
Структурно-функциональные свойства эритроцитарных мембран при дислипопротеинемиях 03. 00. 04 биохимия 14. 00. 16 патологическая физиология
| Вид материала | Автореферат |
- Роль молекул оксида азота в программированной гибели нейтрофилов при окислительном, 343.37kb.
- Пояснительная записка Учебно-методический комплекс по курсу «Патологическая физиология», 140.7kb.
- Гормональный статус, генетический полиморфизм и мотивационно-потребностные особенности, 450.55kb.
- Структурно-семантические и функциональные свойства идиоматичных слов английского языка, 390.28kb.
- Программа вступительных испытаний для специальности магистратуры 1-79 80 29 Патологическая, 348.44kb.
- Пояснительная записка к учебной дисциплине Эпизоотология и инфекционные болезни животных, 166.41kb.
- Николаева Мария Андреевна, доктор технических наук, профессор, заведующий кафедрой, 367.89kb.
- Вопросы к экзамену по дисциплине «спортивная физиология», 205.39kb.
- Патофизиологический анализ влияния факторов риска образовательной среды на функциональное, 816.71kb.
- Исследование механических характеристик нелинейно деформируемых сферических мембран, 43.3kb.
1 2
Рис. 1. Содержание подфракции 2 липопротеинов высокой плотности (ЛПВП2), холестерина липопротеинов высокой плотности (ХС ЛПВП), суммарных апобелков липопротеинов высокой плотности (апо ЛПВП) и общего холестерина у больных сахарным диабетом типа 1 в зависимости от длительности сахарного диабета (а), степени компенсации углеводного обмена (б) и степени сосудитых осложнений (в).
Примечания. Здесь, и в рис. 2-3: * p<0,05 по сравнению со значениями здоровых доноров; ** p<0,01 по сравнению со значениями здоровых доноров; *** p<0,001 по сравнению со значениями здоровых доноров.
Анализируя данные изменения основных показателей липидного спектра сыворотки крови у больных сахарным диабетом в соответствии с классификацией D. Fredrikson, нами было отмечено частое развитие ДЛП IIб типа (у 29,4 % больных СД 1 и у 40,6 % больных СД 2) и IIа типа (22,2 % больных СД 1 и 40,6 % больных СД 2); IV тип ДЛП (12,5% больных СД 2) встречался реже. Таким образом, были выделены категории пациентов СД с различными нарушениями липидного обмена с учетом типа и степени их выраженности.
К настоящему времени сформировалось мнение, что при сахарном диабете патология липидного обмена создает основу для дезорганизации структуры и функции мембран эритроцитов [Кондратьева Е.И., 2001; Broncel M. et al., 2005] В этой связи особый интерес представляет выявление общих закономерностей и особенностей модификации мембран эритроцитов у больных сахарным диабетом с различными нарушениями липидного метаболизма.
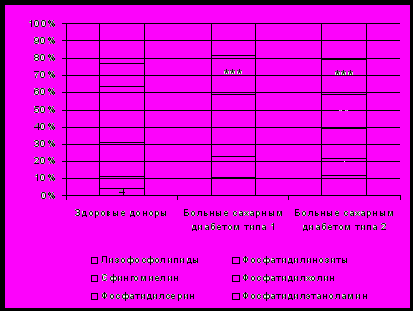
Рис. 2. Содержание фракций фосфолипидов в мембране эритроцитов у больных сахарным диабетом типа 1 и 2
Как показали хроматографические исследования, изменения относительного содержания фракций мембранных липидов эритроцитов у всех пациентов с сахарным диабетом были практически однонаправленными и характеризовались увеличением доли ХС и его эфиров, снижением содержания ФЛ. При этом среди ФЛ преобладали легкоокисляемые формы (фосфатидилсерин, фосфатидилэтаноламин, фосфатидилинозитолы) на фоне значительного уменьшения доли ФХ, составляющего основу липидной фазы мембран эритроцитов у здоровых доноров, повышалось содержание лизофосфолипидов (рис. 2.).
