от него отщепляется транспортный белок, а молекула гормона, благодаря своей липофильности, свободно проходит через клеточную мембрану и, в протоплазме клетки, соединяется с соответствующим внутриклеточным рецептором. По своему строению внутриклеточные рецепторы являются белками, находятся в клетке в количестве 103 - 104 молекул на клетку и обладают высокой избирательностью по отношению к конкретным гормонам.
Рецепторные белки состоят из четырех доменов (гормоносвязывающий домен, сайт-специфичный домен, регуляторный и ДНК-связывающий домены). После соединения гормона с рецепторным белком, этот комплекс проникает внутрь ядра клетки и своим ДНК-связывающим доменом соединяется с так называемом гормон-рецессивным участком ДНК клетки. При этом каждый комплекс гормон – рецептор обеспечивает транскрипцию лишь одного участка ДНК, ответственного за деятельность определенного гена, кодирующего синтез определенного белка. Сам синтез белка идет по классической схеме белкового биосинтеза. Появление нового белка соответствующим образом меняет метаболизм клетки-мишени. Схема, иллюстрирующая механизм действия липофильных гормонов на клетки-мишени, приведена на Рис. 9.
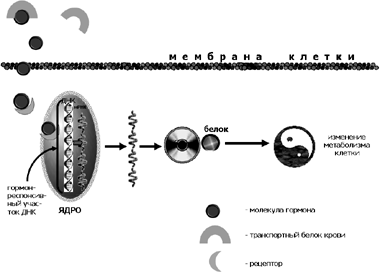
Рис. 9. Механизм действия липофильных гормонов на клетки-мишени
- Механизмы действия гидрофильных гормонов на клетки-мишени
Гидрофильные гормоны не способны проникать через мембрану клетки и поэтому передача сигнала осуществляется при посредстве мембранных белков-рецепторов.
Существует три типа этих рецепторов.
Первый тип – это белки, имеющие одну трансмембранную полипептидную цепь. С рецепторами этого типа соединяются такие гормоны как соматотропный гормон, пролактин, инсулин и ряд гормоноподобных веществ – ростовых факторов. При соединении гормона с рецептором этого типа происходит фосфорилирование цитоплазматической части этого рецептора, в результате чего осуществляется активация белков-посредников (мессенджеров), обладающих ферментативной активностью. Это свойство позволяет белку-мессенджеру проникнуть в ядро клетки и там активировать ядерные белки, участвующие в транскрипции соответствующих генов и мРНК. В конечном итоге, клетка начинает синтезировать специфические белки, которые и меняют ее метаболизм. Cхема, иллюстрирующая этот механизм, представлена на Рис. 10.
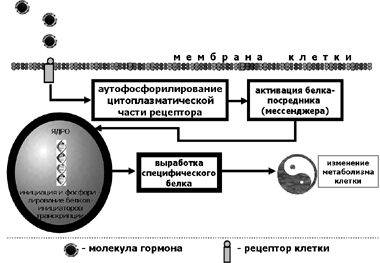
Рис. 10. Механизм действия гидрофильных гормонов на клетку-мишень,
имеющую рецепторы первого типа
Второй тип рецепторов, воспринимающих воздействие гидрофильных гормонов на клетки-мишени – это так называемые «рецепторы – ионные каналы». Рецепторы этого типа являются белками, имеющими четыре трансмембранных фрагмента. Соединение молекулы гормона с таким рецептором приводит к открытию трансмембранных ионных каналов, благодаря чему ионы электролитов по градиенту концентрации могут поступать в протоплазму клетки. С одной стороны это может приводить к деполяризации клеточной мембраны (так, например, происходит с постсинаптической мембраной клеток скелетной мышцы при передаче сигнала с нервного моторного волокна на мышцу), а с другой – ионы электролитов (например, Са++) могут активировать серин-тирозиновые киназы, и за счет их ферментативного действия на внутриклеточные белки вызывать изменение клеточного метаболизма.
Схема, иллюстрирующая этот механизм, представлена на Рис. 11.