 Известно, что стабильное соотношение между фракциями фосфолипидов является необходимым условием оптимального функционирования мембраны эритроцита [Черняк Н.Б., 1976; Крепс Е.М., 1982]. Увеличение содержания ФС и ФИ на фоне уменьшения доли ФХ приводит к значительной дезорганизации липидного бислоя, проявляющейся прежде всего нарушением асимметрии мембраны. Выявленное у пациентов с СД повышение содержания лизофосфолипидов может быть связано как с повышением гидролиза ФЛ, так и со снижением активности лизо-фосфолипазы. В литературе есть данные, позволяющие считать, что ЛПВП принимают участие в «регенерации» бислоя, удаляя перекисномодифицированные остатки ФЛ, тем самым участвуя в восстановлении липидной фазы [Климов А.Н. и соавт., 2001]. Таким образом, недостаточность ЛПВП способствует усугублению дегенеративных процессов, протекающих в мембране. У пациентов с СД 2 наибольшее повышение содержания лизофосфолипидов в мембранах красных клеток крови было отмечено у больных с сопутствующим ожирением и оказалось равным 12,5 (6,0-20,0) %.
Известно, что стабильное соотношение между фракциями фосфолипидов является необходимым условием оптимального функционирования мембраны эритроцита [Черняк Н.Б., 1976; Крепс Е.М., 1982]. Увеличение содержания ФС и ФИ на фоне уменьшения доли ФХ приводит к значительной дезорганизации липидного бислоя, проявляющейся прежде всего нарушением асимметрии мембраны. Выявленное у пациентов с СД повышение содержания лизофосфолипидов может быть связано как с повышением гидролиза ФЛ, так и со снижением активности лизо-фосфолипазы. В литературе есть данные, позволяющие считать, что ЛПВП принимают участие в «регенерации» бислоя, удаляя перекисномодифицированные остатки ФЛ, тем самым участвуя в восстановлении липидной фазы [Климов А.Н. и соавт., 2001]. Таким образом, недостаточность ЛПВП способствует усугублению дегенеративных процессов, протекающих в мембране. У пациентов с СД 2 наибольшее повышение содержания лизофосфолипидов в мембранах красных клеток крови было отмечено у больных с сопутствующим ожирением и оказалось равным 12,5 (6,0-20,0) %. Полагают, что деградация фосфолипидного остова является одной из причин повышения содержания ХС в мембране эритроцитов при патологии. В результате повышенной активности фосфолиполиза и свободно-радикального окисления происходит разрушение фосфолипидных молекул, вследствие чего уменьшается их содержание в мембране, замещаясь на молекулы ХС [Болдырев А.А., 1990, 2006] . При дислипопротеинемиях наибольшее значение приобретает нарушение взаимодействия липопротеинов и плазмалеммы. В кровотоке постоянно происходит обмен липидными компонентами между мембраной эритроцитов и плазменными липопротеинами, причем ХС переходит в мембрану от ЛПОНП и ЛПНП, откуда возможно его включение в состав ЛПВП. Тем самым формируется равновесная система липопротеинов и плазмалеммы красных кровяных клеток, участвующая в регуляции структурного состояния последней [Bruckdorfer K.R., 1967; Quarford S.H., Hilderman H.L. 1970; Hollan S. et al., 1995, 1996; Broncel M. et al., 2005]. Изменение соотношения липопротеиновых комплексов приводит к нарушению равновесия миграции ХС, при этом значительное превалирование количества липопротеинов атерогенных классов над содержанием ЛПВП создают условия для преимущественного поступления ХС в мембраны эритроцитов.
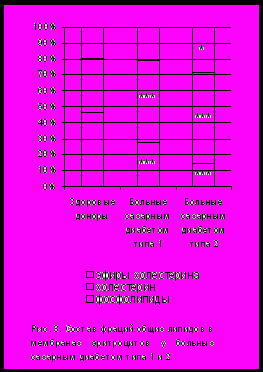
По данным нашего исследования у пациентов с сахарным диабетом повышение содержания ХС в мембранах эритроцитов было достоверно значимым; причем степень увеличения доли ХС в мембранах не различалась у больных СД 1 и СД 2 (рис. 3.). Следует отметить, что у этих пациентов выявлялось увеличение значений индекса атерогенности и отношения ХС/ФЛ в сыворотке крови. Тем не менее обращало на себя внимание, что значительное повышение доли ХС в мембранах эритроцитов у пациентов с СД 1 не коррелировало с какими-либо особенностями клинического течения заболевания. Однако у больных СД 2 было отмечено увеличение доли эстерифицированного ХС по сравнению с его содержанием у пациентов СД 1 и здоровых доноров. Как известно, эстерификация ХС – один из способов его модификации, способствующий последующей акцепции ЛПВП [Болдырев А.А., 1990]. Поэтому накопление эфиров ХС в мембранах эритроцитов у больных с наибольшими значениями концентрации сывороточного ХС свидетельствует о недостаточной функции ЛПВП.
Считается, что увеличение доли ХС неблагоприятно сказывается на динамических свойствах липидов мембраны, так как благодаря своему влиянию на подвижность жирнокислотных хвостов липидного окружения, он способствует повышению микровязкости мембраны. Накопление холестерина в бислое может приводить к ингибированию пероксидации структурированных липидов за счет ограничения молекулярной подвижности жирнокислотных остатков фосфолипидного бислоя. В этом случае ХС выступает в роли структурного антиоксиданта и делает фосфолипидные молекулы более устойчивыми к действию свободных радикалов, которые в свою очередь способны разрыхлять мембранные образования [Горбунов Н.В., 1993; Клебанов Г.И., 1998]. Поэтому поддержание необходимой жесткости мембраны холестерином подчас рассматривается как адаптивная реакция в условиях развития ряда патологических процессов.
Таблица 3
Результаты флуоресцентного зондирования флуорофором пирен и исследования активности Nа+,K+- ATФазы в мембранах эритроцитов
у больных сахарным диабетом типа 1 и 2, Me(Q1-Q3)
| Параметр | Здоровые доноры | Больные сахарным диабетом типа 1 | Больные сахарным диабетом типа 2 |
| J470/J370 (λв=285 нм), усл. ед. | 0,319 (0,275-0,408) | 0,257 (0,206-0,302) p1<0,01 | 0,334 (0,215-0,389) p1>0,05; p2<0,05 |
| J470/J370 (λв=340 нм), усл. ед. | 0,425 (0,340-0,449) | 0,340 (0,306-0,386) p1<0,01 | 0,412 (0,296-0,486) p1>0,05; p2>0,05 |
| J370/J390 (λв=340 нм), усл. ед. | 0,947 (0,932-0,953) | 0,962 (0,946-0,972) p1<0,01 | 0,944 (0,937-0,953) p1>0,05; p2<0,05 |
| Величина миграции энергии с триптофана на пирен, % | 55,77 (51,16-60,30) | 47,44 (43,02-61,76) p1>0,05 | 42,39 (34,14-50,81) p1<0,01; p2<0,05 |
| Активность Nа+,K+- ATФазы, мкмольPi/час·мг белка | 0,107 (0,039-0,183) | 0,032 (0,022-0,049) p1<0,05 | 0,061 (0,13-0,124) p1<0,05; p2>0,05 |
Наряду с этим динамические свойства липидов, входящих в состав бислоя, определяются большим количеством факторов, среди которых имеют значение степень насыщенности и длина ацильных цепей, состояние и количество белка в мембране, pH и ионная сила [Кагава Я., 1985; Болдырев А.А., 1990]. Для определения роли нарушения липопротеинового обмена в механизмах структурных изменений липидного бислоя мембран эритроцитов нами была проведена оценка их микровязкостных свойств у больных с ДЛП. Результаты наших исследований подтвердили наличие нарушения динамических свойств липидной фазы мембран эритроцитов у больных сахарным диабетом (табл. 3). Так, было установлено достоверное снижение соотношения интенсивностей флуоресценции эксимерных и мономерных молекул пирена (J470/J370), регистрируемых при длине волны возбуждающего света 340 нм (на 25% по сравнению с нормой), что является отражением возрастания микровязкости липидной фазы мембран. При исследовании структуры мембран эритроцитов у больных СД 1 в области белок-липидных контактов также было выявлено возрастание их микровязкости, на что указывало снижение (на 23% по сравнению с нормой) степени эксимеризации пирена при λВ=285 нм.