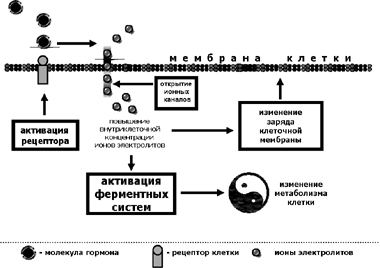
Рис. 11. Механизм действия гидрофильных гормонов на клетку-мишень,
имеющую рецепторы второго типа
Третий тип рецепторов, воспринимающих воздействие гидрофильных гормонов на клетки-мишени, определяется как «рецепторы, сопряженные с G-белками» (сокращенно – GPCR – “G-protein coupled receptors”).
С помощью G-рецепторов на исполнительный клеточный аппарат передаются сигналы, возбуждаемые нейропередатчиками и нейротрансмиттерами (адреналин, норадреналин, ацетилхолин, серотонин, гистамин и др.), гормонами и опиоидами (адренокортикотропин, соматостатин, вазопрессин, ангиотензин, гонадотропин, некоторые факторы роста и нейропептиды и др.). Кроме того, G-рецепторы обеспечивают передачу химических сигналов, воспринимаемых вкусовыми и обонятельными рецепторами.
G-рецепторы, равно как и большинство мембранных рецепторов, состоят из трех частей: внеклеточная часть, часть рецептора, погруженная в мембрану клетки и внутриклеточная часть, контактирующая с G-белком. При этом внутримембранная часть рецептора – это полипептидная цепочка, пересекающая мембрану семь раз.
Функцией G-белков является открытие ионных каналов (т.е. изменение концентрации ионов электролитов в протоплазме клеток-мишеней) и активация белков-посредников (внутриклеточных мессенджеров). В результате с одной стороны происходит активация соответствующих ферментных систем клетки (протеинкиназ, протеинфосфатаз, фосфолипаз), а с другой, фосфорилированные белки, обладающие мощной ферментативной активностью, приобретают возможность проникнуть в ядро клетки и там фосфорилировать и активировать белки, участвующие в транскрипции генов и мРНК. В конечном итоге, метаболизм клетки меняется как за счет ферментативных превращений внутриклеточных белков, так и благодаря биосинтезу новых белков. Схема, иллюстрирующая механизмы взаимодействия молекулы гормона с G-рецептором клетки-мишени, приведена на Рис. 12.
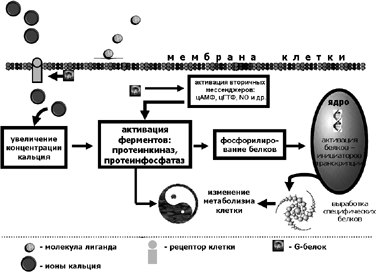
Рис. 12 Механизм действия гидрофильных гормонов на клетку-мишень,
обладающую G-рецептором
- Эндокринопатии как результат нарушения гормональной регуляции функций организма
5.1. Основные причины нарушений гормональной регуляции
Гормональная регуляция функций организма – это сложная, хорошо сбалансированная система, обладающая многократно сдублированными управляющими связями. Несмотря на это, в системе гормональной регуляции возможны сбои и «неисправности», которые возникают на самых разных участках пути, начинающегося в гормонопродуцирующей клетке и заканчивающегося в клетке-мишени. В результате могут возникать самые разнообразные эндокринопатии, диагностике и лечению которых собственно и посвящена такая наука как эндокринология.
В нашу задачу не входит подробный разбор и описание каждого из нарушений гормональной регуляции, каждой эндокринопатии. Мы остановимся только на типовых нарушениях процесса гормональной регуляции функций организма, которые могут возникать в самых различных звеньях регуляционной цепочки, начиная от биосинтеза и секреции соответствующего гормона, и кончая его воздействием на метаболизм клеток-мишеней.
Опишем коротко те типовые нарушения, которые могут возникать в этих звеньях.
На уровне эндокринной железы мы можем встретиться с такими явлениями как нарушение восприятия сигнала, инициирующего биосинтез гормона (то есть, нарушение рецепции сигнала), нарушение пострецепторной передачи сигнала, нарушение синтеза и секреции соответствующего гормона. Следует иметь в виду, что большинство эндокринных желез находятся под двойным регуляционным контролем: со стороны аденогипофиза, вырабатывающего тропные гормоны, выработка которых, в свою очередь, контролируется гипоталамусом, вырабатывающим соответствующие релизинг-факторы.