В ходе исследования было также показано, что выраженность выявленных изменений микровязкости мембраны зависела от длительности диабета. В частности, параметры флуоресценции зонда пирен у пациентов СД 1 со стажем диабета менее 5 лет значимо не отличались от аналогичных показателей у здоровых доноров (величина коэффициента J470/J370 при λВ=285 нм составила 0,323 (0,227-0,348) усл. ед., а при λВ=340 нм – 0,371 (0,342-0,417) усл. ед.). Повышение микровязкости мембран эритроцитов у пациентов с более длительным течением СД 1 по сравнению со здоровыми лицами было более выраженным, о чем свидетельствовали значимо более низкие показатели степени эксимеризации пирена как при λВ=285 нм (величина коэффициента J470/J370 составила 0,248 (0,203-0,293), p<0,01), так и при λВ=340 нм (величина коэффициента J470/J370 составила 0,321 (0,304-0,376) усл. ед., p<0,01). В группе пациентов с декомпенсацией углеводного обмена было отмечено достоверное увеличение коэффициента J370/J390 (0,962 (0,960-0,970) усл. ед., p<0,01) при λв=340 нм, отражающего степень полярности окружения зонда пирен. Изменение других параметров флуоресценции у больных СД 1 с различной компенсацией углеводного обмена имело одинаковую степень выраженности.
Липопротеины, участвуя в регуляции химического состава липидного компонента мембраны, влияют и на её структурные свойства, что подтверждается данными корреляционного анализа. Так, при обследовании больных СД 1 были выявлены корреляционные взаимосвязи между параметрами флуоресценции зонда пирен и показателями липидного спектра сыворотки крови. При этом у больных сахарным диабетом отмечалась положительная корреляция между коэффициентом полярности окружения зонда пирен и содержанием подфракции ЛПОНП1 (r=0,79, р<0,01). Было также обнаружено, что ЛПВП оказывают благоприятное влияние на величину микровязкости интегрального слоя липидной фазы мембран эритроцитов, на что указывало наличие прямой корреляции между содержанием ЛПВП и коэффициентом эксимеризации пирена при λв=340 нм (r=0,73, р<0,05).
Весьма интересные результаты были получены при оценке микровязкости липидной фазы мембран эритроцитов у пациентов с СД 2, отличавшихся существенными нарушениями липидного спектра сыворотки крови. Так, у обследованных больных СД 2 медиана значений параметров флуоресценции зонда пирен при взаимодействии с мембраной эритроцитов статистически не отличалась от соответствующего параметра в группе здоровых доноров (табл. 2). Однако анализ полученных данных методом К-средних позволил выявить среди пациентов СД 2 кластеры с разнонаправленными изменениями текучести мембран эритроцитов. Классификация данных проводилась с учетом коэффициента эксимеризации зонда пирен при λв=340 нм и λв=285 нм, а также коэффициента полярности J370/J390. В первый кластер были объединены 36% пациентов со сниженными величинами коэффициентов J470/J370 при λв=340 нм (средние значения составили 0,220±0,025 усл. ед.) и λв=285 нм (средние значения – 0,182±0,014 усл. ед.), а также уменьшенными значениями коэффициента полярности окружения зонда пирен (средние значения – 0,914±0,027 усл. ед.). У пациентов этого кластера было отмечено менее выраженное повышение индекса атерогенности по сравнению с больными других групп (значения составили 4,25 (3,83-5,76)). Особенностью данного кластера пациентов явилось высокое содержание в крови ТГ и ЛПОНП (содержание ТГ составило 248,57 (181,44-269,43) мг/дл, ЛПОНП – (285,68 (116,87-291,95) мг/дл)). ЛПОНП включают много насыщенных жирных кислот, что, вероятно, может обеспечивать дезорганизацию структуры мембран эритроцитов в связи с уплотнением липидных молекул относительно друг друга. При этом важно отметить снижение коэффициента полярности окружения зонда пирен в мембране эритроцитов J370/J390 (средние значения – 0,914±0,027 усл. ед.) у данной группы пациентов, свидетельствующее об уменьшении количества полярных групп интегральной зоны.
Наряду с этим была выделена группа больных СД 2, у которых мембрана эритроцитов характеризовалась повышенной текучестью и высокой полярностью гидрофобного слоя. У пациентов этой группы были зарегистрированы увеличенные значения J470/J370 при λв=340 нм (средние значения – 0,548±0,057 усл. ед.) и при λв=285 нм (средние значения – 0,574±0,040 усл.ед.). При этом средняя величина коэффициента полярности J370/J390 оказалась выше, чем у пациентов первого класса (0,955±0,007 усл. ед., p<0,05). В этот кластер были включены 25% больных СД 2 с наибольшими значениями индекса атерогенности (8,73 (5,37-12,30)) и наименьшим содержанием ЛПВП (243,03 (120,64-330,95) мг/дл). Вероятно, гидрофильные продукты ПОЛ в больших количествах, накапливаясь в мембранах при недостаточном содержании в плазме ЛПВП, обладают детергентным действием и способствуют повышению разупорядоченности липидного бислоя [Каган В.Е. и соавт., 1975; Новиков К.Н., 2004]. Это в свою очередь вызывает увеличение подвижности полипептидных цепей интегральных белков в мембране. Высокая полярность группировок (гидроперекисных, гидроксильных, карбонильных), появляющихся в результате индукции ПОЛ в полиеновых “хвостах” фосфолипидов, нарушает гидрофобные белок-липидные взаимодействия в мембране. Таким образом, из-за увеличения степени ненасыщенности, избыточной пероксидации изменяется направленность ацильных остатков.
Показатели микровязкостных свойств мембраны эритроцитов у пациентов третьей группы, в состав которой были включены 39% обследованных пациентов СД 2, соответствовали следующим значениям: средние величины коэффициента J470/J370 составили 0,434±0,018 усл. ед. при λв=340 нм и 0,348±0,016 усл. ед. при λв=285 нм. Выраженность нарушений липидного обмена у этих больных также носила практически промежуточный характер. Таким образом, отмеченное нами разнообразие структурных нарушений мембран эритроцитов, видимо, определяется различными проявлениями воздействия ряда факторов, имеющих влияние на биофизические свойства мембраны. К их числу, на наш взгляд, правомерно отнести нарушения липидного обмена.
При рассмотрении изменений структурных свойств мембран эритроцитов у пациентов с различными типами ДЛП отмечено увеличение микровязкости мембран эритроцитов у больных с повышенным содержанием в сыворотке крови ЛПОНП. Структурные свойства мембран красных кровяных клеток у больных с другими типами ДЛП не различались ни между собой, ни с соответствующими параметрами у здоровых доноров.
Как показало проведенное нами исследование, в мембранах эритроцитов у больных СД угнетена активность Na+,K+-ATФазы (табл. 3). Не исключено, что ингибирование активности Na+,K+-ATФазы является результатом выявленной структурной модификации липидного матрикса мембран эритроцитов. Общепринятой является точка зрения о том, что регулирующая роль липид-белковых взаимодействий для Na+,K+-ATФазы сводится к обеспечению ее каталитических функций и переходов между различными конформационными состояниями посредством поддержания определенной микровязкости мембраны, а также за счет взаимодействия заряженных боковых цепей этих ферментов с полярными головками мембранных липидов [Грибанов Г.А., 1991; Зелинский Б.А., 1994; Успенская Ю.А., 2002].
Рассматривая механизмы структурной модификации мембраны эритроцитов, важно отметить множественность внутри- и внеклеточных эффекторов, которые могут индуцировать друг друга или оказывать свое влияние изолированно. При СД имеют значение гипергликемия, стадия ее компенсации, активация свободно-радикального окисления и недостаточность антиоксидантных систем, а также нарушения липидного обмена. В частности известно, что липопротеины как одна из регуляторных систем организма имеет значение для поддержания структурного гомеостаза клеточных мембран. Нарушение транспорта липидов может привести к срыву реакций адаптации и формированию необратимых нарушений в мембранах. Именно поэтому изучение изолированного действия липопротеинов на мембрану эритроцитов в опытах in vitro представилось нам весьма интересным.
Выше было указано на важную роль липопротеинов в обновлении мембранных компонентов, причем ЛПВП, ЛПНП и ЛПОНП могут оказывать разнонаправленные эффекты, меняя тем самым структурно-функциональные свойства мембраны. Некоторые авторы предполагают факт возникновения комплекса эритроцит-липопротеин, в котором и происходит свободная диффузия ХС [Болдырев А.А., 1990]. Изучая спектральные характеристики взаимодействия липотропного зонда пирен с мембраной эритроцитов крыс до и после инкубации мембранной взвеси с липопротеинами основных классов (ЛПВП, ЛПНП и ЛПОНП), мы обнаружили модифицирующее влияние, проявляющееся в изменении текучести липидной фазы и характера белок-липидных взаимодействий (табл. 4). Очевидно, что образование комплекса липопротеин-мембрана сопровождается кратковременными локальными изменениями физико-химических свойств бислойной структуры, которые способствуют молекулярному обмену липидов. Сформировавшийся комплекс, вероятно, представляет собой особую структуру с существованием раздела фаз, преодолевать который помогают силы гидрофобного взаимодействия.