«Классические» гормоны после этапа секреции поступают в общий кровоток. Многие из них (например, все липофильные гормоны, то есть стероиды и иодтиронины) переносятся кровотоком в соединении с транспортными белками плазмы. Другие гормоны (гидрофильные) переносятся током крови без посредников – транспортных белков. Однако, и в том, и в другом случае на этапе транспортировки гормонов могут происходить некоторые нарушения, которые, в конечном итоге, скажутся на эффективности гормональной регуляции функций организма.
На этапе воздействия гормона на клетку-мишень так же могут возникать различные нарушения, приводящие к развитию тех или иных эндокринопатий. Это может быть нарушение рецепции гормона клеткой-мишенью, нарушения в деятельности различных первичных или вторичных мессенджеров (пострецепторных исполнителей и передатчиков полученного гормонального сигнала). Наконец, нарушения могут коснуться этапов транскрипции соответствующих генов и, как следствие, привести к нарушению синтеза соответствующих белков, участвующих в изменении метаболизма клетки-мишени.
5.1.1. Гипо- и гиперфункция эндокринных желез
Гипо – и гиперфункция эндокринных желез – это наиболее часто встречающаяся эндокринная патология.
В большинстве случаев гипофункцию эндокринных желез можно по ее происхождению разделить на первичную (нарушение секреторной функции самой железы), вторичную (гипофизарную, связанную с недостатком выработки соответствующего тропного гормона) и третичную (гипоталамическую, связанную с недостатком выработки релизинг-фактора, стимулирующего деятельность аденогифифиза, или с избытком гипоталамического ингибирующего фактора, тормозящего выработку соответствующего тропного гормона).
Примером хорошо известной гипофункции эндокринной железы является гипотиреоз.
Первичный (тиреогенный) гипотиреоз может возникнуть в результате развития аутоиммунного тиреоидита, аномалий развития щитовидной железы, врожденных энзимопатий с нарушением биосинтеза тиреоидных гормонов.
Вторичный (гипофизарный) гипотиреоз может быть связан с гипофизарной недостаточностью. Например, при гормононепродуктивной опухолью гипофиза.
Третичный (гипоталамический) гипотиреоз обусловлен нарушениями синтеза и секреции клетками гипоталамуса релизинг-фактора тиреолиберина.
Как и в случаях с гипофункцией эндокринных желез, их гиперфункция по происхождению может быть разделена на первичную (связанную с гиперсекреторными процессами, происходящими в самой железе), вторичную (гиперпродукция гормона в результате увеличения синтеза тропного гормона) и третичную (гипоталамическую, обусловленную избытком выработки соответствующего релизинг-фактора).
Примером хорошо известной гиперфункции эндокринной железы является гиперкортицизм.
Первичный гиперкортицизм – синдром Иценко-Кушинга возникает чаще всего в результате гормонально активной опухоли надпочечника.
Вторичный гиперкортицизм – болезнь Иценко-Кушинга связан с гиперпродукцией адренокортикотропного гормона аденогипофизом. Чаще всего такая ситуация возникает при развитии гормонально активной опухоли гипофиза – базофильной аденоме.
Третичный гиперкортицизм – результат повышенной выработки релизинг-фактора кортиколиберина клетками гипоталамуса.
5.1.2. Нарушения транспорта гормонов
Многие гормоны, циркулирующие в крови, находятся в связанном состоянии с белками плазмы. Связанные с белками гормоны представляют собой своеобразное «депо» гормонов, откуда по мере потребности гормоны высвобождаются и оказывают свое действие на клетки-мишени.
Концентрация свободных липофильных гормонов в крови (а только свободные гормоны могут проникать через мембрану клеток-мишеней) определяется прочностью их связей с транспортными белками, а также зависит от достаточности содержания белков в плазме крови. Нехватка белков плазмы, например, при заболеваниях печени или при повышенном выведении белков плазмы из организма при хронических нефрозах, приводит к избытку свободных гормонов, что, в свою очередь, может вызвать их повышенный распад под влиянием ферментных систем крови и вывод продуктов метаболизма гормонов из организма. С другой стороны, есть белки плазмы, обладающие повышенным сродством к некоторым гормонам, в результате чего могут образовываться очень прочные соединения молекул гормонов с транспортным белком. При такой ситуации в крови снижается количество свободных гормонов и может развиться своеобразная «транспортная» гипофункция эндокринных желез. К белкам плазмы, обладающим такими свойствами, относятся, например, тироксинсвязывающий глобулин и тироксинсвязывающий преальбумин, а также кортикостероидсвязывающий глобулин.