Таблица 4
Результаты флуоресцентного зондирования флуорофором пирен мембраны
эритроцитов у крыс при инкубации с липопротеинами высокой плотности (ЛПВП),
липопротеинами низкой плотности (ЛПНП) и липопротеинами очень низкой
плотности (ЛПОНП), Me(Q1-Q3)
| Этапы исследования | Параметры | |||
| J470/J370 (λв=285 нм), усл. ед. | J470/J370 (λв=340 нм), усл. ед. | J370/J390 (λв=340 нм), усл. ед. | Величина миграции энергии с триптофана на пирен, % | |
| Взаимодействие с ЛПВП: | ||||
| до инкубации | 0,162 (0,134-0,195) | 0,242 (0,226-0,257) | 0,780 (0,773-0,793) | 43,75 (40,00-44,74) |
| после инкубации | 0,229 (0,159-0,275) p>0,05 | 0,347 (0,280-0,380) p<0,05 | 0,775 (0,767-0,844) p>0,05 | 47,00 (43,48-47,59) p>0,05 |
| Взаимодействие с ЛПНП: | ||||
| до инкубации | 0,203 (0,192-0,234) | 0,332 (0,253-0,438) | 0,829 (0,778-0,880) | 46,50 (43,67-50,00) |
| после инкубации | 0,235 (0,222-0,286) p>0,05 | 0,408 (0,373-0,428) p<0,05 | 0,820 (0,761-0,833) p>0,05 | 53,12 (46,15-54,84) p<0,05 |
| Взаимодействие с ЛПОНП: | ||||
| до инкубации | 0,230 (0,183-0,294) | 0,332 (0,245-0,439) | 0,829 (0,776-0,844) | 46,50 (42,63-50,00) |
| после инкубации | 0,253 (0,200-0,296) p<0,05 | 0,357 (0,312-0,446) p<0,05 | 0,810 (0,771-0,869) p>0,05 | 50,00 (47,00-64,29) p<0,05 |
Примечание: р – уровень значимости различий по сравнению со значениями параметров, характеризующих состояние мембран эритроцитов у крыс до инкубации с липопротеинами
Ранее отмечалось, что мембрану ХС покидает в этерифицированном виде. Будучи менее полярными соединениями, эфиры ХС легче проходят через границу раздела фаз, где связываются с ЛПВП и подвергаются дальнейшему метаболизму. Возможно, приток ХС от ЛПНП и ЛПОНП в мембрану проходит похожим образом, только в обратном направлении. Важное значение в этих перемещениях имеют входящие в состав липопротеинов апобелки. Известно, что белки снижают энергию активации, необходимую для десорбции липидов в жидкую фазу, тем самым облегчая процесс переноса амфипатной молекулы [Sprong H., 2001]. Кроме того, особенности строения аполипопротеинов, высокая степень спирализации структуры и наличие амфипатных областей, ответственных за связывание с липидами, обусловливает их детергентные свойства [Поляков Л.М., Панин Л.Е., 2000]. На наш взгляд, изучение влияния очищенных аполипопротеинов (А-I, С и Е) при ДЛП является закономерным продолжением исследования механизмов модификации мембраны эритроцитов. Нами были обнаружены принципиальные различия воздействия этих апобелков. В частности, аполипопротеин А-I, наибольшее количество которого присутствует в ЛПВП, оказывал «разжижающее» действие на мембрану эритроцитов. Об этом свидетельствовали достоверное повышение отношения интенсивностей флуоресценции димерной и мономерной форм флуорофора пирен (J470/J370) после инкубации с раствором апобелка как при длине волны возбуждающего света 340 нм (значения коэффициента до инкубации составили 0,240 (0,237-0,253) усл. ед., после – 0,353 (0,318-0,403) усл.ед. при, p<0,05), так и при 285 нм (значения коэффициента до – 0,167 (0,159-0,190) усл. ед., после инкубации составили 0,283 (0,214-0,322) усл.ед., p<0,05). При этом в мембране изменялся характер белок-липидных взаимодействий. Медиана величины миграции энергии с триптофана на пирен увеличилась с 42,63 (40,00-43,75) % до и до 46,06 (44,64-47,62) % – после инкубации (p<0,05).
Известно, что апо A-I способен связывать свободный ХС, являясь очень сильным его акцептором, и опосредует его обратный транспорт в печень. По данным А.Н. Климова и соавт. [1984, 1992], стерин способен образовывать комплексы с апопротеином A-I без вовлечения ФЛ. При этом образование комплекса происходит путем взаимодействия холестеринового гидроксила с гуанидиновыми и ε-аминогруппами аминокислотных остатков белка. При этом происходит уменьшение содержания ХС в мембране, что сопровождается увеличением количества связанной воды и снижением жесткости липидного бислоя.
Иное влияние оказывает на мембрану аполипопротеин С. Исследование интенсивности флуоресценции димерной и мономерной форм флуорофора пирен (J470/J370) при длине волны возбуждающего света 285 нм выявило снижение значения коэффициента эксимеризации, свидетельствующее об увеличении жесткости мембраны в зоне белок-липидных взаимодействий. Значения изучаемого параметра до инкубации с апо С составили 0,200 (0,166-0,250) усл. ед., после инкубации – 0,174 (0,118-0,214) усл. ед. Величина соотношения интенсивностей флуоресценции мономеров пирена (J370/J390) при λв=340 нм после взаимодействия с апо С оказалась достоверно ниже и составила 0,763 (0,703-0,784) усл. ед. (при 0,831 (0,772-0,950) усл.ед. до инкубации p<0,05). Функциональные особенности апо С позволяют нам предположить возможный механизм подобного взаимодействия: скорее всего, выявленный эффект обусловлен связыванием аполипопротеином жирнокислотных хвостов фосфолипидов [Панин Л.Е. и соавт., 2001].
Наряду с этим оказалось, что аполипопротеин Е не изменяет спектральные характеристики взаимодействия мембраны эритроцита с зондом пирен, обнаруживая свою химическую инертность по отношению к липидному бислою в условиях нашего эксперимента. Известно, что апо Е также, как и апо A-I, является сильным акцептором ХС. Однако отсутстсвие изменений биофизических свойств мембраны после инкубации с этим аполипопротеином позволяет думать, что для влияния на структуру мембран необходимы какие-то другие дополнительные факторы.
Таким образом, состояние липидной фазы мембран эритроцитов обеспечивается внутриклеточными и внеклеточными механизмами регуляции (рис. 4). Для эритроцита – безъядерной клетки – среди внутриклеточных механизмов имеют значение прежде всего процессы свободно-радикального окисления ненасыщенных жирных кислот, фосфолиполиз, транслокация липидных молекул в мембранах и др. Среди плазменных факторов, влияющих на метаболизм липидов мембран эритроцитов, рассматривается значение липопротеинов как структур с аналогичной молекулярной организацией.
По результатам нашего исследования, у пациентов с сахарным диабетом параметры микровязкости мембран эритроцитов были взаимосвязаны с показателями липидного обмена, что подтверждалось данными корреляционного анализа. Кроме того, выделенные клинические группы больных СД 1, различавшиеся по степени выраженности нарушений липидного обмена, отличались и по степени выраженности структурной дезорганизации мембран эритроцитов. У пациентов с СД 2 закономерности изменения структурных свойств мембраны красных клеток крови оказались сложнее и зависели от характера нарушения липидного обмена. Факт взаимодействия мембран эритроцитов с липопротеинами был продемонстрирован экспериментально, при этом мембранотропным эффектом обладали и аполипопротеины.
ВЫВОДЫ
- Нарушения структуры мембран эритроцитов у больных сахарным диабетом типа 1 и 2 с дислипопротеинемиями характеризуются изменениями липидного состава (увеличение содержания холестерина и его эфиров, лизофосфолипидов, фосфатидилсерина при одновременном уменьшении доли фосфатидилхолина) и микровязкости липидного бислоя, нарушением межмолекулярных липид-липидных и белок-липидных взаимодействий, а также угнетением активности Na+,K+-АТФазы.
- Изменения микровязкости мембран эритроцитов, нарушение липид-липидных и белок-липидных взаимодействий у больных сахарным диабетом типа 1 и 2 коррелируют с увеличением содержания липопротеинов низкой и очень низкой плотности в сыворотке крови. Низкое содержание липопротеинов высокой плотности является дополнительным фактором, усугубляющим нарушения структуры мембран эритроцитов.
- Нарушения липопротеинового состава крови и степень дезорганизации мембран эритроцитов при сахарном диабете типа 2 более выражены, чем при сахарном диабете типа 1. Выраженность указанных изменений при сахарном диабете типа 1 зависит от длительности заболевания, наличия декомпенсации углеводного обмена и сосудистых осложнений.