5.1.3. Нарушения рецепции гормонов клетками-мишенеми
Нарушения рецепции гормонов могут происходить либо в результате недостатка синтеза рецепторов клеткой мишеней, либо снижения или отсутствия сродства рецепторов к соответствующему гормону благодаря их аномальному синтезу. Последнее обстоятельство чаще всего бывает обусловлено наследственными заболеваниями (например, синдром резистентности к адренокортикотропному гормону благодаря мутации гена рецептора АКТГ).
Воздействие гормона на клетку-мишень может быть нарушено и в результате патологии пострецепторных механизмов, например, на уровне G-белков вследствие мутации, а также при повреждении и других клеточных мессенджеров.
Изменение деятельности гормонопродуцирующих клеток может происходить в результате своеобразной «мимикрии» некоторых высокомолекулярных веществ под молекулы соответствующих гормонов. В этом случае клетка, получив подобный сигнал, начинает интенсивно вырабатывать соответствующий гормон, в котором в данный момент организму не нужен. Так происходит, например, когда в организме беременной женщины в силу тех или иных обстоятельств начинают образовываться антитела класса IgG, высокоаффинные к рецепторам гормонопродуцирующих клеток щитовидной железы плода и имитирующие действие на них тиреотропного гормона. В результате у плода появляются признаки тиреотоксикоза. Такой же мимикрией обладают антитела, которые при некоторых обстоятельствах могут вырабатываться к гормону инсулину. Соединяясь с инсулиновыми рецепторами клеток различных тканей организма, антитела блокируют их и не дают нормальным молекулам инсулина соединиться с инсулиновыми рецепторами. В конечном итоге у такого человека может развиться диабет.
5.1.4. Эндокринопатии, обусловленные нарушением регуляции по принципу «обратной связи»
Регуляция физиологических функций по принципу обратной связи (в первую очередь – по принципу отрицательной обратной связи) хорошо известна. В эндокринной системе по принципу обратной отрицательной связи регулируется выработка релизинг-факторов, тропных гормонов и, в конечном итоге, выработка гормонов соответствующими железами внутренней секреции. Однако применение заместительной гормональной терапии, а также лечение некоторых заболеваний гормональными препаратами может вносить определенный разлад в стройную систему эндокринной регуляции. Речь идет о так называемом «синдроме отмены», то есть длительной гипофункции эндокринной железы, связанной с угнетением выработки определенного тропного гормона гипофизом. Регулярное введение в организм достаточно больших доз гормонов (например, при лечении хронических воспалительных процессов кортикостероидами) по принципу отрицательной обратной связи тормозит выработку тропного гормона гипофизом и при резком прекращении приема препаратов, содержащих определенный гормон, эндокринная железа, не получающая должной стимуляции от гипофиза, впадает в состояние длительной гипофункции. Чтобы избежать подобной ситуации отмена гормоносодержащих препаратов должна производить постепенно. В этом случае так же постепенно будет нарастать выработка тропного гормона и гипофункция эндокринной железы не разовьется.
5.1.5. Эндокринопатии, обусловленные образованием аномальных гормонов
Аномальные гормоны могут продуцироваться эндокринным железами в результате мутаций генов, ответственных за их синтез. Известен, например, аномальный инсулин, неспособный соединяться с инсулиновыми рецепторами клеток различных тканей организма. В этом случае клетки лишаются возможности получать глюкозу, и у такого пациента развивается диабет.
В известной степени к аномальным гормонам можно отнести и аутоантитела, соединение которых рецепторами клеток эндокринных желез или клеток-мишеней приводит к гиперпродукции соответствующего гормона или не позволяет нормальным гормонам воздействовать на рецепторный аппарат клеток-мишеней. Впрочем, эта ситуация уже обсуждалась в разделе, посвященном так называемой «мимикрии гормонов».
Таковы основные типовые причины, лежащие в основе эндокринопатий.