- Инкубация мембран эритроцитов с липопротеинами низкой и очень низкой плотности в опытах in vitro приводит к выраженным наноструктурным изменениям и сопровождается изменениями липид-липидных и белок-липидных взаимодействий, снижением микровязкости эритроцитарных мембран.
- Изменения структурных свойств теней эритроцитов крыс под влиянием аполипопротеина A-I и липопротеинов высокой плотности, обладающих свойством акцептировать мембранный холестерин, в опытах in vitro имеют одинаковую направленность и проявляется уменьшением микровязкости липидной фазы мембран эритроцитов.
- Аполипопротеины С вызывают увеличение жесткости анулярной зоны мембран эритроцитов и уменьшение их полярности . Аполипопротеин Е в опытах in vitro не оказывает мембранотропного эффекта на тени эритроцитов.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
- Бутусова В.Н., Ананина Е.А. Феноменология модификации плазматической мембраны клеток при дислипидемии // VI международный конгресс молодых ученых и специалистов «Науки о человеке», г. Томск, 20-21 мая 2005 г. - Томск, 2005. - С.2-3.
- Бутусова В.Н., Рязанцева Н.В., Кравец Е.Б, Яковлева Н.М., Кощевец Т.Ю., Ананина Е.А., Жаворонок Т.В., Биктасова А.К. Особенности структуры и функции мембраны эритроцитов у больных сахарным диабетом с сопутствующей гиперхолестеринемией // Материалы научно-практической межрегиональной конференции, посвященной 70-летию ГОУ ВПО Новосибирской государственной медицинской академии «Актуальные проблемы современной эндокринологии». - Новосибирск: Изд-во НГТУ. - 2005. - С. 88-89.
- Кравец Е.Б., Рязанцева Н.В., Яковлева Н.М., Бутусова В.Н., Тухватулин Р.Т., Новикова Л.К. Молекулярные нарушения мембраны эритроцитов при сосудистых осложнениях сахарного диабета типа 1 // Сахарный диабет. - 2006. - № 1. - С. 10-17.
- Яковлева Н.М., Рязанцева Н.В., Бутусова В.Н. Изменения структурно-функционального статуса мембраны эритроцитов при сосудистых осложнениях сахарного диабета типа 1 // XI Всероссийская научно-практическая конференция «Молодые ученые в медицине», г. Казань, 26-27 апреля 2006 г. - Казань: ЗАО « Новое знание», 2006. - С.189.
- Новицкий В.В., Рязанцева Н.В., Степовая Е.А., Федорова Т.С., Кравец Е.Б., Иванов В.В., Жаворонок Т.В., Часовских Н.Ю., Чудакова О.М., Бутусова В.Н., Яковлева Н.М. Молекулярные нарушения мембраны эритроцитов при патологии разного генеза являются типовой реакцией организма: контуры проблемы // Бюллетень сибирской медицины. - 2006. - № 2. - С. 62-67.
- Кравец Е.Б., Рязанцева Н.В., Яковлева Н.М., Бутусова В.Н., Чудакова О.М., Сапрыкина Э.В. Молекулярные нарушения мембран эритроцитов и тромбоцитов при сосудистых осложнениях сахарного диабета типа 1 // Бюллетень сибирской медицины. - 2006. - № 4. - С. - 33-41.
СПИСОК СОКРАЩЕНИЙ
Апо – аполипопротеин
ДЛП – дислипопротеинемия
ЛПВП – липопротеины высокой плотности
ЛПНП – липопротеины низкой плотности
ЛПОНП – липопротеины очень низкой плотности
ПОЛ – перекисное окисление липидов
СД 1 – сахарный диабет типа 1
СД 2 – сахарный диабет типа 2
ТГ – триацилглицериды
ХМ – хиломикроны
ХС – холестерин
Автор выражает благодарность заведующей кафедрой эндокринологии и диабетологии СибГМУ проф. Е.Б. Кравец за предоставление клинического материала, д.б.н. Ф.В. Тузикову (Институт молекулярной биологии ГНЦ вирусологии и биотехнологии "Вектор") за проведение исследования липопротеинового спектра крови у больных и здоровых доноров, а также благодарит проф. Л.М. Полякова и весь коллектив лаборатории молекулярной биотехнологии НИИ биохимии СО РАМН за предоставленные липопротеины и аполипопротеины для проведения экспериментального исследования.